
Exploring the Anticancer Effects of Brominated Plastoquinone Analogs with Promising Cytotoxic Activity in MCF-7 Breast Cancer Cells via Cell Cycle Arrest and Oxidative Stress Induction

 , , , ,
, , , ,  , and
, and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Design Strategy
2.2. Biological Activities
2.2.1. Preliminary Screening of the In Vitro Antiproliferative Activity
2.2.2. In Vitro Antiproliferative Activity at the One-Dose Assay
2.2.3. In Vitro Antiproliferative Activity at Five-Dose Assay
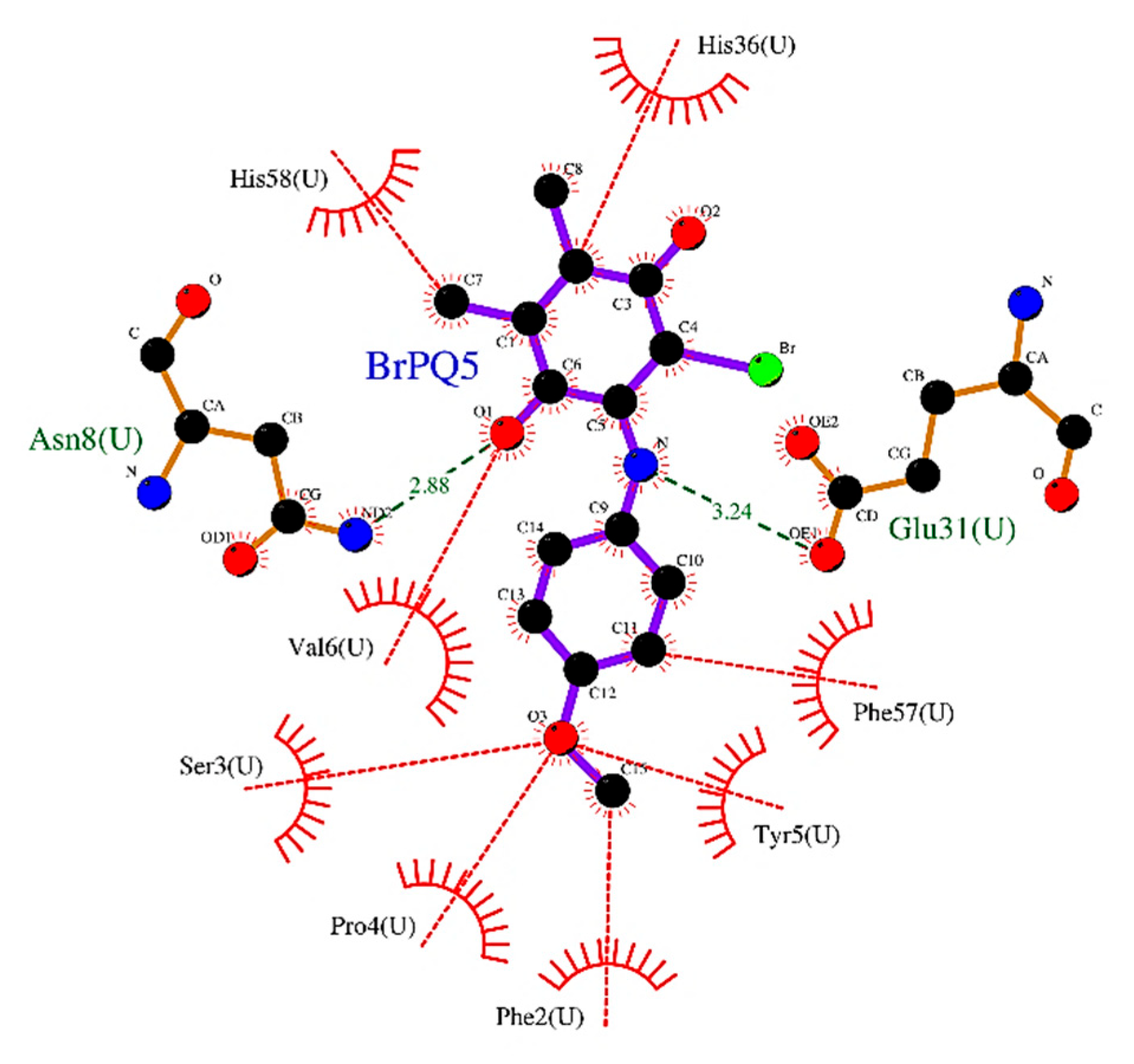
2.2.4. in silico Study
2.2.5. In Vitro Anticancer Activity
3. Conclusions
4. Materials and Methods
4.1. Biological Evaluation
4.1.1. In Vitro Antiproliferative Activity at a One-Dose Concentration by NCI
4.1.2. In Vitro Antiproliferative Activity at a Five-Dose Concentration by NCI
4.1.3. Cell Culture and Cytotoxicity Assay
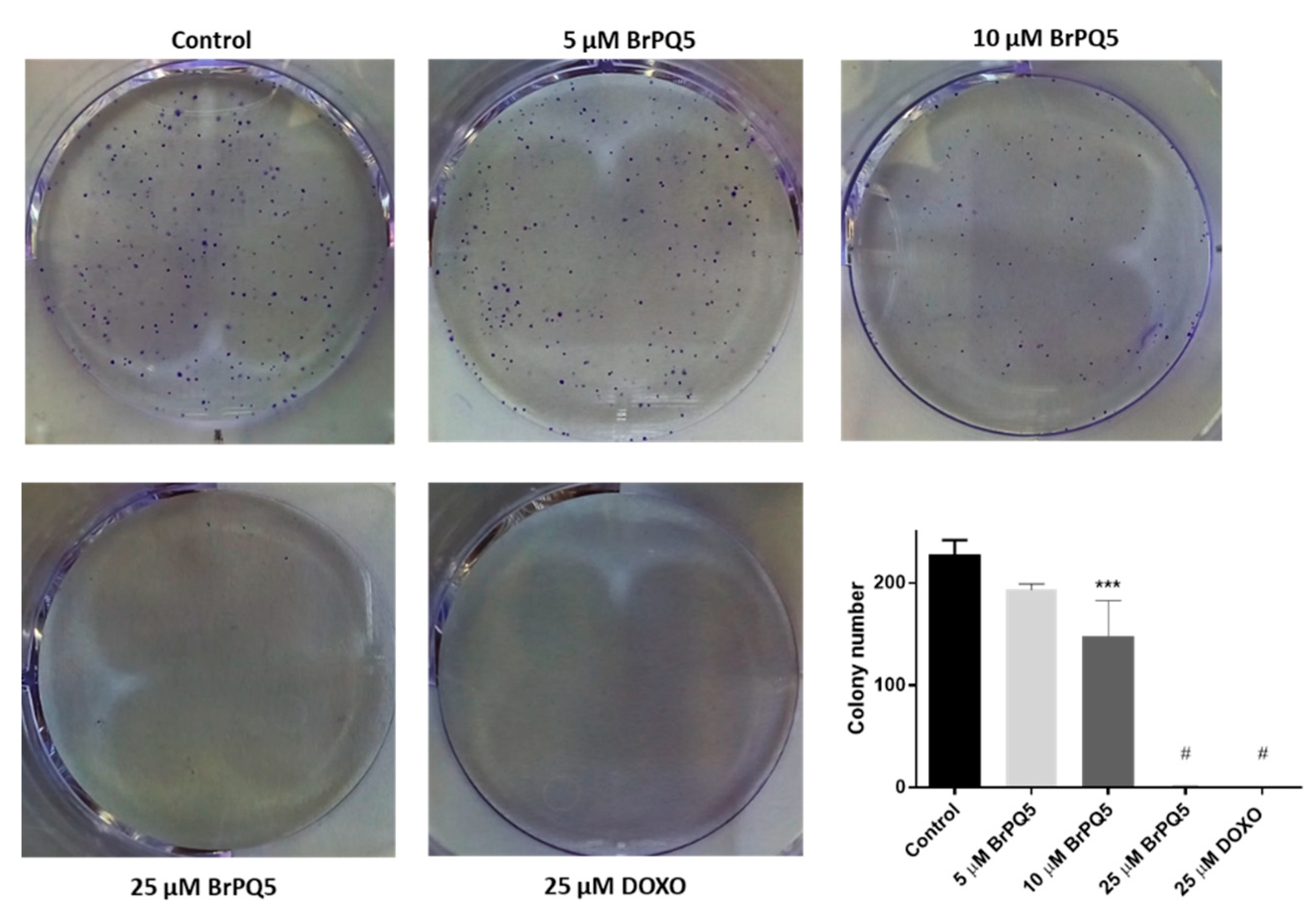
4.1.4. Colony Formation Assay
4.1.5. Oxidative Stress Evaluation
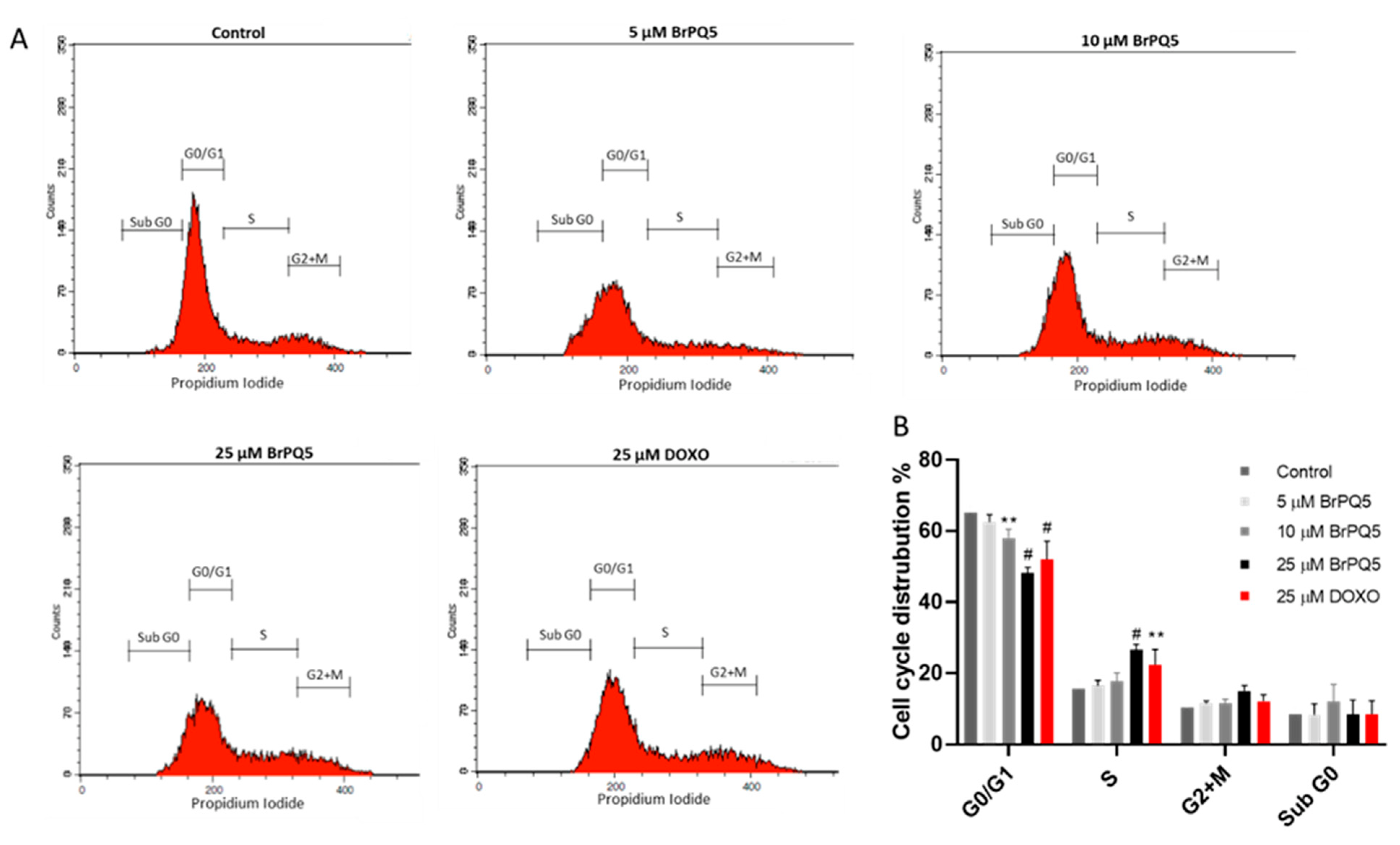
4.1.6. Cell Cycle Analysis
4.1.7. Apoptosis and Necrosis Analysis
4.1.8. Proteasome Activity
4.1.9. Statistics
4.2. In Silico Study
- (i)
- Protein preparation: Coordinates of human 20S proteasome subunits beta-1 (chain M), beta-2 (chain K), and beta-5 (chain L) were manually extracted from the X-ray crystal structure of the human 20S proteasome in a complex with carfilzomib (PDB:4R67) [56]. The coordinates of subunits beta-1 and beta-2 were then superimposed on beta-5 and were rewritten using UCSF chimera [57]. This allowed us to use the coordinates of the co-crystallized ligand (3BV) of subunit beta-5 to be used for all three to specify the grid box. Then, all the three extracted chains were prepared for docking through the pdb2pqr server. Further, the non-polar hydrogens were merged, the AD4 atom type was assigned, and gasteiger charges were added using MGLTools-1.5.7. It was then saved as respective_protein.pdbqt.
- (ii)
- Ligand preparation: The structure of BrPQ5 was sketched in Chemdraw-19, 3D geometry optimized, energy minimized, and saved as ligand.pdb. Torsion and charge were assigned to the ligand and then saved as ligand.pdbqt using MGLTools-1.5.7.
- (iii)
- Docking preparation, run, analysis: (a) Generation of grid map files: the grid over the binding site on respective_protein (*.pdbqt) was generated using the coordinates of carfilzomib (3BV) bound to subunit beta-5 (chain L). The center of the grid was specified as the center of the protein-bound ligand with a box dimension of 40:42:40 and grid spacing of 0.375 Å. Map types were set considering the atom types in the BrPQ5. The grid parameter file was then saved as respective_protein.gpf and used to generate map files using the autogrid4 execution file; (b) Docking: the docking parameter file for each subunit was then written for BrPQ5 (respective_protein_BrPQ5.dpf) with 100 hybrid GA-LS runs, 150 population size, 2,500,000 energy evaluations, and 27,000 generations. The generated map files and the *.dpf files docking simulation were performed using the autodock4 execution file. (c) Analysis: the top scoring conformer in the largest cluster of the respective docking log file (*.dlg) was then picked for interaction analysis, and the 3D interaction plot was then saved as *.png (in Supplementary File). All these steps were carried out using MGLTools-1.5.7. Further 2D plots were also generated for each complex to better understand protein–ligand interactions using LigPlot+ [58]. The coordinates of respective_proteins, co-crystallized ligand and BrPQ5, respective_protein.gpf, and respective_protein_BrPQ5.dpf files used in the study are provided in the Supplementary File.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khuwaja, G.A.; Abu-Rezq, A.N. Bimodal breast cancer classification system. Pattern Anal. Appl. 2004, 7, 235–242. [Google Scholar] [CrossRef]
- Wu, X.; Chen, G.; Lu, J.; Zhu, W.; Qiu, J.; Chen, J.; Xie, S.; Zhuo, S.; Yan, J. Label-Free Detection of Breast Masses Using Multiphoton Microscopy. PLoS ONE 2013, 8, e65933. [Google Scholar] [CrossRef] [PubMed]
- Breast Cancer. Available online: https://www.who.int/news-room/fact-sheets/detail/breast-cancer (accessed on 1 September 2021).
- Breast Cancer Now Most Common form of Cancer: WHO Taking Action. Available online: https://www.who.int/news/item/03-02-2021-breast-cancer-now-most-common-form-of-cancer-who-taking-action (accessed on 1 September 2021).
- Yoshimaru, T.; Nakamura, Y.; Katagiri, T. Functional genomics for breast cancer drug target discovery. J. Hum. Genet. 2021, 66, 927–935. [Google Scholar] [CrossRef] [PubMed]
- Al-Mahmood, S.; Sapiezynski, J.; Garbuzenko, O.B.; Minko, T. Metastatic and triple-negative breast cancer: Challenges and treatment options. Drug Deliv. Transl. Res. 2018, 8, 1483–1507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hebar, A.; Valent, P.; Selzer, E. The impact of molecular targets in cancer drug development: Major hurdles and future strategies. Expert Rev. Clin. Pharmacol. 2013, 6, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Gewirtz, D.A.; Bristol, M.L.; Yalowich, J.C. Toxicity issues in cancer drug development. Curr. Opin. Investig. Drugs 2010, 11, 612–614. [Google Scholar]
- Wellington, K.W.; Kolesnikova, N.I.; Nyoka, N.B.P.; McGaw, L.J. Investigation of the antimicrobial and anticancer activity of aminonaphthoquinones. Drug Dev. Res. 2019, 80, 138–146. [Google Scholar] [CrossRef] [Green Version]
- Santoso, K.T.; Menorca, A.; Cheung, C.Y.; Cook, G.M.; Stocker, B.L.; Timmer, M.S.M. The synthesis and evaluation of quinolinequinones as anti-mycobacterial agents. Bioorg. Med. Chem. 2019, 27, 3532–3545. [Google Scholar] [CrossRef]
- Wellington, K.W. Understanding cancer and the anticancer activities of naphthoquinones—A review. Rsc. Adv. 2015, 5, 20309–20338. [Google Scholar] [CrossRef]
- Defant, A.; Mancini, I. Design, Synthesis and Cancer Cell Growth Inhibition Evaluation of New Aminoquinone Hybrid Molecules. Molecules 2019, 24, 2224. [Google Scholar] [CrossRef] [Green Version]
- Nain-Perez, A.; Barbosa, L.C.A.; Rodriguez-Hernandez, D.; Kramell, A.E.; Heller, L.; Csuk, R. Natural abenquines and synthetic analogues: Preliminary exploration of their cytotoxic activity. Bioorg. Med. Chem. Lett. 2017, 27, 1141–1144. [Google Scholar] [CrossRef]
- Nain-Perez, A.; Barbosa, L.C.A.; Maltha, C.R.A.; Forlani, G. First total synthesis and phytotoxic activity of streptomyces sp metabolites abenquines. Tetrahedron Lett. 2016, 57, 1811–1814. [Google Scholar] [CrossRef]
- Aminin, D.; Polonik, S. 1,4-Naphthoquinones: Some Biological Properties and Application. Chem. Pharm. Bull. 2020, 68, 46–57. [Google Scholar] [CrossRef] [Green Version]
- Kruschel, R.D.; Buzid, A.; Khandavilli, U.B.R.; Lawrence, S.E.; Glennon, J.D.; McCarthy, F.O. Isoquinolinequinone N-oxides as anticancer agents effective against drug resistant cell lines. Org. Biomol. Chem. 2020, 18, 557–568. [Google Scholar] [CrossRef]
- Ravichandiran, P.; Maslyk, M.; Sheet, S.; Janeczko, M.; Premnath, D.; Kim, A.R.; Park, B.H.; Han, M.K.; Yoo, D.J. Synthesis and Antimicrobial Evaluation of 1,4-Naphthoquinone Derivatives as Potential Antibacterial Agents. Chemistryopen 2019, 8, 589–600. [Google Scholar] [CrossRef] [Green Version]
- Ravichandiran, P.; Sheet, S.; Premnath, D.; Kim, A.R.; Yoo, D.J. 1,4-Naphthoquinone Analogues: Potent Antibacterial Agents and Mode of Action Evaluation. Molecules 2019, 24, 1437. [Google Scholar] [CrossRef] [Green Version]
- Ravichandiran, P.; Subramaniyan, S.A.; Kim, S.Y.; Kim, J.S.; Park, B.H.; Shim, K.S.; Yoo, D.J. Synthesis and Anticancer Evaluation of 1,4-Naphthoquinone Derivatives Containing a Phenylaminosulfanyl Moiety. Chemmedchem 2019, 14, 532–544. [Google Scholar] [CrossRef] [Green Version]
- Bayrak, N.; Ciftci, H.I.; Yildiz, M.; Yildirim, H.; Sever, B.; Tateishi, H.; Otsuka, M.; Fujita, M.; Tuyun, A.F. Structure based design, synthesis, and evaluation of anti-CML activity of the quinolinequinones as LY83583 analogs. Chem. Biol. Interact. 2021, 345, 109555. [Google Scholar] [CrossRef]
- Bayrak, N.; Yildirim, H.; Yildiz, M.; Radwan, M.O.; Otsuka, M.; Fujita, M.; Ciftci, H.I.; Tuyun, A.F. A novel series of chlorinated plastoquinone analogs: Design, synthesis, and evaluation of anticancer activity. Chem. Biol. Drug Des. 2020, 95, 343–354. [Google Scholar] [CrossRef]
- Ciftci, H.I.; Bayrak, N.; Yildirim, H.; Yildiz, M.; Radwan, M.O.; Otsuka, M.; Fujita, M.; Tuyun, A.F. Discovery and structure-activity relationship of plastoquinone analogs as anticancer agents against chronic myelogenous leukemia cells. Arch. Der Pharm. 2019, 352, e1900170. [Google Scholar] [CrossRef]
- Mataraci-Kara, E.; Bayrak, N.; Yildirim, H.; Yildiz, M.; Ataman, M.; Ozbek-Celik, B.; Tuyun, A.F. Plastoquinone analogs: A potential antimicrobial lead structure intensely suppressing staphylococcus epidermidis and candida albicans growth. Med. Chem. Res. 2021, 30, 1728–1737. [Google Scholar] [CrossRef]
- Tuyun, A.F.; Yildiz, M.; Bayrak, N.; Yildirim, H.; Kara, E.M.; Jannuzzi, A.T.; Celik, B.O. Discovery of a new family of heterocyclic amine linked plastoquinone analogs for antimicrobial evaluation. Drug Dev. Res. 2019, 80, 1098–1109. [Google Scholar] [CrossRef] [PubMed]
- Yildiz, M.; Bayrak, N.; Yildirim, H.; Mataraci-Kara, E.; Shilkar, D.; Jayaprakash, V.; Tuyun, A.F. Exploration of brominated Plastoquinone analogs: Discovery and structure-activity relationships of small antimicrobial lead molecules. Bioorg. Chem. 2021, 116, 105316. [Google Scholar] [CrossRef] [PubMed]
- Kara, E.M.; Bayrak, N.; Yildirim, H.; Yildiz, M.; Celik, B.O.; Tuyun, A.F. Chlorinated plastoquinone analogs that inhibit staphylococcus epidermidis and candida albicans growth. Folia Microbiol. 2020, 65, 785–795. [Google Scholar] [CrossRef] [PubMed]
- Jannuzzi, A.T.; Yıldız, M.; Bayrak, N.; Yıldırım, H.; Shilkar, D.; Jayaprakash, V.; Tuyun, A.F. Anticancer agents based on Plastoquinone analogs with N-phenylpiperazine: Structure-activity relationship and mechanism of action in breast cancer cells. Chem. Biol. Interact. 2021, 349, 109673. [Google Scholar] [CrossRef]
- Bayrak, N.; Yildiz, M.; Yildirim, H.; Kara, E.M.; Celik, B.O.; Tuyun, A.F. Brominated plastoquinone analogs: Synthesis, structural characterization, and biological evaluation. J. Mol. Struct. 2020, 1219, 128560. [Google Scholar] [CrossRef]
- Moloney, J.N.; Cotter, T.G. ROS signalling in the biology of cancer. Semin. Cell Dev. Biol. 2018, 80, 50–64. [Google Scholar] [CrossRef]
- Boler, J.; Pardini, R.; Mustafa, H.T.; Folkers, K.; Dilley, R.A.; Crane, F.L. Synthesis of plastoquinone analogs and inhibition of photosynthetic and mammalian enzyme systems. Proc. Natl. Acad. Sci. USA 1972, 69, 3713–3717. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Lu, S. Plastoquinone and Ubiquinone in Plants: Biosynthesis, Physiological Function and Metabolic Engineering. Front. Plant. Sci. 2016, 7, 1898. [Google Scholar] [CrossRef] [Green Version]
- Bayrak, N.; Yildiz, M.; Yildirim, H.; Mataraci-Kara, E.; Tuyun, A.F. Novel plastoquinone analogs containing benzocaine and its analogs: Structure-based design, synthesis, and structural characterization. Res. Chem. Intermed. 2021, 47, 2125–2141. [Google Scholar] [CrossRef]
- Yildirim, H.; Bayrak, N.; Tuyun, A.F.; Kara, E.M.; Celik, B.O.; Gupta, G.K. 2,3-Disubstituted-1,4-naphthoquinones containing an arylamine with trifluoromethyl group: Synthesis, biological evaluation, and computational study. RSC Adv. 2017, 7, 25753–25764. [Google Scholar] [CrossRef] [Green Version]
- Boyd, M.R.; Pauli, K.D. Some Practical Considerations and Applications of the National-Cancer-Institute in-Vitro Anticancer Drug Discovery Screen. Drug Dev. Res. 1995, 34, 91–109. [Google Scholar] [CrossRef]
- Monks, A.; Scudiero, D.; Skehan, P.; Shoemaker, R.; Paull, K.; Vistica, D.; Hose, C.; Langley, J.; Cronise, P.; Vaigrowolff, A.; et al. Feasibility of a High-Flux Anticancer Drug Screen Using a Diverse Panel of Cultured Human Tumor-Cell Lines. JNCI J. Natl. Cancer Inst. 1991, 83, 757–766. [Google Scholar] [CrossRef]
- Erasmus, C.; Aucamp, J.; Smit, F.J.; Seldon, R.; Jordaan, A.; Warner, D.F.; N’Da, D.D. Synthesis and comparison of in vitro dual anti-infective activities of novel naphthoquinone hybrids and atovaquone. Bioorg. Chem. 2021, 114, 105118. [Google Scholar] [CrossRef]
- Refsgaard, H.H.F.; Jensen, B.F.; Brockhoff, P.B.; Padkjaer, S.B.; Guldbrandt, M.; Christensen, M.S. In silico prediction of membrane permeability from calculated molecular parameters. J. Med. Chem. 2005, 48, 805–811. [Google Scholar] [CrossRef]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [Green Version]
- Daina, A.; Michielin, O.; Zoete, V. iLOGP: A simple, robust, and efficient description of n-octanol/water partition coefficient for drug design using the GB/SA approach. J. Chem. Inf. Model. 2014, 54, 3284–3301. [Google Scholar] [CrossRef]
- Daina, A.; Zoete, V. A BOILED-Egg To Predict Gastrointestinal Absorption and Brain Penetration of Small Molecules. ChemMedChem 2016, 11, 1117–1121. [Google Scholar] [CrossRef] [Green Version]
- Meredith, A.M.; Dass, C.R. Increasing role of the cancer chemotherapeutic doxorubicin in cellular metabolism. J. Pharm. Pharmacol. 2016, 68, 729–741. [Google Scholar] [CrossRef] [Green Version]
- Guzman, C.; Bagga, M.; Kaur, A.; Westermarck, J.; Abankwa, D. ColonyArea: An ImageJ plugin to automatically quantify colony formation in clonogenic assays. PLoS ONE 2014, 9, e92444. [Google Scholar] [CrossRef]
- Abu, N.; Akhtar, M.N.; Ho, W.Y.; Yeap, S.K.; Alitheen, N.B. 3-Bromo-1-hydroxy-9,10-anthraquinone (BHAQ) inhibits growth and migration of the human breast cancer cell lines MCF-7 and MDA-MB231. Molecules 2013, 18, 10367–10377. [Google Scholar] [CrossRef] [Green Version]
- Perillo, B.; Di Donato, M.; Pezone, A.; Di Zazzo, E.; Giovannelli, P.; Galasso, G.; Castoria, G.; Migliaccio, A. ROS in cancer therapy: The bright side of the moon. Exp. Mol. Med. 2020, 52, 192–203. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Kim, H.S.; Seo, Y.R. Understanding of ROS-Inducing Strategy in Anticancer Therapy. Oxid. Med. Cell. Longev. 2019, 2019, 5381692. [Google Scholar] [CrossRef] [PubMed]
- Chang, H.-Y.; Huang, H.-C.; Huang, T.-C.; Yang, P.-C.; Wang, Y.-C.; Juan, H.-F. Flow Cytometric Detection of Reactive Oxygen Species. Bio-Protocol 2013, 3, e431. [Google Scholar] [CrossRef] [Green Version]
- Kong, N.; Zhang, H.; Feng, C.; Liu, C.; Xiao, Y.; Zhang, X.; Mei, L.; Kim, J.S.; Tao, W.; Ji, X. Arsenene-mediated multiple independently targeted reactive oxygen species burst for cancer therapy. Nat. Commun. 2021, 12, 4777. [Google Scholar] [CrossRef] [PubMed]
- Ge, Y.Y.; Kazi, A.; Marsilio, F.; Luo, Y.T.; Jain, S.; Brooks, W.; Daniel, K.G.; Guida, W.C.; Sebti, S.M.; Lawrence, H.R. Discovery and Synthesis of Hydronaphthoquinones as Novel Proteasome Inhibitors. J. Med. Chem. 2012, 55, 1978–1998. [Google Scholar] [CrossRef]
- Marastoni, M.; Trapella, C.; Scotti, A.; Fantinati, A.; Ferretti, V.; Marzola, E.; Eleonora, G.; Gavioli, R.; Preti, D. Naphthoquinone amino acid derivatives, synthesis and biological activity as proteasome inhibitors. J. Enzym. Inhib. Med. Chem. 2017, 32, 865–877. [Google Scholar] [CrossRef] [Green Version]
- Uysal, S.; Soyer, Z.; Saylam, M.; Tarikogullari, A.H.; Yilmaz, S.; Kirmizibayrak, P.B. Design, synthesis and biological evaluation of novel naphthoquinone-4-aminobenzensulfonamide/carboxamide derivatives as proteasome inhibitors. Eur. J. Med. Chem. 2021, 209, 112890. [Google Scholar] [CrossRef]
- Fogli, S.; Galimberti, S.; Gori, V.; Del Re, M.; Danesi, R. Pharmacology differences among proteasome inhibitors: Implications for their use in clinical practice. Pharmacol. Res. 2021, 167, 105537. [Google Scholar] [CrossRef]
- Lubos, M.; Dębowski, D.; Barcińska, E.; Meid, A.; Inkielewicz-Stepniak, I.; Burster, T.; Rolka, K. Inhibition of human constitutive 20S proteasome and 20S immunoproteasome with novel N-terminally modified peptide aldehydes and their antitumor activity. Pept. Sci. 2019, 111, e24100. [Google Scholar] [CrossRef]
- Grever, M.R.; Schepartz, S.A.; Chabner, B.A. The National-Cancer-Institute—Cancer Drug Discovery and Development Program. Semin. Oncol. 1992, 19, 622–638. [Google Scholar]
- Skehan, P.; Storeng, R.; Scudiero, D.; Monks, A.; McMahon, J.; Vistica, D.; Warren, J.T.; Bokesch, H.; Kenney, S.; Boyd, M.R. New colorimetric cytotoxicity assay for anticancer-drug screening. J. Natl. Cancer Inst. 1990, 82, 1107–1112. [Google Scholar] [CrossRef]
- Morris, G.M.; Goodsell, D.S.; Halliday, R.S.; Huey, R.; Hart, W.E.; Belew, R.K.; Olson, A.J. Automated docking using a Lamarckian genetic algorithm and an empirical binding free energy function. J. Comput. Chem. 1998, 19, 1639–1662. [Google Scholar] [CrossRef] [Green Version]
- Harshbarger, W.; Miller, C.; Diedrich, C.; Sacchettini, J. Crystal Structure of the Human 20S Proteasome in Complex with Carfilzomib. Structure 2015, 23, 418–424. [Google Scholar] [CrossRef] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [Green Version]
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple Ligand–Protein Interaction Diagrams for Drug Discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecule | BrPQ5 (NCI: D-825197/1) | ||
|---|---|---|---|
| Panel/Cell Line | GI50 | TGI | LC50 |
| Leukemia | |||
| CCRF-CEM | 2.33 | >100 | >100 |
| HL-60 (TB) | 2.34 | 6.82 | >100 |
| K-562 | 2.25 | >100 | >100 |
| MOLT-4 | 2.67 | >100 | >100 |
| RPMI-8226 | 2.21 | 5.01 | >100 |
| SR | 3.21 | >100 | >100 |
| Non-Small Cell Lung Cancer | |||
| A549/ATCC | 18.70 | 45.70 | >100 |
| EKVX | 1.69 | 3.12 | 5.77 |
| HOP-62 | 15.00 | 38.60 | 99.70 |
| HOP-92 | 1.55 | 3.08 | 6.10 |
| NCI-H226 | 13.00 | 27.60 | 58.50 |
| NCI-H23 | 4.41 | 23.70 | >100 |
| NCI-H322M | 14.60 | 27.80 | 53.10 |
| NCI-H460 | 18.80 | 42.60 | 96.10 |
| NCI-H522 | 12.70 | 31.80 | 79.50 |
| Colon Cancer | |||
| COLO 205 | 21.30 | 51.10 | >100 |
| HCC-2998 | 12.40 | 26.50 | 56.50 |
| HCT-116 | 1.76 | 3.37 | 6.46 |
| HCT-15 | 11.40 | 27.70 | 67.40 |
| HT29 | 27.40 | 79.60 | >100 |
| KM12 | 5.02 | 19.00 | 58.00 |
| SW-620 | 2.07 | 4.54 | 9.97 |
| CNS Cancer | |||
| SF-268 | 11.90 | 28.20 | 67.00 |
| SF-295 | 13.80 | 27.00 | 52.80 |
| SF-539 | 13.50 | 26.70 | 52.90 |
| SNB-19 | 10.50 | 22.80 | 49.10 |
| SNB-75 | 11.00 | 23.50 | 50.20 |
| U251 | 19.30 | 3.98 | 8.24 |
| Melanoma | |||
| LOX IMVI | 1.67 | 3.17 | 5.99 |
| MALME-3M | 14.50 | 28.30 | 55.20 |
| M14 | 11.70 | 24.80 | 52.60 |
| MDA-MB-435 | 1.91 | 3.49 | 6.36 |
| SK-MEL-2 | 11.40 | 28.30 | 70.60 |
| SK-MEL-28 | 2.30 | 4.76 | 9.88 |
| SK-MEL-5 | 13.10 | 26.00 | 51.50 |
| UACC-257 | 1.75 | 3.37 | 6.51 |
| UACC-62 | 7.74 | 20.70 | 46.80 |
| Ovarian Cancer | |||
| IGROV1 | 1.80 | 3.39 | 6.38 |
| OVCAR-3 | 1.72 | 3.13 | 5.72 |
| OVCAR-4 | 1.60 | 2.97 | 5.49 |
| OVCAR-5 | 5.07 | 21.80 | 75.40 |
| OVCAR-8 | 2.22 | 5.31 | >100 |
| NCI/ADR-RES | 2.04 | 4.65 | >100 |
| SK-OV-3 | 12.30 | 26.10 | 55.00 |
| Renal Cancer | |||
| 786-0 | 15.70 | 29.90 | 56.90 |
| A498 | 1.62 | 33.60 | >100 |
| ACHN | 2.87 | 8.76 | 29.40 |
| CAKI-1 | 1.72 | 3.11 | 5.63 |
| RXF 393 | 2.71 | 8.13 | 32.30 |
| SN12C | 11.00 | 23.20 | 49.10 |
| TK-10 | 25.20 | 40.90 | 66.30 |
| UO-31 | 1.73 | 3.16 | 5.77 |
| Prostate Cancer | |||
| PC-3 | 2.78 | 9.05 | 33.50 |
| DU-145 | 15.00 | 29.10 | 56.50 |
| Breast Cancer | |||
| MCF7 | 1.78 | 3.77 | 7.98 |
| MDA-MB-231/ATCC | 1.88 | 3.96 | 8.34 |
| HS 578T | 11.80 | 57.50 | >100 |
| BT-549 | 11.80 | 24.40 | 50.40 |
| T-47D | 1.93 | 4.81 | >100 |
| MDA-MB-468 | 1.56 | 3.16 | 6.40 |
| Cell Lines | IC50± (µM) | |
|---|---|---|
| BrPQ5 | DOXO | |
| U-251 | >100 | >100 |
| MCF-7 | 33.57 ± 1.7 | 17.52 ± 2.6 |
| MDA-MB-231 | 33.65 ± 2.2 | 44.66 ± 9.8 |
| DU-145 | 83.89 ± 12.8 | >100 |
| HCT-116 | 74.33 ± 11 | 12.84 ± 4.5 |
| VHF93 | >100 | >100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jannuzzi, A.T.; Yilmaz Goler, A.M.; Bayrak, N.; Yıldız, M.; Yıldırım, H.; Karademir Yilmaz, B.; Shilkar, D.; Venkatesan, R.J.; Jayaprakash, V.; TuYuN, A.F. Exploring the Anticancer Effects of Brominated Plastoquinone Analogs with Promising Cytotoxic Activity in MCF-7 Breast Cancer Cells via Cell Cycle Arrest and Oxidative Stress Induction. Pharmaceuticals 2022, 15, 777. https://doi.org/10.3390/ph15070777
Jannuzzi AT, Yilmaz Goler AM, Bayrak N, Yıldız M, Yıldırım H, Karademir Yilmaz B, Shilkar D, Venkatesan RJ, Jayaprakash V, TuYuN AF. Exploring the Anticancer Effects of Brominated Plastoquinone Analogs with Promising Cytotoxic Activity in MCF-7 Breast Cancer Cells via Cell Cycle Arrest and Oxidative Stress Induction. Pharmaceuticals. 2022; 15(7):777. https://doi.org/10.3390/ph15070777
Chicago/Turabian StyleJannuzzi, Ayse Tarbin, Ayse Mine Yilmaz Goler, Nilüfer Bayrak, Mahmut Yıldız, Hatice Yıldırım, Betul Karademir Yilmaz, Deepak Shilkar, Raghusrinivasan Jayaprakash Venkatesan, Venkatesan Jayaprakash, and Amaç Fatih TuYuN. 2022. "Exploring the Anticancer Effects of Brominated Plastoquinone Analogs with Promising Cytotoxic Activity in MCF-7 Breast Cancer Cells via Cell Cycle Arrest and Oxidative Stress Induction" Pharmaceuticals 15, no. 7: 777. https://doi.org/10.3390/ph15070777
APA StyleJannuzzi, A. T., Yilmaz Goler, A. M., Bayrak, N., Yıldız, M., Yıldırım, H., Karademir Yilmaz, B., Shilkar, D., Venkatesan, R. J., Jayaprakash, V., & TuYuN, A. F. (2022). Exploring the Anticancer Effects of Brominated Plastoquinone Analogs with Promising Cytotoxic Activity in MCF-7 Breast Cancer Cells via Cell Cycle Arrest and Oxidative Stress Induction. Pharmaceuticals, 15(7), 777. https://doi.org/10.3390/ph15070777