
Methylene Blue Is a Nonspecific Protein–Protein Interaction Inhibitor with Potential for Repurposing as an Antiviral for COVID-19


 , ,
, , 

Abstract
:1. Introduction
2. Results
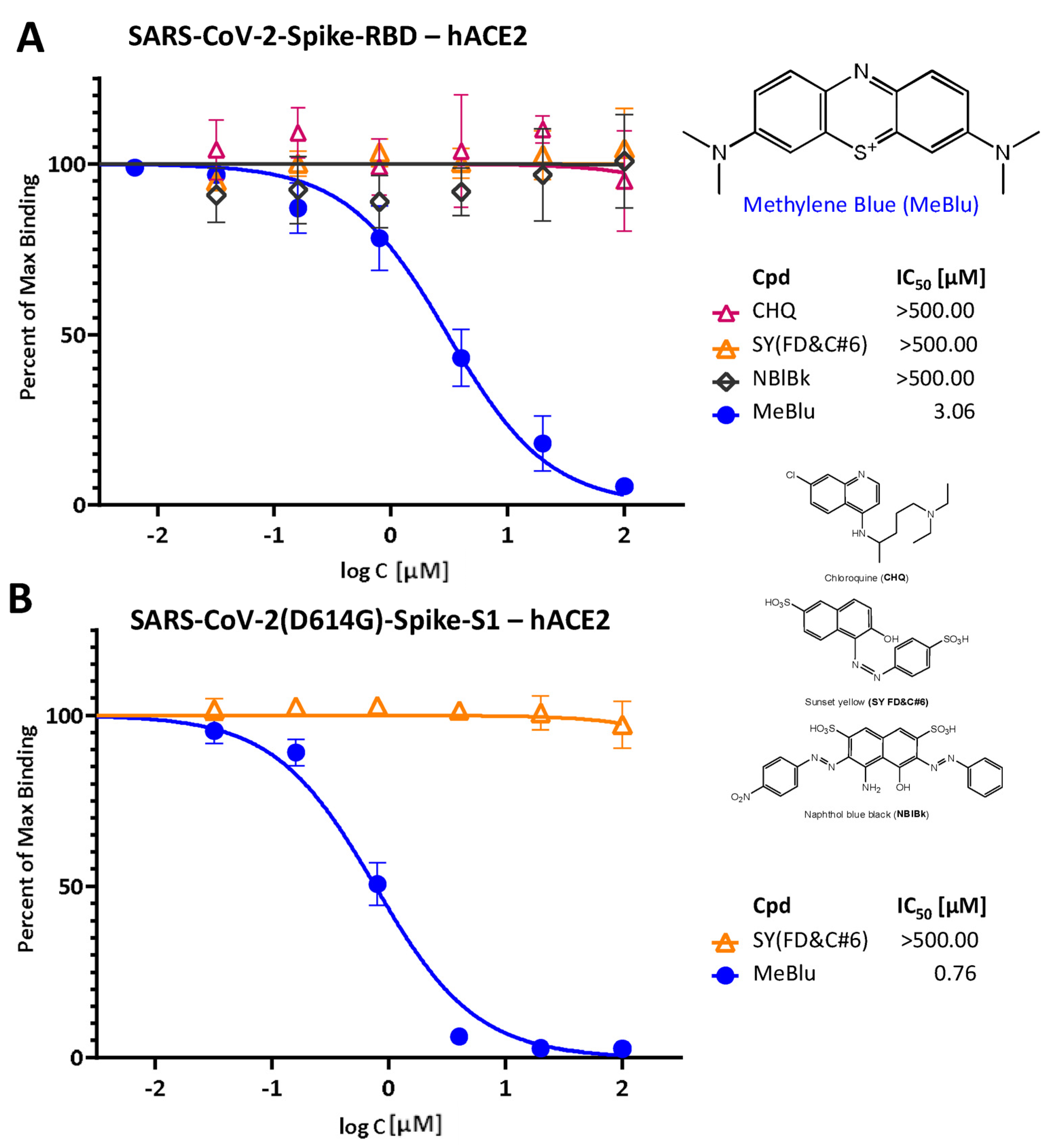
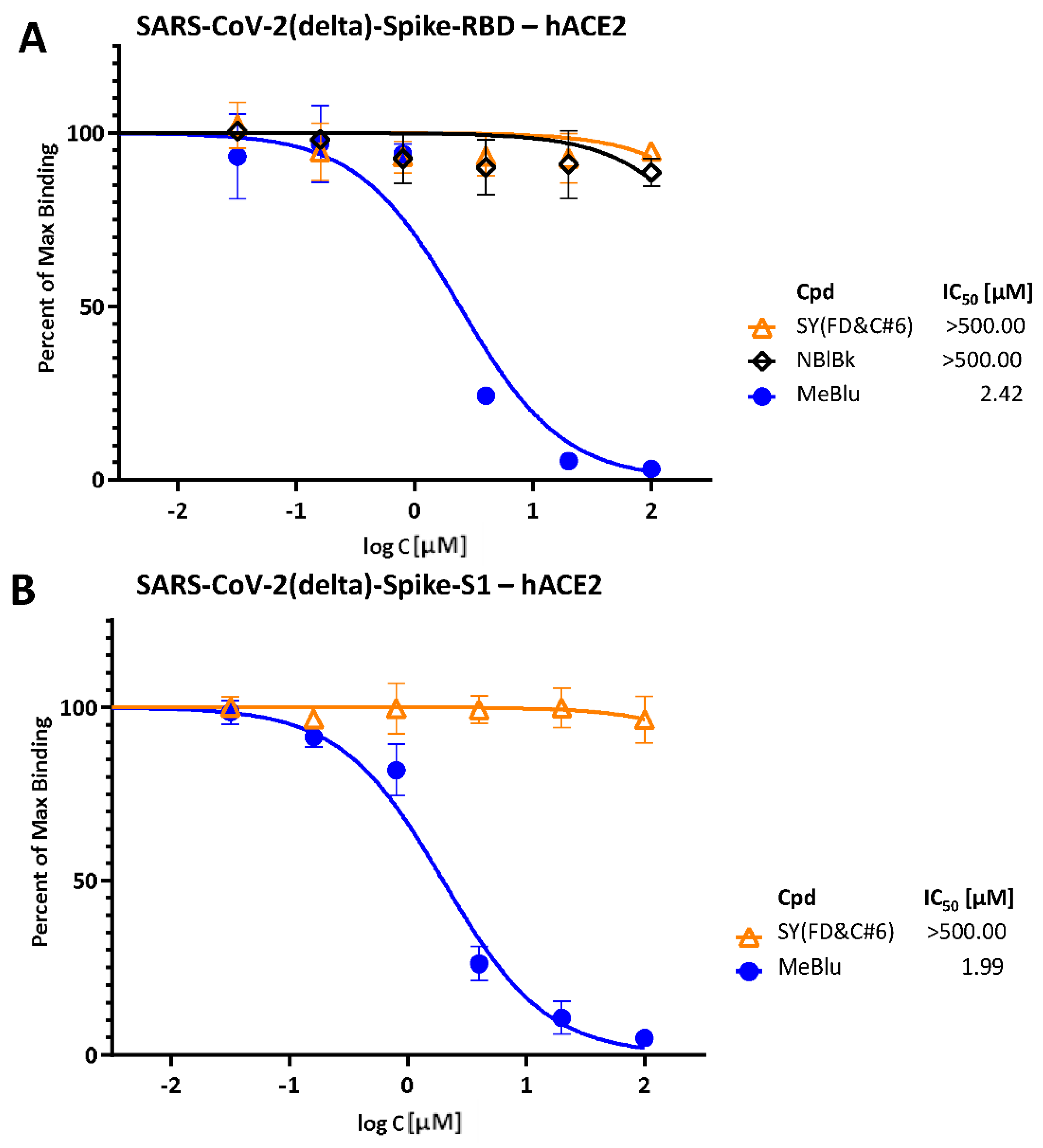
2.1. Inhibition of SARS-CoV-2 Spike–hACE2 PPIs
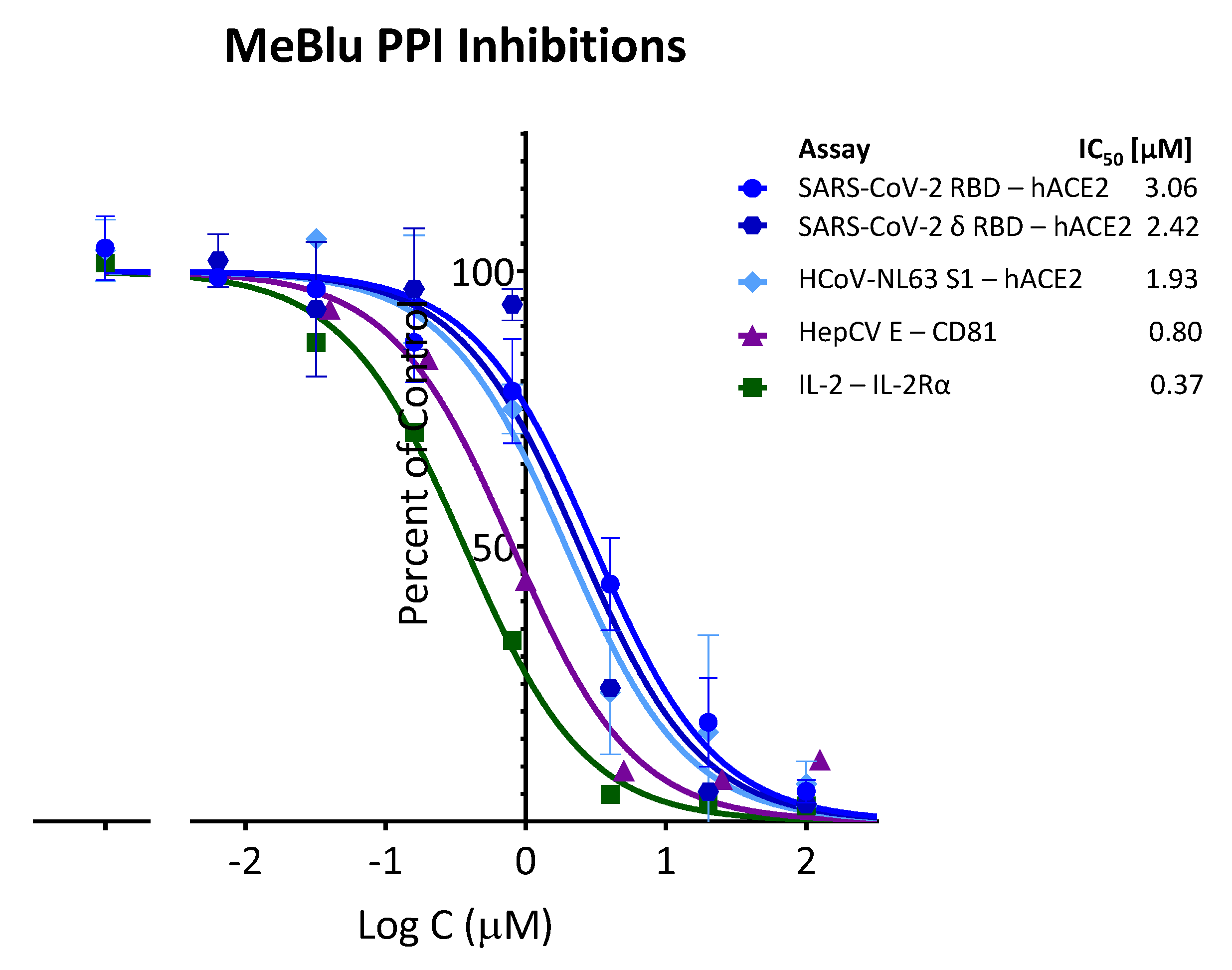
2.2. Inhibition of Other PPIs
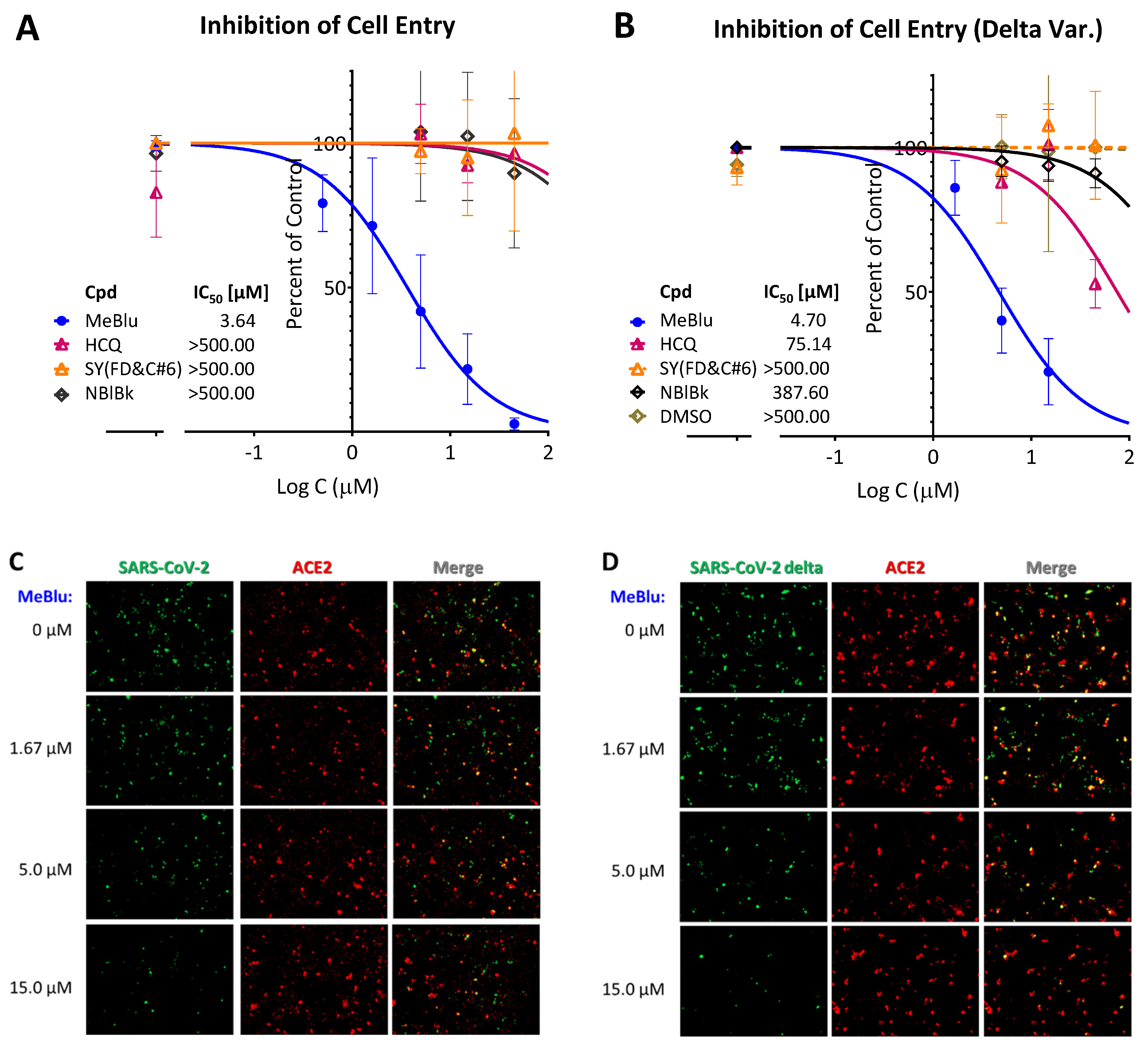
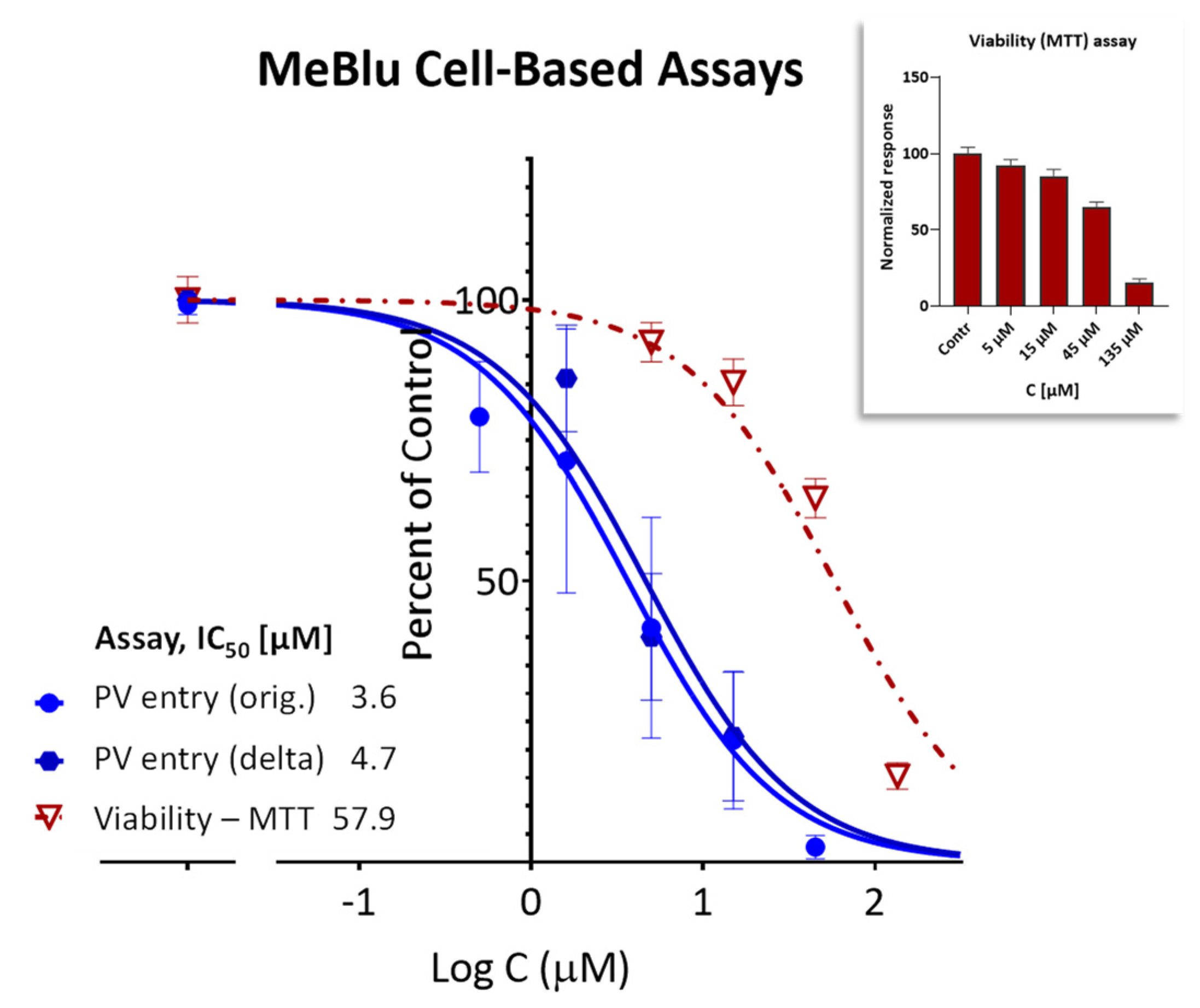
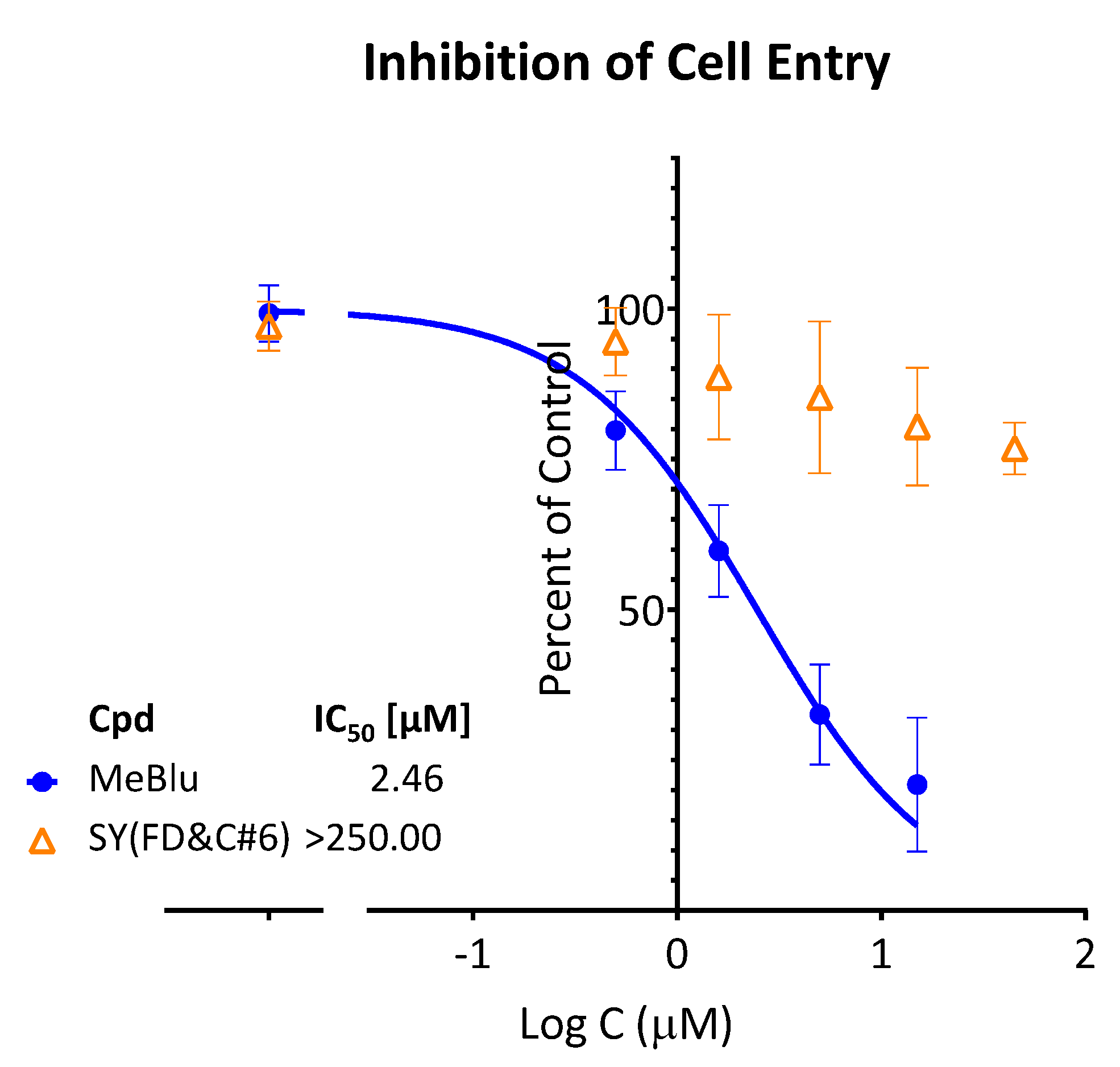
2.3. SARS-CoV-2 Pseudovirus Entry Inhibition
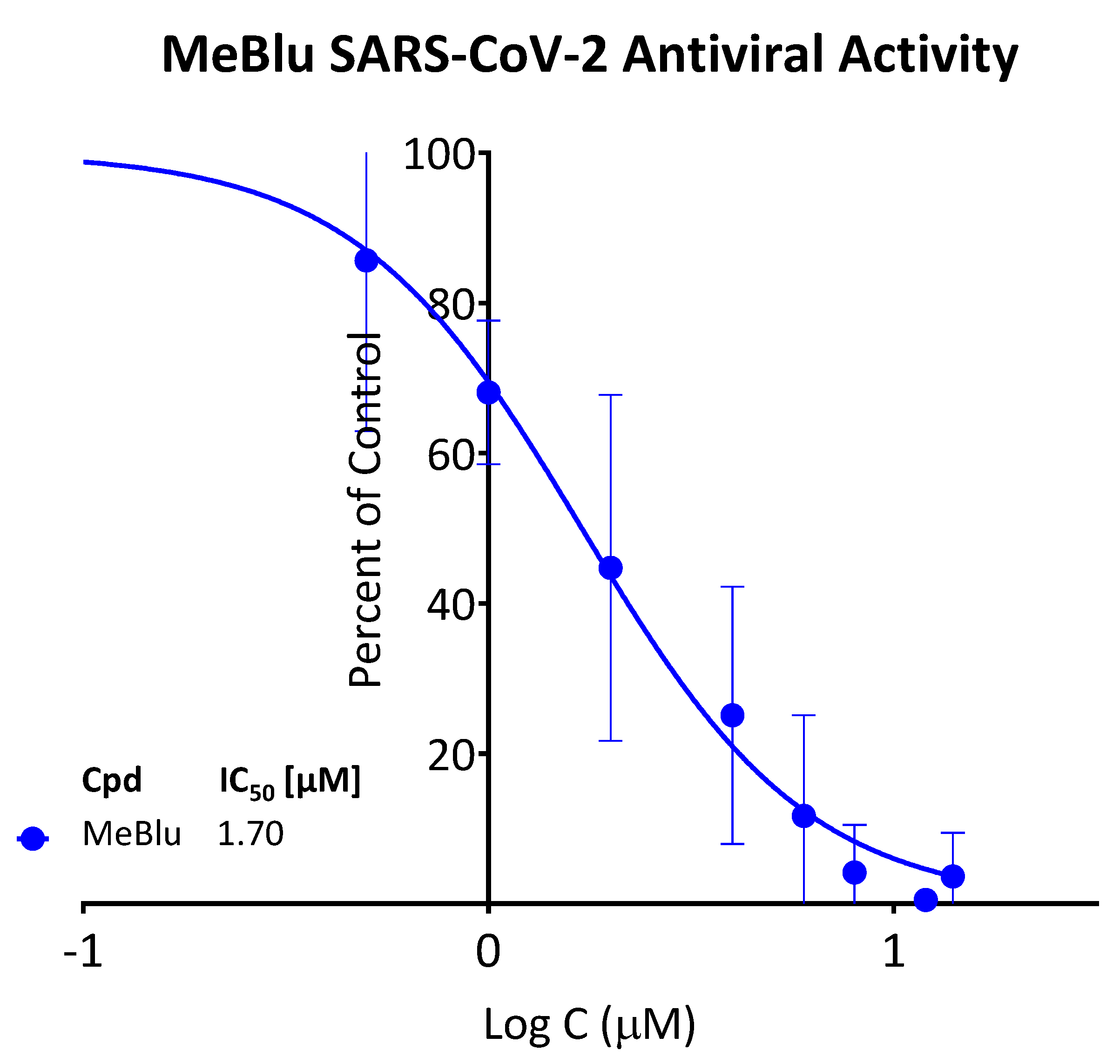
2.4. SARS-CoV-2 Antiviral Activity
3. Discussion
4. Materials and Methods
4.1. Binding Assays
4.2. SARS-CoV-2 Pseudovirus Assays
4.3. Cytotoxicity Assay
4.4. Surface Plasmon Resonance
4.5. In Vitro SARS-CoV-2 Antiviral Assay
4.6. Statistics and Data Fitting
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACE2 | angiotensin converting enzyme 2 |
| CoV | coronavirus |
| MeBlu | methylene blue |
| NBlBk | naphthol blue black |
| PPI | protein–protein interaction |
| SARS | severe acute respiratory syndrome |
| SMI | small-molecule inhibitor |
| SPR | surface plasmon resonance |
| SY | sunset yellow FCF |
| TNF | tumor necrosis factor |
| VoC | variant of concern |
References
- Matheson, N.J.; Lehner, P.J. How does SARS-CoV-2 cause COVID-19? Science 2020, 369, 510–511. [Google Scholar] [CrossRef] [PubMed]
- Santacroce, L.; Charitos, I.A.; Carretta, D.M.; De Nitto, E.; Lovero, R. The human coronaviruses (HCoVs) and the molecular mechanisms of SARS-CoV-2 infection. J. Mol. Med. 2021, 99, 93–106. [Google Scholar] [CrossRef] [PubMed]
- V’Kovski, P.; Kratzel, A.; Steiner, S.; Stalder, H.; Thiel, V. Coronavirus biology and replication: Implications for SARS-CoV-2. Nat. Rev. Microbiol. 2021, 19, 155–170. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, V.; Beer, J.C.; Sankaranarayanan, N.V.; Swanson-Mungerson, M.; Desai, U.R. Discovering small-molecule therapeutics against SARS-CoV-2. Drug Discov. Today 2020, 25, 1535–1544. [Google Scholar] [CrossRef] [PubMed]
- Nabel, K.G.; Clark, S.A.; Shankar, S.; Pan, J.; Clark, L.E.; Yang, P.; Coscia, A.; McKay, L.G.A.; Varnum, H.H.; Brusic, V.; et al. Structural basis for continued antibody evasion by the SARS-CoV-2 receptor binding domain. Science 2022, 375, eabl6251. [Google Scholar] [CrossRef] [PubMed]
- WHO Solidarity Trial Consortium. Repurposed antiviral drugs for COVID-19—Interim WHO Solidarity trial results. N. Engl. J. Med. 2020, 384, 497–511. [Google Scholar] [CrossRef]
- Tummino, T.A.; Rezelj, V.V.; Fischer, B.; Fischer, A.; O’Meara, M.J.; Monel, B.; Vallet, T.; White, K.M.; Zhang, Z.; Alon, A.; et al. Drug-induced phospholipidosis confounds drug repurposing for SARS-CoV-2. Science 2021, 373, 541–547. [Google Scholar] [CrossRef]
- Jayk Bernal, A.; Gomes da Silva, M.M.; Musungaie, D.B.; Kovalchuk, E.; Gonzalez, A.; Delos Reyes, V.; Martin-Quiros, A.; Caraco, Y.; Williams-Diaz, A.; Brown, M.L.; et al. Molnupiravir for oral treatment of COVID-19 in nonhospitalized patients. N. Engl. J. Med. 2021, 386, 509–520. [Google Scholar] [CrossRef]
- Owen, D.R.; Allerton, C.M.N.; Anderson, A.S.; Aschenbrenner, L.; Avery, M.; Berritt, S.; Boras, B.; Cardin, R.D.; Carlo, A.; Coffman, K.J.; et al. An oral SARS-CoV-2 M(pro) inhibitor clinical candidate for the treatment of COVID-19. Science 2021, 374, 1586–1593. [Google Scholar] [CrossRef]
- Chen, J.; Song, Y.; Bojadzic, D.; Tamayo-Garcia, A.; Landin, A.M.; Blomberg, B.B.; Buchwald, P. Small-molecule inhibitors of the CD40-CD40L costimulatory protein-protein interaction. J. Med. Chem. 2017, 60, 8906–8922. [Google Scholar] [CrossRef]
- Bojadzic, D.; Buchwald, P. Toward small-molecule inhibition of protein-protein interactions: General aspects and recent progress in targeting costimulatory and coinhibitory (immune checkpoint) interactions. Curr. Top. Med. Chem. 2018, 18, 674–699. [Google Scholar] [CrossRef] [PubMed]
- Bojadzic, D.; Alcazar, O.; Chen, J.; Chuang, S.T.; Condor Capcha, J.M.; Shehadeh, L.A.; Buchwald, P. Small-molecule inhibitors of the coronavirus spike—ACE2 protein-protein interaction as blockers of viral attachment and entry for SARS-CoV-2. ACS Infect. Dis. 2021, 7, 1519–1534. [Google Scholar] [CrossRef] [PubMed]
- Souers, A.J.; Leverson, J.D.; Boghaert, E.R.; Ackler, S.L.; Catron, N.D.; Chen, J.; Dayton, B.D.; Ding, H.; Enschede, S.H.; Fairbrother, W.J.; et al. ABT-199, a potent and selective BCL-2 inhibitor, achieves antitumor activity while sparing platelets. Nat. Med. 2013, 19, 202–208. [Google Scholar] [CrossRef] [PubMed]
- Gadek, T.R.; Burdick, D.J.; McDowell, R.S.; Stanley, M.S.; Marsters, J.C., Jr.; Paris, K.J.; Oare, D.A.; Reynolds, M.E.; Ladner, C.; Zioncheck, K.A.; et al. Generation of an LFA-1 antagonist by the transfer of the ICAM-1 immunoregulatory epitope to a small molecule. Science 2002, 295, 1086–1089. [Google Scholar] [CrossRef] [PubMed]
- Meanwell, N.A.; Krystal, M.R.; Nowicka-Sans, B.; Langley, D.R.; Conlon, D.A.; Eastgate, M.D.; Grasela, D.M.; Timmins, P.; Wang, T.; Kadow, J.F. Inhibitors of HIV-1 attachment: The discovery and development of temsavir and its prodrug fostemsavir. J. Med. Chem. 2018, 61, 62–80. [Google Scholar] [CrossRef] [PubMed]
- Arkin, M.R.; Wells, J.A. Small-molecule inhibitors of protein-protein interactions: Progressing towards the dream. Nat. Rev. Drug Discov. 2004, 3, 301–317. [Google Scholar] [CrossRef]
- Milroy, L.G.; Grossmann, T.N.; Hennig, S.; Brunsveld, L.; Ottmann, C. Modulators of protein-protein interactions. Chem. Rev. 2014, 114, 4695–4748. [Google Scholar] [CrossRef] [Green Version]
- Scott, D.E.; Bayly, A.R.; Abell, C.; Skidmore, J. Small molecules, big targets: Drug discovery faces the protein-protein interaction challenge. Nat. Rev. Drug Discov. 2016, 15, 533–550. [Google Scholar] [CrossRef] [Green Version]
- Clifton, J., 2nd; Leikin, J.B. Methylene blue. Am. J. Ther. 2003, 10, 289–291. [Google Scholar] [CrossRef]
- Schirmer, R.H.; Adler, H.; Pickhardt, M.; Mandelkow, E. “Lest we forget you—methylene blue...”. Neurobiol. Aging 2011, 32, 2325.e7–2325.e16. [Google Scholar] [CrossRef]
- Oz, M.; Lorke, D.E.; Hasan, M.; Petroianu, G.A. Cellular and molecular actions of Methylene Blue in the nervous system. Med. Res. Rev. 2011, 31, 93–117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bistas, E.; Sanghavi, D. Methylene blue. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Bojadzic, D.; Alcazar, O.; Buchwald, P. Methylene blue inhibits the SARS-CoV-2 spike—ACE2 protein-protein interaction—a mechanism that can contribute to its antiviral activity against COVID-19. Front. Pharmacol. 2021, 11, 600372. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Jackson, C.B.; Mou, H.; Ojha, A.; Peng, H.; Quinlan, B.D.; Rangarajan, E.S.; Pan, A.; Vanderheiden, A.; Suthar, M.S.; et al. SARS-CoV-2 spike-protein D614G mutation increases virion spike density and infectivity. Nat. Commun. 2020, 11, 6013. [Google Scholar] [CrossRef] [PubMed]
- CDC, Center for Disease Control and Prevention. SARS-CoV-2 Variant Classifications and Definitions. 2021. Available online: https://www.cdc.gov/coronavirus/2019-ncov/variants/variant-classifications.html (accessed on 14 February 2022).
- Kupferschmidt, K. Evolving threat. Science 2021, 373, 844–849. [Google Scholar] [CrossRef] [PubMed]
- McGovern, S.L.; Caselli, E.; Grigorieff, N.; Shoichet, B.K. A common mechanism underlying promiscuous inhibitors from virtual and high-throughput screening. J. Med. Chem. 2002, 45, 1712–1722. [Google Scholar] [CrossRef] [PubMed]
- Aldrich, C.; Bertozzi, C.; Georg, G.I.; Kiessling, L.; Lindsley, C.; Liotta, D.; Merz, K.M., Jr.; Schepartz, A.; Wang, S. The ecstasy and agony of assay interference compounds. J. Med. Chem. 2017, 60, 2165–2168. [Google Scholar] [CrossRef] [Green Version]
- Arkin, M.R.; Tang, Y.; Wells, J.A. Small-molecule inhibitors of protein-protein interactions: Progressing toward the reality. Chem. Biol. 2014, 21, 1102–1114. [Google Scholar] [CrossRef] [Green Version]
- Condor Capcha, J.M.; Lambert, G.; Dykxhoorn, D.M.; Salerno, A.G.; Hare, J.M.; Whitt, M.A.; Pahwa, S.; Jayaweera, D.T.; Shehadeh, L.A. Generation of SARS-CoV-2 spike pseudotyped virus for viral entry and neutralization assays: A 1-week protocol. Front. Cardiovasc. Med. 2020, 7, 618651. [Google Scholar] [CrossRef]
- Bajusz, D.; Wade, W.S.; Satala, G.; Bojarski, A.J.; Ilas, J.; Ebner, J.; Grebien, F.; Papp, H.; Jakab, F.; Douangamath, A.; et al. Exploring protein hotspots by optimized fragment pharmacophores. Nat. Commun. 2021, 12, 3201. [Google Scholar] [CrossRef]
- Bereczki, I.; Papp, H.; Kuczmog, A.; Madai, M.; Nagy, V.; Agócs, A.; Batta, G.; Milánkovits, M.; Ostorházi, E.; Mitrović, A.; et al. Natural apocarotenoids and their synthetic glycopeptide conjugates inhibit SARS-CoV-2 replication. Pharmaceuticals 2021, 14, 1111. [Google Scholar] [CrossRef]
- Lozano, M.; Cid, J.; Muller, T.H. Plasma treated with methylene blue and light: Clinical efficacy and safety profile. Transfus. Med. Rev. 2013, 27, 235–240. [Google Scholar] [CrossRef] [PubMed]
- Alemany, A.; Millat-Martinez, P.; Corbacho-Monne, M.; Malchair, P.; Ouchi, D.; Ruiz-Comellas, A.; Ramirez-Morros, A.; Rodriguez Codina, J.; Amado Simon, R.; Videla, S.; et al. High-titre methylene blue-treated convalescent plasma as an early treatment for outpatients with COVID-19: A randomised, placebo-controlled trial. Lancet Respir. Med. 2022, 10, 278–288. [Google Scholar] [CrossRef]
- Coghi, P.; Yang, L.J.; Ng, J.P.L.; Haynes, R.K.; Memo, M.; Gianoncelli, A.; Wong, V.K.W.; Ribaudo, G. A drug repurposing approach for antimalarials interfering with SARS-CoV-2 spike protein receptor binding domain (RBD) and human angiotensin-converting enzyme 2 (ACE2). Pharmaceuticals 2021, 14, 954. [Google Scholar] [CrossRef] [PubMed]
- Cagno, V.; Medaglia, C.; Cerny, A.; Cerny, T.; Zwygart, A.C.; Cerny, E.; Tapparel, C. Methylene blue has a potent antiviral activity against SARS-CoV-2 and H1N1 influenza virus in the absence of UV-activation in vitro. Sci. Rep. 2021, 11, 14295. [Google Scholar] [CrossRef] [PubMed]
- Gendrot, M.; Andreani, J.; Duflot, I.; Boxberger, M.; Bideau, M.L.; Mosnier, J.; Jardot, P.; Fonta, I.; Rolland, C.; Bogreau, H.; et al. Methylene blue inhibits the replication of SARS-CoV-2 in vitro. Int. J. Antimicrob. Agents 2020, 56, 106202. [Google Scholar] [CrossRef] [PubMed]
- Gendrot, M.; Jardot, P.; Delandre, O.; Boxberger, M.; Andreani, J.; Duflot, I.; Le Bideau, M.; Mosnier, J.; Fonta, I.; Hutter, S.; et al. In vitro evaluation of the antiviral activity of methylene blue alone or in combination against SARS-CoV-2. J. Clin. Med. 2021, 10, 3007. [Google Scholar] [CrossRef]
- Ellinger, B.; Bojkova, D.; Zaliani, A.; Cinatl, J.; Claussen, C.; Westhaus, S.; Keminer, O.; Reinshagen, J.; Kuzikov, M.; Wolf, M.; et al. A SARS-CoV-2 cytopathicity dataset generated by high-content screening of a large drug repurposing collection. Sci. Data 2021, 8, 70. [Google Scholar] [CrossRef]
- Xiong, H.L.; Cao, J.L.; Shen, C.G.; Ma, J.; Qiao, X.Y.; Shi, T.S.; Ge, S.X.; Ye, H.M.; Zhang, J.; Yuan, Q.; et al. Several FDA-approved drugs effectively inhibit SARS-CoV-2 infection in vitro. Front. Pharmacol. 2021, 11, 609592. [Google Scholar] [CrossRef]
- Murer, L.; Volle, R.; Andriasyan, V.; Petkidis, A.; Gomez-Gonzalez, A.; Yang, L.; Meili, N.; Suomalainen, M.; Bauer, M.; Sequeira, D.; et al. Identification of broad anti-coronavirus chemical agents for repurposing against SARS-CoV-2 and variants of concern. Curr. Res. Vir. Sci. 2022, 3, 100019. [Google Scholar] [CrossRef]
- Walter-Sack, I.; Rengelshausen, J.; Oberwittler, H.; Burhenne, J.; Mueller, O.; Meissner, P.; Mikus, G. High absolute bioavailability of methylene blue given as an aqueous oral formulation. Eur. J. Clin. Pharmacol. 2009, 65, 179–189. [Google Scholar] [CrossRef]
- Baddeley, T.C.; McCaffrey, J.; Storey, J.M.; Cheung, J.K.; Melis, V.; Horsley, D.; Harrington, C.R.; Wischik, C.M. Complex disposition of methylthioninium redox forms determines efficacy in tau aggregation inhibitor therapy for Alzheimer’s disease. J. Pharmacol. Exp. Ther. 2015, 352, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, L.; Margolles-Clark, E.; Song, Y.; Buchwald, P. The food colorant erythrosine is a promiscuous protein-protein interaction inhibitor. Biochem. Pharmacol. 2011, 81, 810–818. [Google Scholar] [CrossRef] [PubMed]
- Scigliano, G.; Scigliano, G.A. Acute respiratory distress syndrome from COVID-19: A perfect storm from free radicals? Proposal for a new treatment. Med. Hypotheses 2020, 144, 110120. [Google Scholar] [CrossRef] [PubMed]
- Scigliano, G.; Scigliano, G.A. Methylene blue in COVID-19. Med. Hypotheses 2021, 146, 110455. [Google Scholar] [CrossRef] [PubMed]
- Dabholkar, N.; Gorantla, S.; Dubey, S.K.; Alexander, A.; Taliyan, R.; Singhvi, G. Repurposing methylene blue in the management of COVID-19: Mechanistic aspects and clinical investigations. Biomed. Pharmacother. 2021, 142, 112023. [Google Scholar] [CrossRef] [PubMed]
- Tobin, M.J.; Laghi, F.; Jubran, A. Why COVID-19 silent hypoxemia is baffling to physicians. Am. J. Respir. Crit. Care Med. 2020, 202, 356–360. [Google Scholar] [CrossRef] [PubMed]
- Henry, M.; Summa, M.; Patrick, L.; Schwartz, L. A cohort of cancer patients with no reported cases of SARS-CoV-2 infection: The possible preventive role of methylene blue. Substantia 2020, 4, 888. [Google Scholar] [CrossRef]
- Alamdari, D.H.; Moghaddam, A.B.; Amini, S.; Keramati, M.R.; Zarmehri, A.M.; Alamdari, A.H.; Damsaz, M.; Banpour, H.; Yarahmadi, A.; Koliakos, G. Application of methylene blue-vitamin C-N-acetyl cysteine for treatment of critically ill COVID-19 patients, report of a phase-I clinical trial. Eur. J. Pharmacol. 2020, 173494. [Google Scholar] [CrossRef]
- Alamdari, D.H.; Lotfabadi, S.H.; Moghaddam, A.B.; Safari, H.; Mozdourian, M.; Javidarabshahi, Z.; Peivandi-Yazdi, A.; Ali-Zeraati, A.; Sedaghat, A.; Poursadegh, F.; et al. Methylene blue for treatment of hospitalized COVID-19 patients: A randomized, controlled, open-label clinical trial, phase 2. Rev. De Investig. Clínica 2021, 73, 190–198. [Google Scholar] [CrossRef]
- Alamdari, D.H.; Lotfabadi, S.H.; Darban, B.M.; Agheli-Rad, M.; Saadatian, S.; Hashemi, S.H.; Ahmadabadi, F.B.; Morovatdar, N.; Arastoo, M.; Bhushan, B. Methylene blue for treatment of hospitalized COVID-19 patients, randomized, controlled, open-label clinical trial, Phase 3. Aristotle Biomed. J. 2021, 3, 12–18. Available online: https://ejournals.lib.auth.gr/ABJ/article/view/8158 (accessed on 7 February 2022).
- Mahale, N.; Godavarthy, P.; Marreddy, S.; Gokhale, S.D.; Funde, P.; Rajhans, P.A.; Akole, P.V.; Pawar, B.; Bhurke, B.; Dalvi, P.; et al. Intravenous methylene blue as a rescue therapy in the management of refractory hypoxia in COVID-19 ARDS patients: A case series. Indian J. Crit. Care Med. 2021, 25, 934–938. [Google Scholar] [CrossRef] [PubMed]
- Golwalkar, D. Treatment for COVID-19 Using Methylene Blue. Medium. 2020. Available online: https://medium.com/@dr.deepak.golwalkar/treatment-for-covid-19-using-methylene-blue-d23fc5a31a4d (accessed on 21 August 2020).
- Patidar, V.; Sharma, A.; Bhoraskar, S.; Tripathi, A.P.; Dhaneriya, S. The role of nebulized methylene blue (NMB) in the management of COVID-19 cases: An observational study. Int. J. Med. Arts 2022, 4, 2129–2132. [Google Scholar] [CrossRef]
- Bawaskar, H.S.; Bawaskar, P.H. Role of methylene blue in the management of mild, moderate and severe COVID-19 disease. J. Fam. Med. Prim. Care 2022, 11, 812–814. [Google Scholar] [CrossRef] [PubMed]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Buchwald, P. A single unified model for fitting simple to complex receptor response data. Sci. Rep. 2020, 10, 13386. [Google Scholar] [CrossRef]
- Michel, M.C.; Murphy, T.J.; Motulsky, H.J. New author guidelines for displaying data and reporting data analysis and statistical methods in experimental biology. J. Pharmacol. Exp. Ther. 2020, 372, 136–147. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Assay | Cell Line | IC50 (μM) | Comment | Reference |
|---|---|---|---|---|
| PPI inhibition (SARS-CoV-2-S—hACE2) | N/A | 1–5 | Including VoCs (e.g., delta) | [23] and this work (VoC) |
| SARS-CoV-2 pseudovirus cell entry | HEK293T | 3–5 | Including VoCs (e.g., delta) | [23] and this work (VoC) |
| SARS-CoV-2 antiviral activity | Vero E6 | 1.7 | Inhibition of SARS-CoV-2 (B.1.5) replication | This work |
| Binding to SARS-CoV-2 S RBD | N/A | 0.36 | Bio-layer interferometry | [35] |
| SARS-CoV-2 antiviral activity | Vero E6 | ~0.3 | Also active against influenza virus H1N1 | [36] |
| SARS-CoV-2 antiviral activity | Vero E6 | 0.4–1.1 | clinically isolated SARS-CoV-2 strains (IHUMI-3 and IHUMI-6) | [37,38] |
| SARS-CoV-2 cytopathicity | Caco-2 | 2.03 | HTS hit, confirmed in concentration response (Supp Info in Res Square preprint) | [39] |
| VSV-SARS-CoV-2-Sdel18 pseudovirus entry | Vero E6 | 9 | HTS hit, confirmed in concentration response | [40] |
| Image-based multicycle replication assay (hCoV-229E) | Huh7 | 1.4 | HTS hit, confirmed in concentration response | [41] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chuang, S.-T.; Papp, H.; Kuczmog, A.; Eells, R.; Condor Capcha, J.M.; Shehadeh, L.A.; Jakab, F.; Buchwald, P. Methylene Blue Is a Nonspecific Protein–Protein Interaction Inhibitor with Potential for Repurposing as an Antiviral for COVID-19. Pharmaceuticals 2022, 15, 621. https://doi.org/10.3390/ph15050621
Chuang S-T, Papp H, Kuczmog A, Eells R, Condor Capcha JM, Shehadeh LA, Jakab F, Buchwald P. Methylene Blue Is a Nonspecific Protein–Protein Interaction Inhibitor with Potential for Repurposing as an Antiviral for COVID-19. Pharmaceuticals. 2022; 15(5):621. https://doi.org/10.3390/ph15050621
Chicago/Turabian StyleChuang, Sung-Ting, Henrietta Papp, Anett Kuczmog, Rebecca Eells, Jose M. Condor Capcha, Lina A. Shehadeh, Ferenc Jakab, and Peter Buchwald. 2022. "Methylene Blue Is a Nonspecific Protein–Protein Interaction Inhibitor with Potential for Repurposing as an Antiviral for COVID-19" Pharmaceuticals 15, no. 5: 621. https://doi.org/10.3390/ph15050621
APA StyleChuang, S. -T., Papp, H., Kuczmog, A., Eells, R., Condor Capcha, J. M., Shehadeh, L. A., Jakab, F., & Buchwald, P. (2022). Methylene Blue Is a Nonspecific Protein–Protein Interaction Inhibitor with Potential for Repurposing as an Antiviral for COVID-19. Pharmaceuticals, 15(5), 621. https://doi.org/10.3390/ph15050621