
Assessing Gene Expression Related to Cisplatin Resistance in Human Oral Squamous Cell Carcinoma Cell Lines



Abstract
:1. Introduction
2. Results
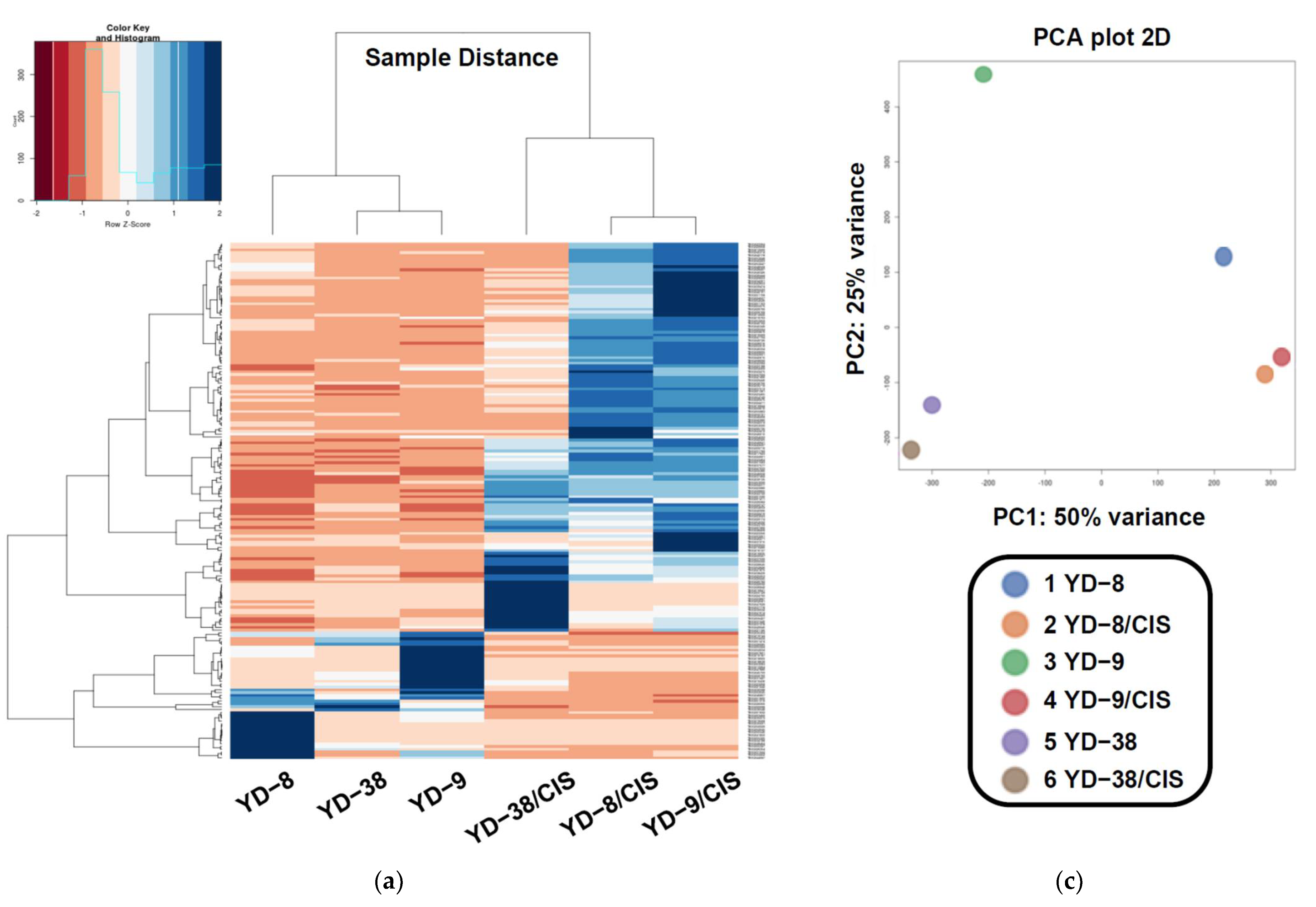
2.1. Seauencing and Unigene Assembly
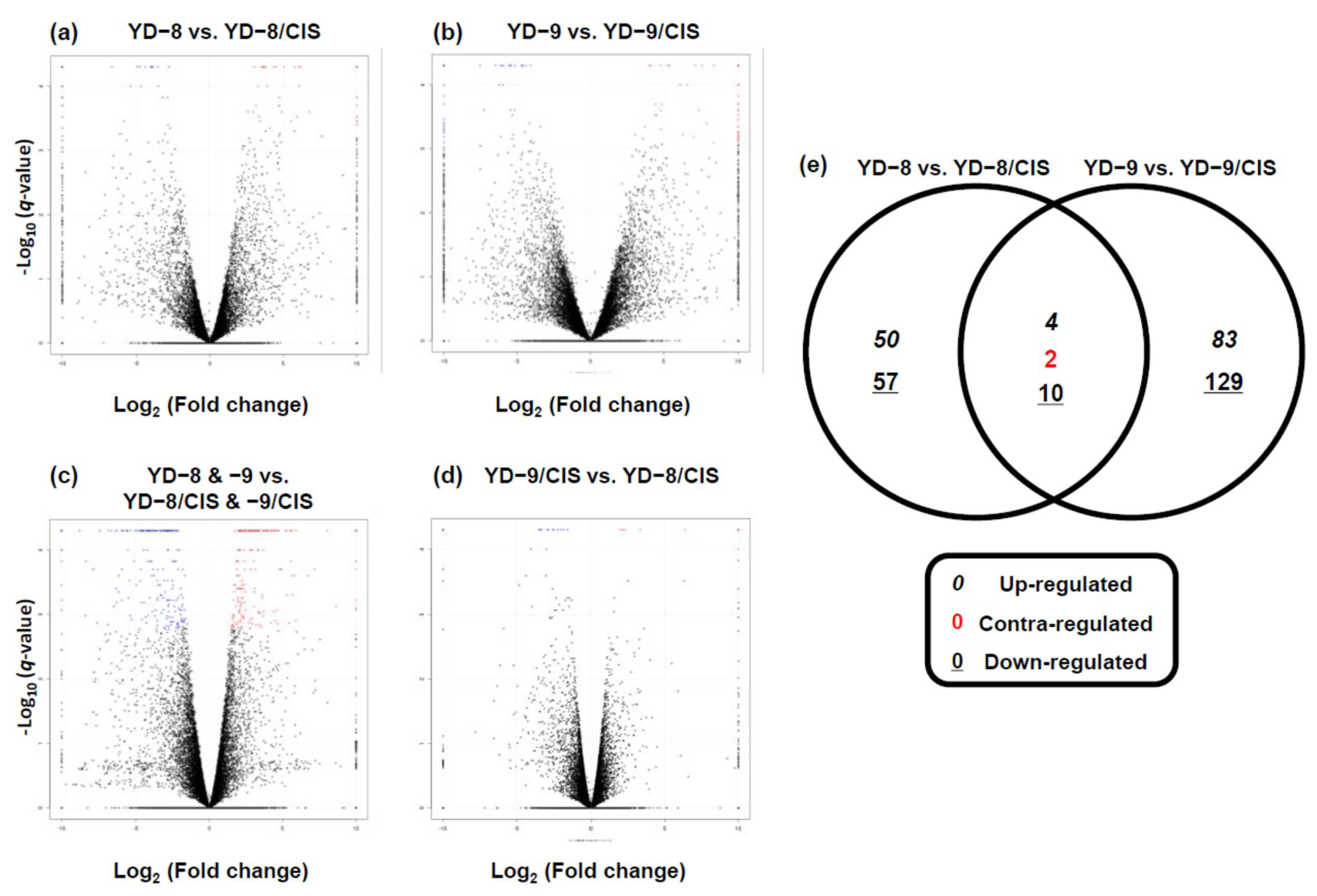
2.2. Analysis of Differentially Expressed Genes (DEGs)
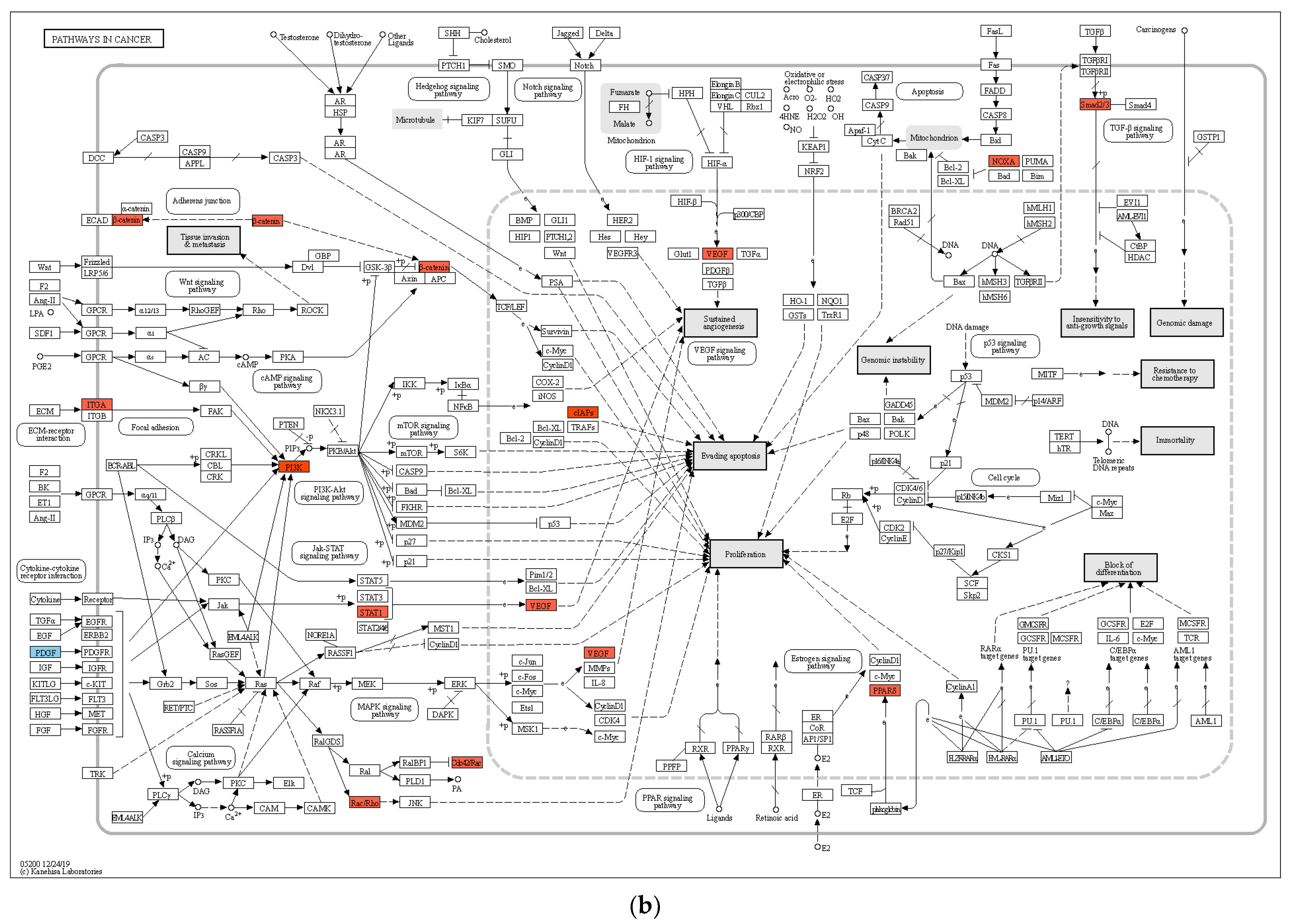
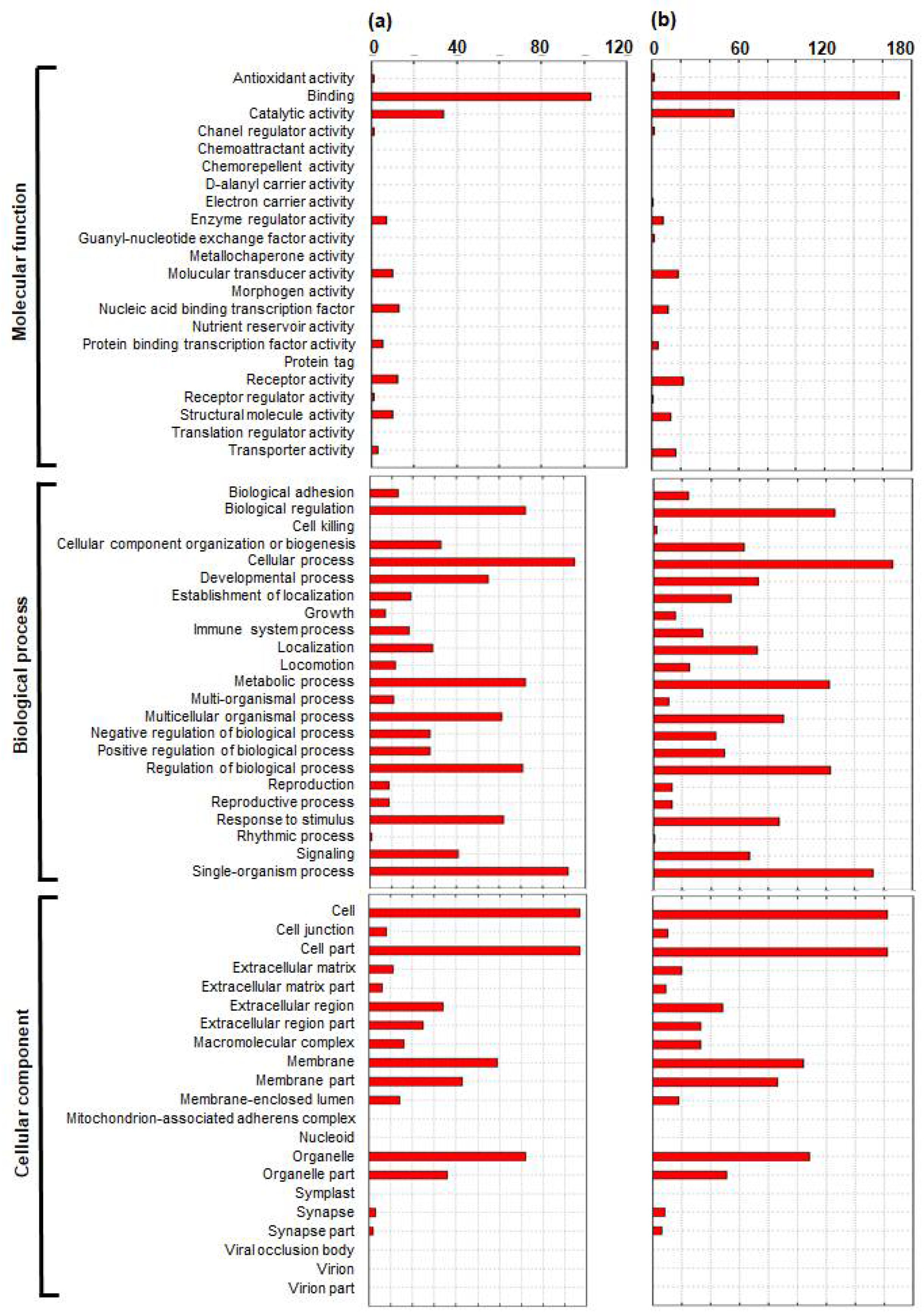
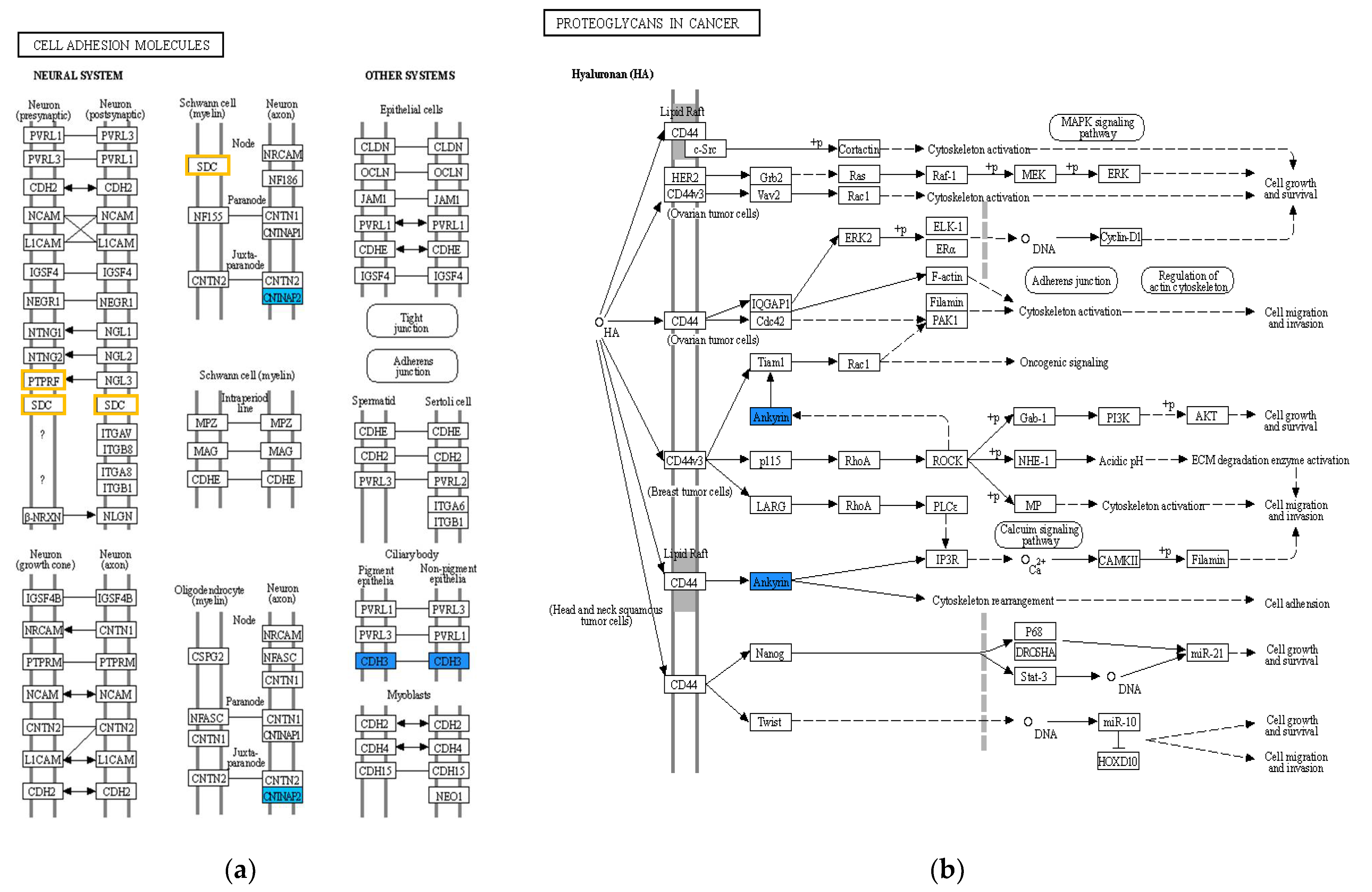
2.3. Functional Enrichment and Pathway Analyses of DEGs
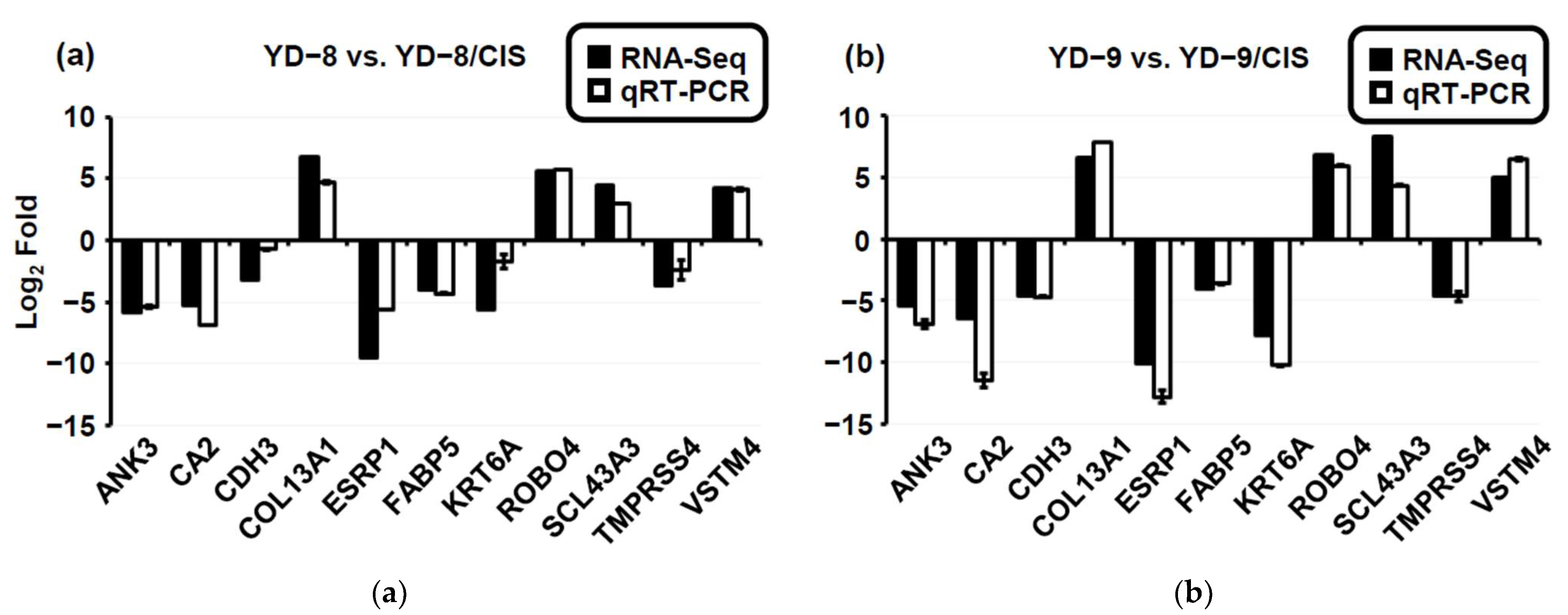
2.4. Validation of RNA-Seq Results by qRT-PCR
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Lines and Cell Culture
4.3. RNA Preparation
4.4. Generation of the Transcriptome Library and RNA Sequencing
4.5. Analysis of RNA Sequence Reads and Sequence Alignment
4.6. Gene Expression Estimation
4.7. Heatmap
4.8. PCA
4.9. Correlation Analysis
4.10. DEG Analysis
4.11. Experimental Validation via qRT-PCR
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Chattopadhyay, I.; Verma, M.; Panda, M. Role of Oral Microbiome Signatures in Diagnosis and Prognosis of Oral Cancer. Technol. Cancer Res. Treat. 2019, 18, 1533033819867354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.C.; Shen, Z.; Bavarian, R.; Yang, F.; Bhattacharya, A. Oral Cancer: Genetics and the Role of Precision Medicine. Surg. Oncol. Clin. N. Am. 2020, 29, 127–144. [Google Scholar] [CrossRef] [PubMed]
- Montero, P.H.; Patel, S.G. Cancer of the oral cavity. Surg. Oncol. Clin. N. Am. 2015, 24, 491–508. [Google Scholar] [CrossRef] [Green Version]
- Markopoulos, A.K. Current aspects on oral squamous cell carcinoma. Open Dent. J. 2012, 6, 126–130. [Google Scholar] [CrossRef]
- Sha, J.; Bai, Y.; Ngo, H.X.; Okui, T.; Kanno, T. Overview of Evidence-Based Chemotherapy for Oral Cancer: Focus on Drug Resistance Related to the Epithelial-Mesenchymal Transition. Biomolecules 2021, 11, 893. [Google Scholar] [CrossRef]
- Rivera, C. Essentials of oral cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 11884–11894. [Google Scholar]
- Mosaddad, S.A.; Beigi, K.; Doroodizadeh, T.; Haghnegahdar, M.; Golfeshan, F.; Ranjbar, R.; Tebyanian, H. Therapeutic applications of herbal/synthetic/bio-drug in oral cancer: An update. Eur. J. Pharmacol. 2021, 890, 173657. [Google Scholar] [CrossRef]
- Cheng, Y.; Li, S.; Gao, L.; Zhi, K.; Ren, W. The Molecular Basis and Therapeutic Aspects of Cisplatin Resistance in Oral Squamous Cell Carcinoma. Front. Oncol. 2021, 11, 761379. [Google Scholar] [CrossRef]
- Dasari, S.; Tchounwou, P.B. Cisplatin in cancer therapy: Molecular mechanisms of action. Eur. J. Pharmacol. 2014, 740, 364–378. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Zhang, Z.; Zhou, X.; Qiu, W.; Chen, F.; Chen, W. Identification of genes associated with cisplatin resistance in human oral squamous cell carcinoma cell line. BMC Cancer 2006, 6, 224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, D.W.; Pouliot, L.M.; Hall, M.D.; Gottesman, M.M. Cisplatin resistance: A cellular self-defense mechanism resulting from multiple epigenetic and genetic changes. Pharmacol. Rev. 2012, 64, 706–721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, H.S.; Kim, Y.-K.; Yun, P.-Y. Upregulation of MDR- and EMT-Related Molecules in Cisplatin-Resistant Human Oral Squamous Cell Carcinoma Cell Lines. Int. J. Mol. Sci. 2019, 20, 3034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, D.S.; Lan, H.Y.; Huang, C.H.; Tai, S.K.; Chang, S.Y.; Tsai, T.L.; Chang, C.C.; Tzeng, C.H.; Wu, K.J.; Kao, J.Y.; et al. Regulation of excision repair cross-complementation group 1 by Snail contributes to cisplatin resistance in head and neck cancer. Clin. Cancer Res. 2010, 16, 4561–4571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, H.S.; Kim, Y.-K.; Hwang, K.-G.; Yun, P.-Y. Increased FOXM1 Expression by Cisplatin Inhibits Paclitaxel-Related Apoptosis in Cisplatin-Resistant Human Oral Squamous Cell Carcinoma (OSCC) Cell Lines. Int. J. Mol. Sci. 2020, 21, 8897. [Google Scholar] [CrossRef]
- Choi, H.S.; Kim, Y.-K.; Yun, P.-Y. Cisplatin Plus Cetuximab Inhibits Cisplatin-Resistant Human Oral Squamous Cell Carcinoma Cell Migration and Proliferation but Does Not Enhance Apoptosis. Int. J. Mol. Sci. 2021, 22, 8167. [Google Scholar] [CrossRef]
- Meyer, T.; Hart, I.R. Mechanisms of tumour metastasis. Eur. J. Cancer 1998, 34, 214–221. [Google Scholar] [CrossRef]
- Makrilia, N.; Kollias, A.; Manolopoulos, L.; Syrigos, K. Cell adhesion molecules: Role and clinical significance in cancer. Cancer Investig. 2009, 27, 1023–1037. [Google Scholar] [CrossRef]
- Harjunpää, H.; Llort Asens, M.; Guenther, C.; Fagerholm, S.C. Cell Adhesion Molecules and Their Roles and Regulation in the Immune and Tumor Microenvironment. Front. Immunol. 2019, 10, 1078. [Google Scholar] [CrossRef] [Green Version]
- Schwab, M. Cell Adhesion Molecules. In Encyclopedia of Cancer; V, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2008; pp. 572–576. [Google Scholar]
- Koukoulis, G.K.; Patriarca, C.; Gould, V.E. Adhesion molecules and tumor metastasis. Hum. Pathol. 1998, 29, 889–892. [Google Scholar] [CrossRef]
- Iozzo, R.V.; Sanderson, R.D. Proteoglycans in cancer biology, tumour microenvironment and angiogenesis. J. Cell. Mol. Med. 2011, 15, 1013–1031. [Google Scholar] [CrossRef] [PubMed]
- Nikitovic, D.; Berdiaki, A.; Spyridaki, I.; Krasanakis, T.; Tsatsakis, A.; Tzanakakis, G.N. Proteoglycans-Biomarkers and Targets in Cancer Therapy. Front. Endocrinol. 2018, 9, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, B.; Shim, J.S. Targeting Epithelial-Mesenchymal Transition (EMT) to Overcome Drug Resistance in Cancer. Molecules 2016, 21, 965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashrafizadeh, M.; Zarrabi, A.; Hushmandi, K.; Kalantari, M.; Mohammadinejad, R.; Javaheri, T.; Sethi, G. Association of the Epithelial-Mesenchymal Transition (EMT) with Cisplatin Resistance. Int. J. Mol. Sci. 2020, 21, 4002. [Google Scholar] [CrossRef]
- Wu, T.; Xiao, Z.; Li, Y.; Jiao, Z.; Liang, X.; Zhang, Y.; Liu, H.; Yang, A. CDH3 is associated with a poor prognosis by promoting the malignance and chemoresistance in oral squamous cell carcinoma. Asian J. Surg. 2022, in press. [CrossRef]
- Bourguignon, L.Y.W.; Shiina, M.; Li, J.-J. Chapter Ten—Hyaluronan–CD44 Interaction Promotes Oncogenic Signaling, microRNA Functions, Chemoresistance, and Radiation Resistance in Cancer Stem Cells Leading to Tumor Progression. In Advances in Cancer Research; Simpson, M.A., Heldin, P., Eds.; Academic Press: Cambridge, MA, USA, 2014; Volume 123, pp. 255–275. [Google Scholar]
- Lee, E.J.; Kim, J.; Lee, S.A.; Kim, E.J.; Chun, Y.C.; Ryu, M.H.; Yook, J.I. Characterization of newly established oral cancer cell lines derived from six squamous cell carcinoma and two mucoepidermoid carcinoma cells. Exp. Mol. Med. 2005, 37, 379–390. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Pachter, L.; Salzberg, S.L. TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 2009, 25, 1105–1111. [Google Scholar] [CrossRef]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [Green Version]
- Trapnell, C.; Roberts, A.; Goff, L.; Pertea, G.; Kim, D.; Kelley, D.R.; Pimentel, H.; Salzberg, S.L.; Rinn, J.L.; Pachter, L. Differential gene and transcript expression analysis of RNA-seq experiments with TopHat and Cufflinks. Nat. Protoc. 2012, 7, 562–578. [Google Scholar] [CrossRef] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Glaab, E.; Baudot, A.; Krasnogor, N.; Schneider, R.; Valencia, A. EnrichNet: Network-based gene set enrichment analysis. Bioinformatics 2012, 28, i451–i457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinderer, E.W., 3rd; Flight, R.M.; Dubey, R.; MacLeod, J.N.; Moseley, H.N. Advances in gene ontology utilization improve statistical power of annotation enrichment. PLoS ONE 2019, 14, e0220728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, R.A. On the Interpretation of χ2 from Contingency Tables, and the Calculation of P. J. R. Stat. Soc. 1922, 85, 87–94. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample (Name) | Total Reads (Raw) | Processed Reads (Cleaned) | Mapped Reads | Mapping Rate | Uniquely Mapped | Mapping Rate |
|---|---|---|---|---|---|---|
| YD-8 | 48,790,084 | 47,574,638 | 42,418,196 | 89.2% | 41,494,996 | 87.2% |
| YD-8/CIS | 54,379,620 | 53,296,460 | 47,104,007 | 88.4% | 46,142,562 | 86.6% |
| YD-9 | 49,933,926 | 48,870,688 | 40,886,333 | 83.7% | 39,607,099 | 81.0% |
| YD-9/CIS | 54,914,214 | 53,564,370 | 46,767,206 | 87.3% | 45,748,506 | 85.4% |
| YD-38 | 49,258,722 | 47,880,336 | 41,381,531 | 86.4% | 40,323,662 | 84.2% |
| YD-38/CIS | 54,601,488 | 53,489,836 | 42,157,460 | 78.8% | 41,168,565 | 77.0% |
| Scale of Correlation Coefficient | Value |
|---|---|
| 0.00 < r ≤ 0.70 | Very low correlation |
| 0.70 < r ≤ 0.80 | Low correlation |
| 0.80 < r ≤ 0.92 | Moderate correlation |
| 0.92 < r ≤ 1.00 | Very high correlation |
| YD-8 | YD-8/CIS | YD-9 | YD-9/CIS | YD-38 | YD-38/CIS | |
| 0.856 | 0.794 | 0.833 | 0.825 | 0.765 | YD-8 | |
| 0.733 | 0.960 | 0.805 | 0.787 | YD-8/CIS | ||
| 0.717 | 0.842 | 0.803 | YD-9 | |||
| 0.790 | 0.779 | YD-9/CIS | ||||
| 0.896 | YD-38 | |||||
| YD-38/CIS |
| Group 1 | Group 2 | Genes | ||
|---|---|---|---|---|
| Up | Down | Sum | ||
| YD-8 | YD-8/CIS | 55 | 68 | 123 |
| YD-9 | YD-9/CIS | 88 | 140 | 228 |
| YD-8 & -9 | YD-8/CIS & -9/CIS | 188 | 242 | 430 |
| YD-9/CIS | YD-8/CIS | 12 | 23 | 35 |
| Gene Symbol | Gene Name | Pathway |
|---|---|---|
| ADGRL2 | Adhesion G Protein-Coupled Receptor L2 | |
| ANK3 | Ankyrin 3 | hsa05205 |
| AUTS2 | Autism susceptibility candidate 2 | |
| CA2 | Carbonic anhydrase II | hsa04976, hsa00910, hsa04964 hsa04966, hsa04971, hsa04972 |
| CDH3 | Cadherin 3 | hsa04514 |
| CNTNAP2 | Contactin-associated protein-like 2 | hsa04514 |
| COL13A1 | Collagen, type XIII, alpha 1 | hsa04974 |
| ESRP1 | Epithelial Splicing Regulatory Protein 1 | |
| FABP5 | Fatty acid binding protein 5 | hsa03320 |
| GRHL2 | Grainyhead-like 2 | |
| KRT6A | Keratin 6A | |
| ROBO4 | Roundabout guidance receptor 4 | |
| SDC2 | Syndecan 2 | hsa04514, hsa05144 hsa05205, hsa05418 |
| SLC43A3 | Solute carrier family 43 member 3 | |
| TMPRSS4 | Transmembrane protease serine 4 | hsa05164 |
| Pathway Entry | Name |
|---|---|
| hsa00910 | Nitrogen metabolism |
| hsa03320 | PPAR signaling pathway |
| hsa04514 | Cell adhesion molecules |
| hsa04964 | Proximal tubule bicarbonate reclamation |
| hsa04966 | Collecting duct acid secretion |
| hsa04971 | Gastric acid secretion |
| hsa04972 | Pancreatic secretion |
| hsa04974 | Protein digestion and absorption |
| hsa04976 | Bile secretion |
| hsa05144 | Malaria |
| hsa05164 | Influenza A |
| hsa05205 | Proteoglycans in cancer |
| hsa05418 | Fluid shear stress and atherosclerosis |
| Gene Name | Primer Sequence | Length | References | |
|---|---|---|---|---|
| ANK3 | F | AAAGGACTGCCTCAAACAGCGG | 22 | Origene (Gene ID: 288) |
| R | CTAAGGATGCGAAGCTCTGTCG | 22 | ||
| CA2 | F | CAATGGTCATGCTTTCAACG | 20 | Clin Cancer Res. 2005 Nov 15;11(22):8201–8207. |
| R | TCCATCAAGTGAACCCCAGT | 20 | doi: 10.1158/1078-0432.CCR-05-0816. | |
| CDH3 | F | CCCCCAGAAGTACGAGGCCCA | 20 | Anat Cell Biol. 2010 Jun;43(2):110–117. |
| R | ACGCCACGCTGGTGAGTTGG | 21 | doi: 10.5115/acb.2010.43.2.110 | |
| COL13A1 | F | CAAAGGGAGAAGCAGGTGTC | 20 | Int J Mol Sci. 2019 Oct; 20(19): 4890. |
| R | TCACTGGAGAGCCTCATTGAT | 21 | doi: 10.3390/ijms20194890. | |
| ESRP1 | F | TCCTGCTGTTCTGGAAAGTCG | 21 | Cancer Lett. 2011 Jan 1;300(1):66–78. |
| R | TCCGGTCTAACTAGCACTTCGTG | 23 | doi: 10.1016/j.canlet.2010.09.007. | |
| FABP5 | F | GCTGATGGCAGAAAAACTCAGA | 22 | Oncotarget. 2018 Aug 3; 9(60): 31753–31770. |
| R | CCTGATGCTGAACCAATGCA | 20 | doi: 10.18632/oncotarget.25878. | |
| KRT6A | F | TCACCGTCAACCAGAGTCTC | 20 | Mol Med Rep. 2019 May;19(5):3477–3484. |
| R | GAACCTTGTTCTGCTGCTCC | 20 | doi: 10.3892/mmr.2019.10055. | |
| ROBO4 | F | GACACTTGGCGTTCCACCTC | 20 | BMC Cancer. 2008 Dec 29;8:392. |
| R | AGAGCAAGGAGCGACGACAG | 20 | doi: 10.1186/1471-2407-8-392. | |
| SLC43A3 | F | CACCGCCACACTCATCATAG | 20 | J Pharm Sci. 2020 Aug;109(8):2622–2628. |
| R | GGTGTTGGCCAAATAGGTTC | 20 | doi: 10.1016/j.xphs.2020.04.013. | |
| TMPRSS4 | F | CCGATGTGTTCAACTGGAAG | 20 | Br J Cancer. 2011 Nov 8;105(10):1608–1614. |
| R | GAGAAAGTGAGTGGGAACTG | 20 | doi: 10.1038/bjc.2011.432. | |
| VSTM4 | F | TGTCACTAGCGTGACCAGCTTG | 22 | Origene (Gene ID: 196740) |
| R | CAGCTTCGGTTTATGGAACGTGG | 23 | ||
| GAPDH | F | AATCCCATCACCATCTTCCA | 20 | Cell Mol Life Sci. 2016 Sep 11;73:1067–1084. |
| R | TGGACTCCACGACGTACTCA | 20 | doi: 10.1007/s00018-015-2036-6. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, H.S.; Kim, Y.-K.; Yun, P.-Y. Assessing Gene Expression Related to Cisplatin Resistance in Human Oral Squamous Cell Carcinoma Cell Lines. Pharmaceuticals 2022, 15, 704. https://doi.org/10.3390/ph15060704
Choi HS, Kim Y-K, Yun P-Y. Assessing Gene Expression Related to Cisplatin Resistance in Human Oral Squamous Cell Carcinoma Cell Lines. Pharmaceuticals. 2022; 15(6):704. https://doi.org/10.3390/ph15060704
Chicago/Turabian StyleChoi, Hyeong Sim, Young-Kyun Kim, and Pil-Young Yun. 2022. "Assessing Gene Expression Related to Cisplatin Resistance in Human Oral Squamous Cell Carcinoma Cell Lines" Pharmaceuticals 15, no. 6: 704. https://doi.org/10.3390/ph15060704
APA StyleChoi, H. S., Kim, Y. -K., & Yun, P. -Y. (2022). Assessing Gene Expression Related to Cisplatin Resistance in Human Oral Squamous Cell Carcinoma Cell Lines. Pharmaceuticals, 15(6), 704. https://doi.org/10.3390/ph15060704