
Neuroprotective Effect of Morin Hydrate against Attention-Deficit/Hyperactivity Disorder (ADHD) Induced by MSG and/or Protein Malnutrition in Rat Pups: Effect on Oxidative/Monoamines/Inflammatory Balance and Apoptosis

 ,
,  ,
,  , , ,
, , ,
Abstract
:1. Introduction
2. Results
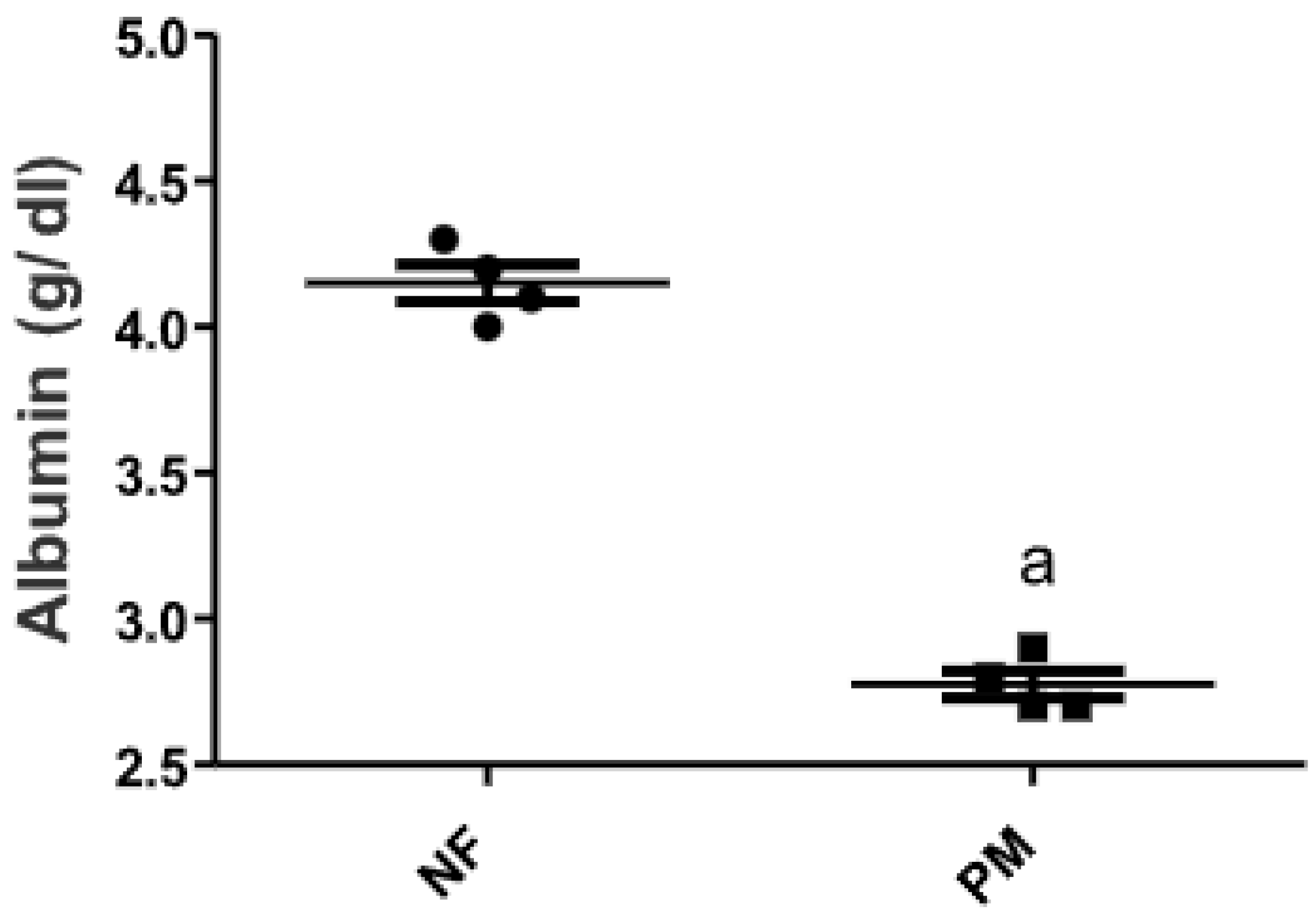
2.1. The Level of Serum Albumin in PM and/or MSG—Fed Rat Pups
2.2. Rat Behavior Tests
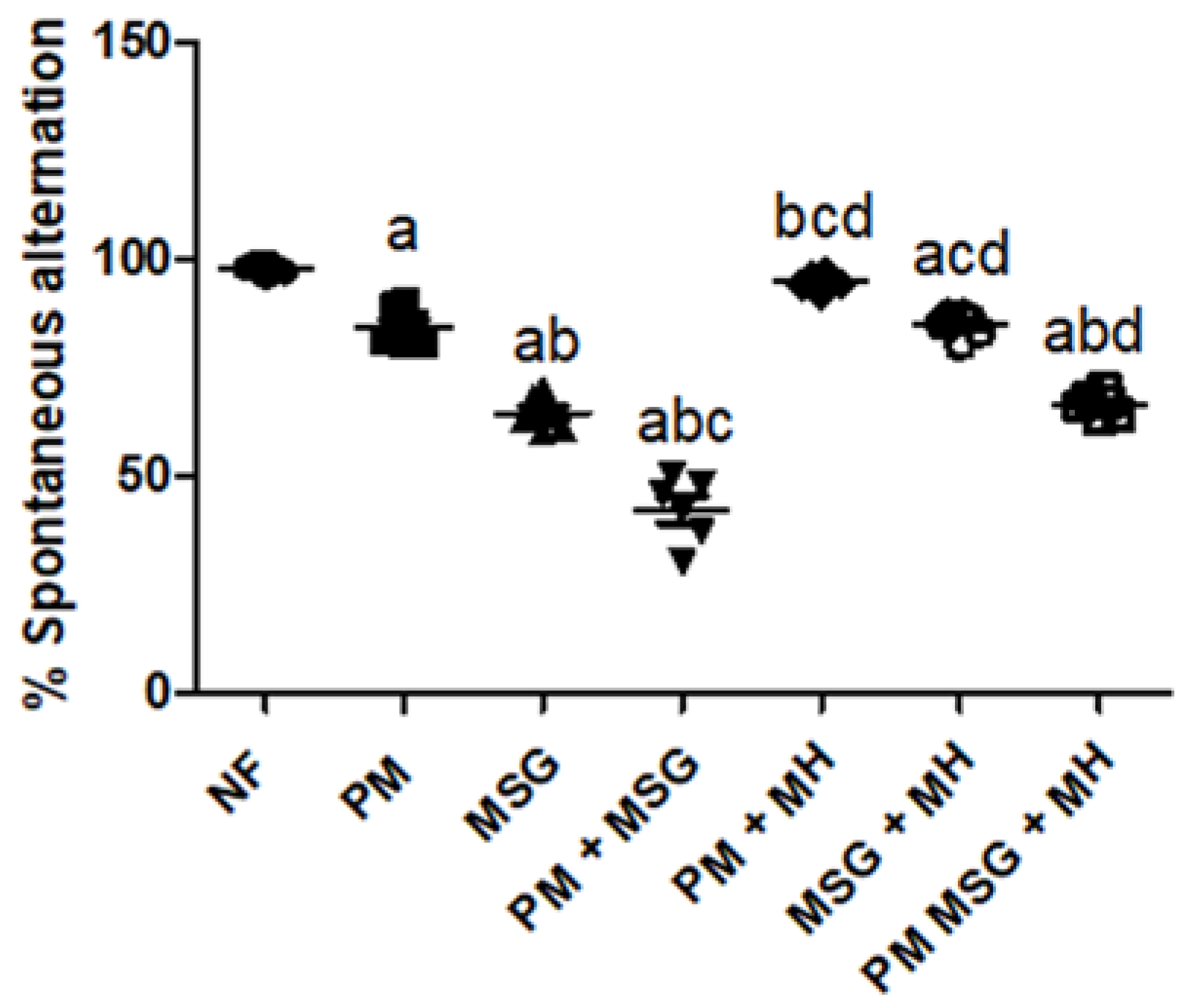
2.2.1. MH Attenuates Alterations in Spatial Working Memory of PM and/or MSG—Fed Rat Pups
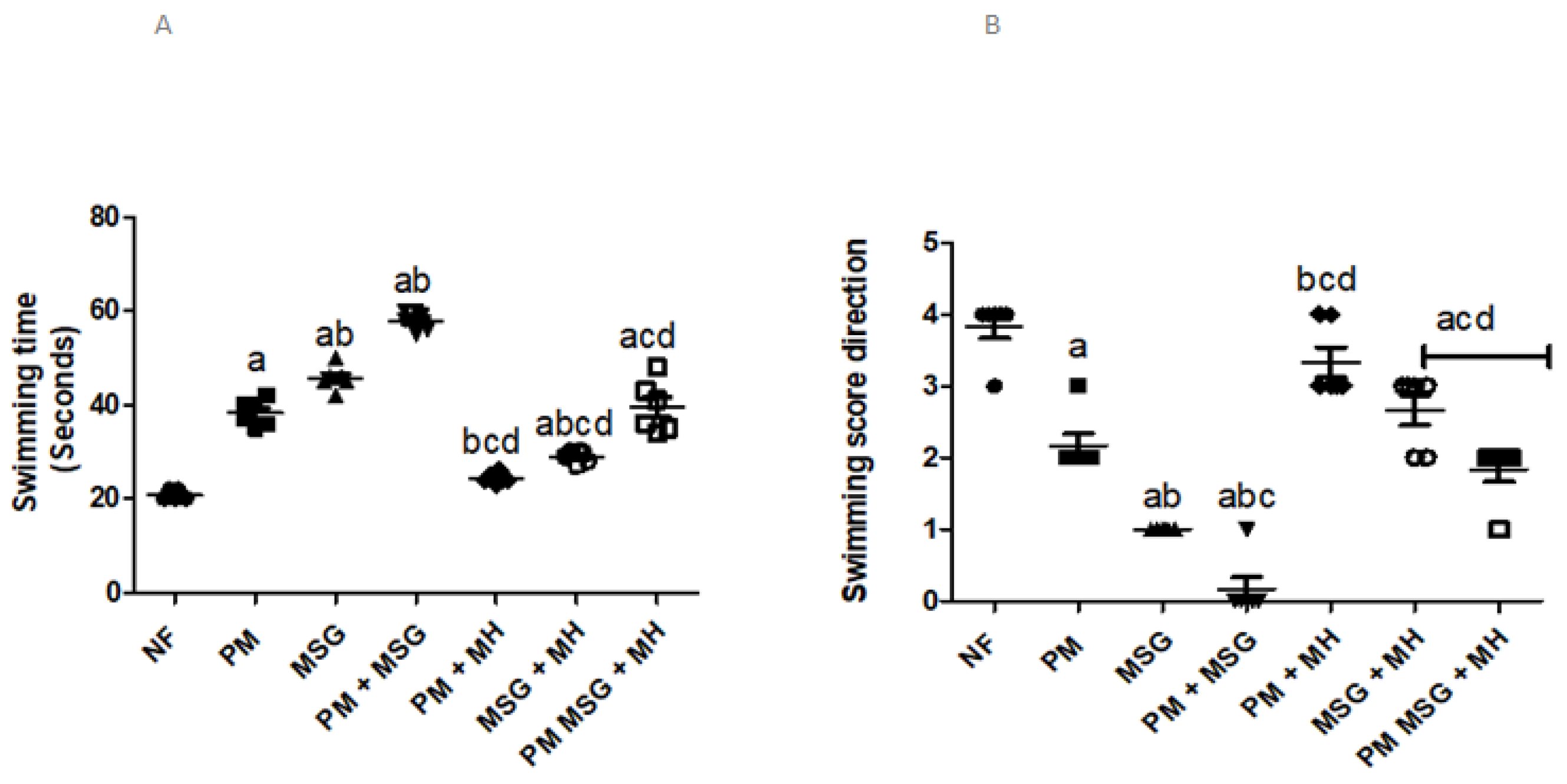
2.2.2. MH Affects Swimming Ability in PM and/or MSG—Fed Rat Pups
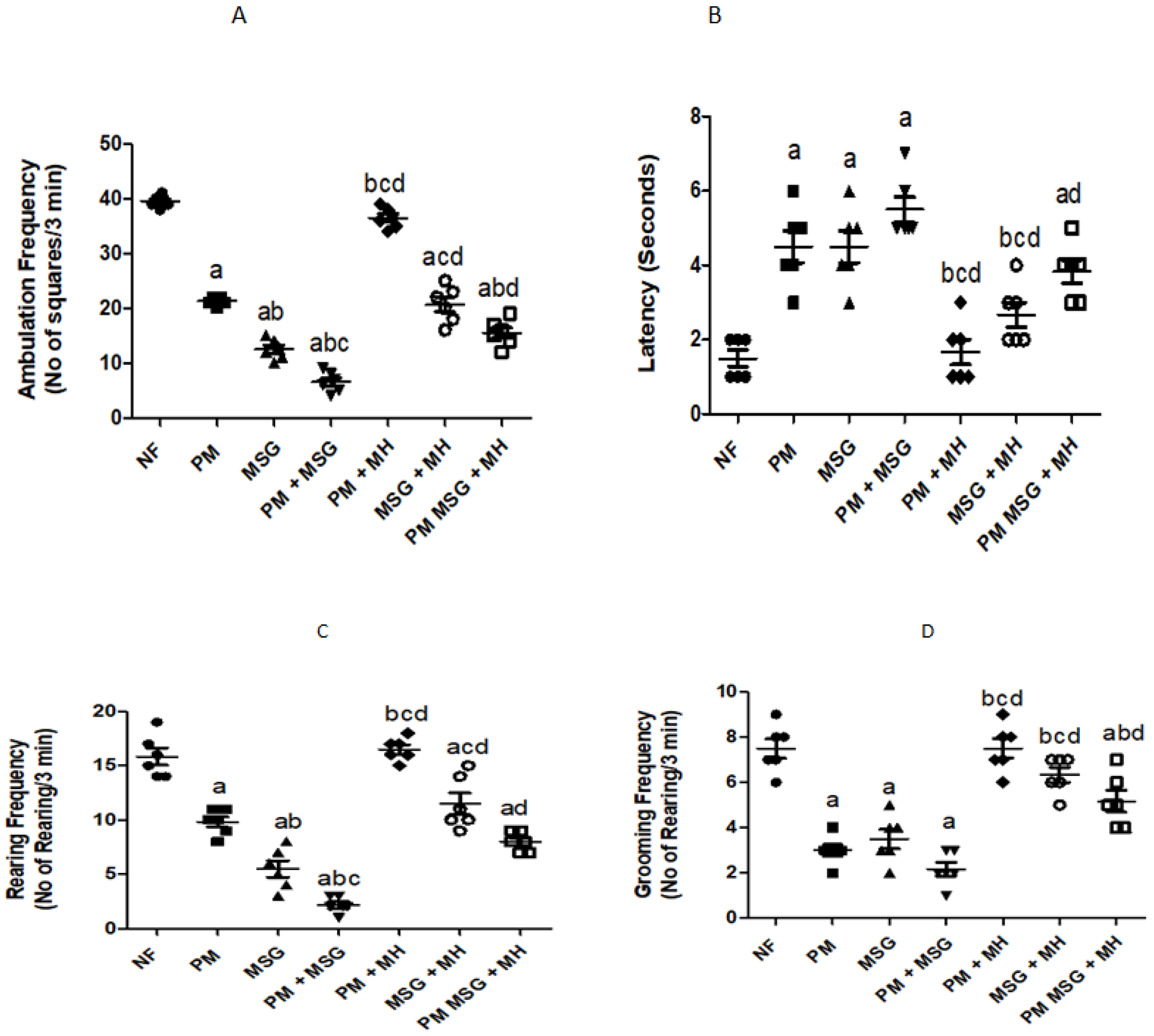
2.2.3. MH Restores Frequencies in OFT of PM and/or MSG—Fed Rat Pups
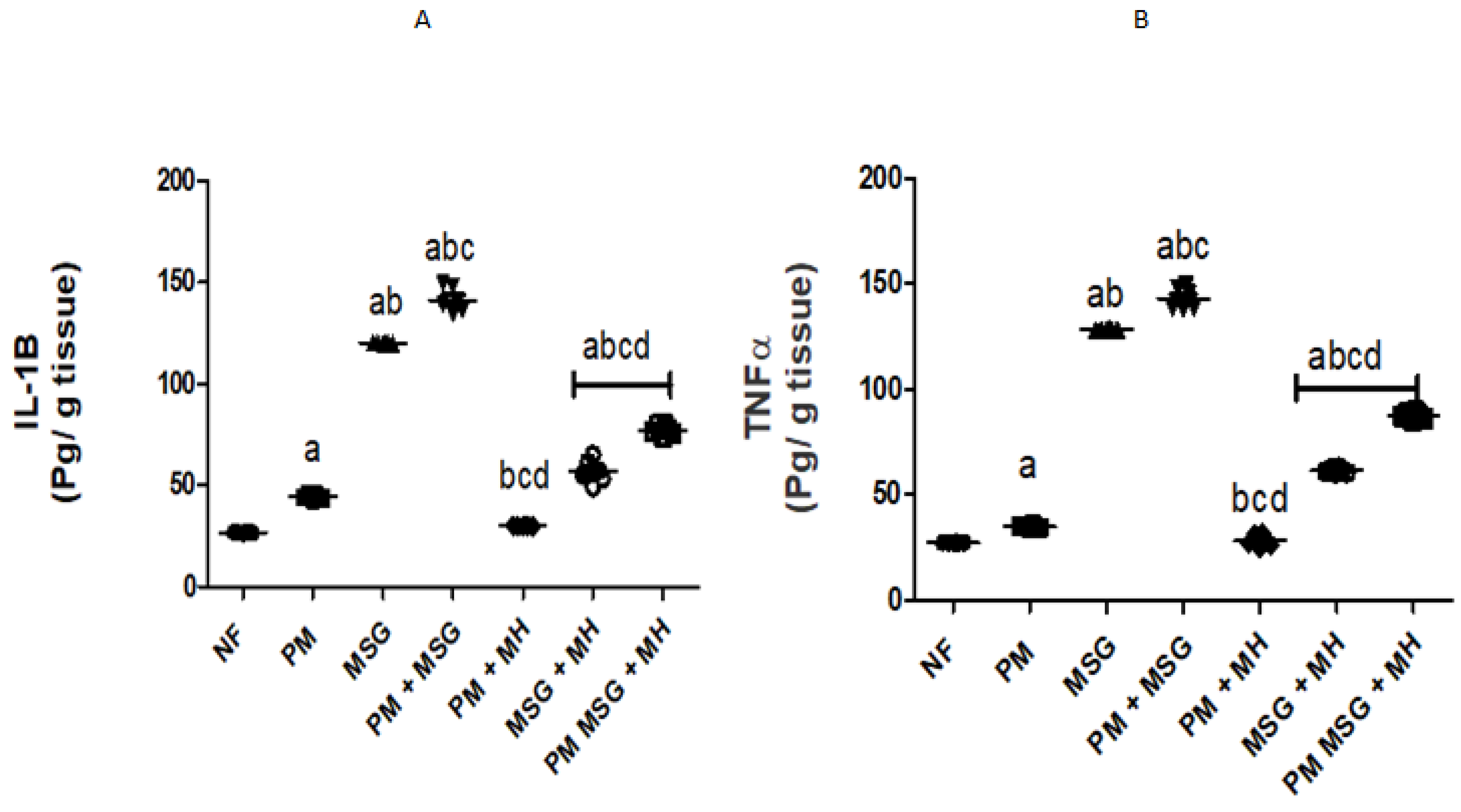
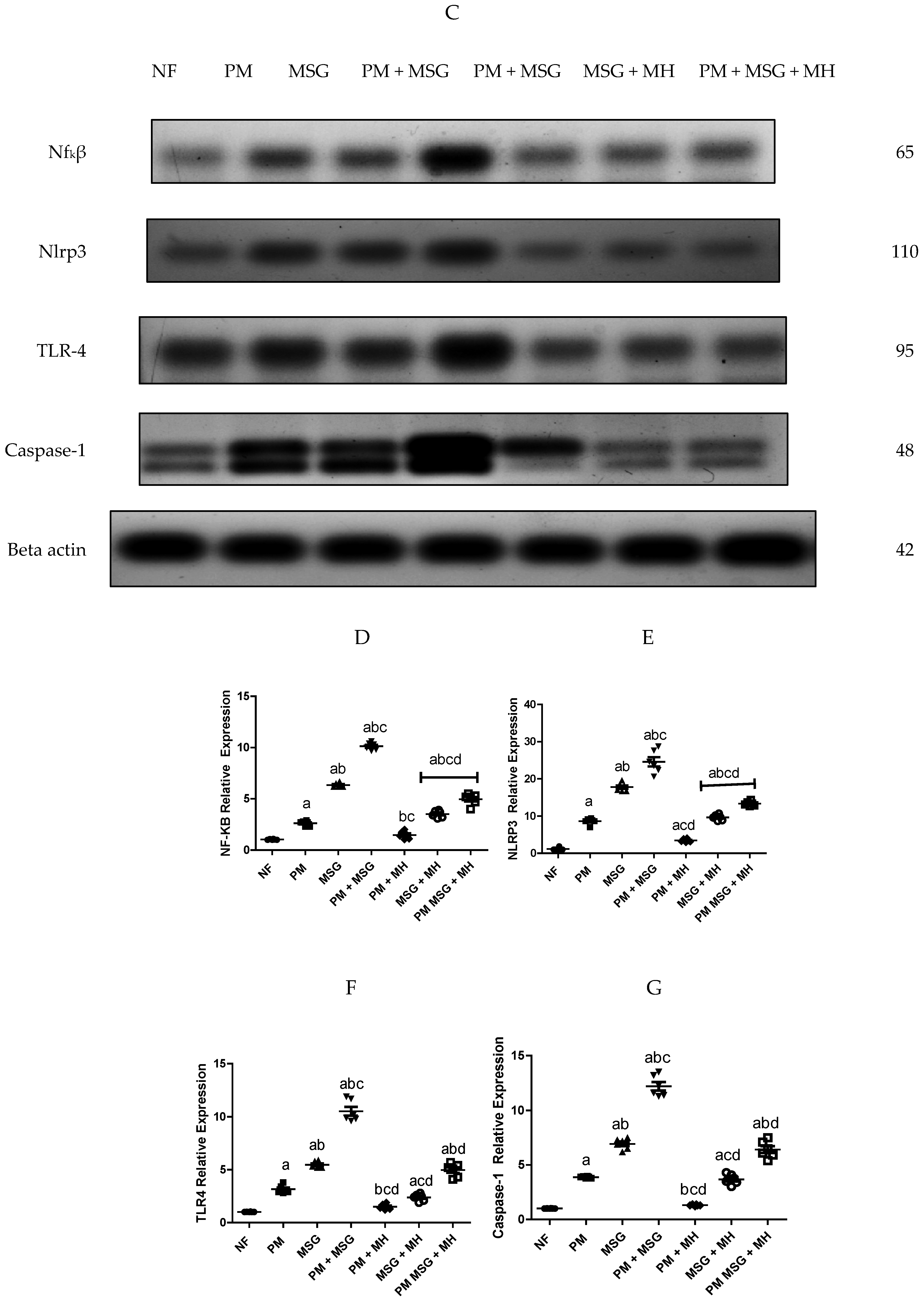
2.3. MH Reestablishes Inflammatory Biomarkers in PM and/or MSG—Fed Rat Pups
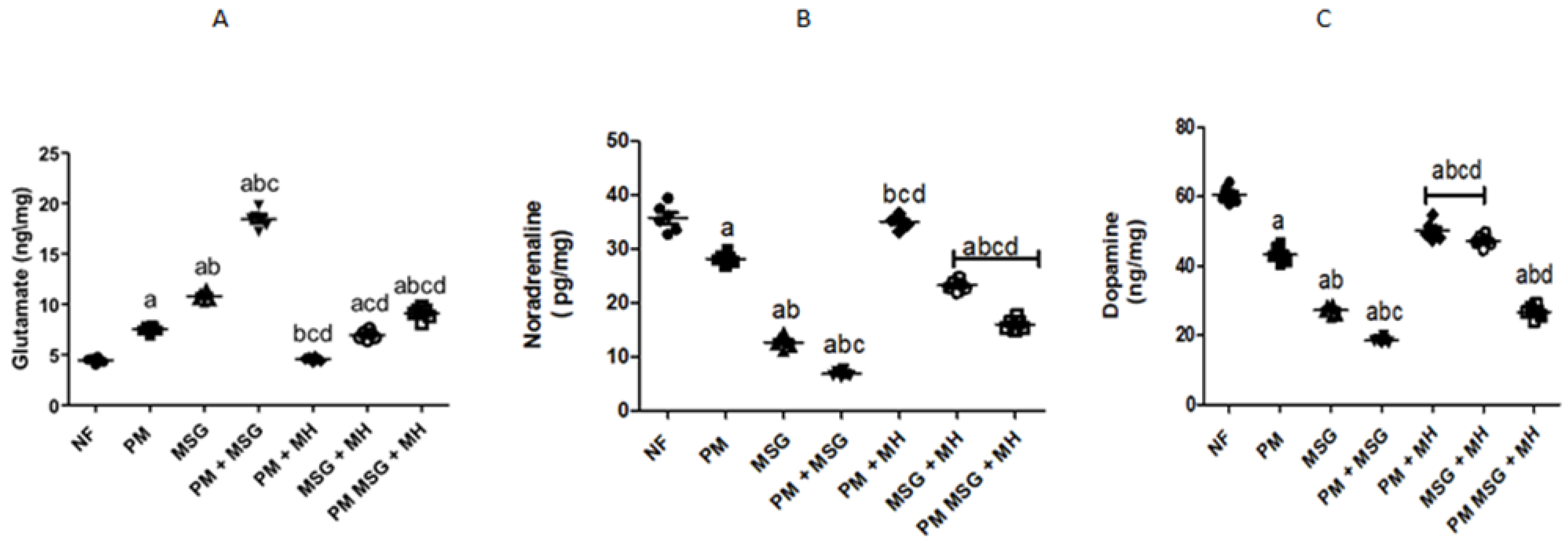
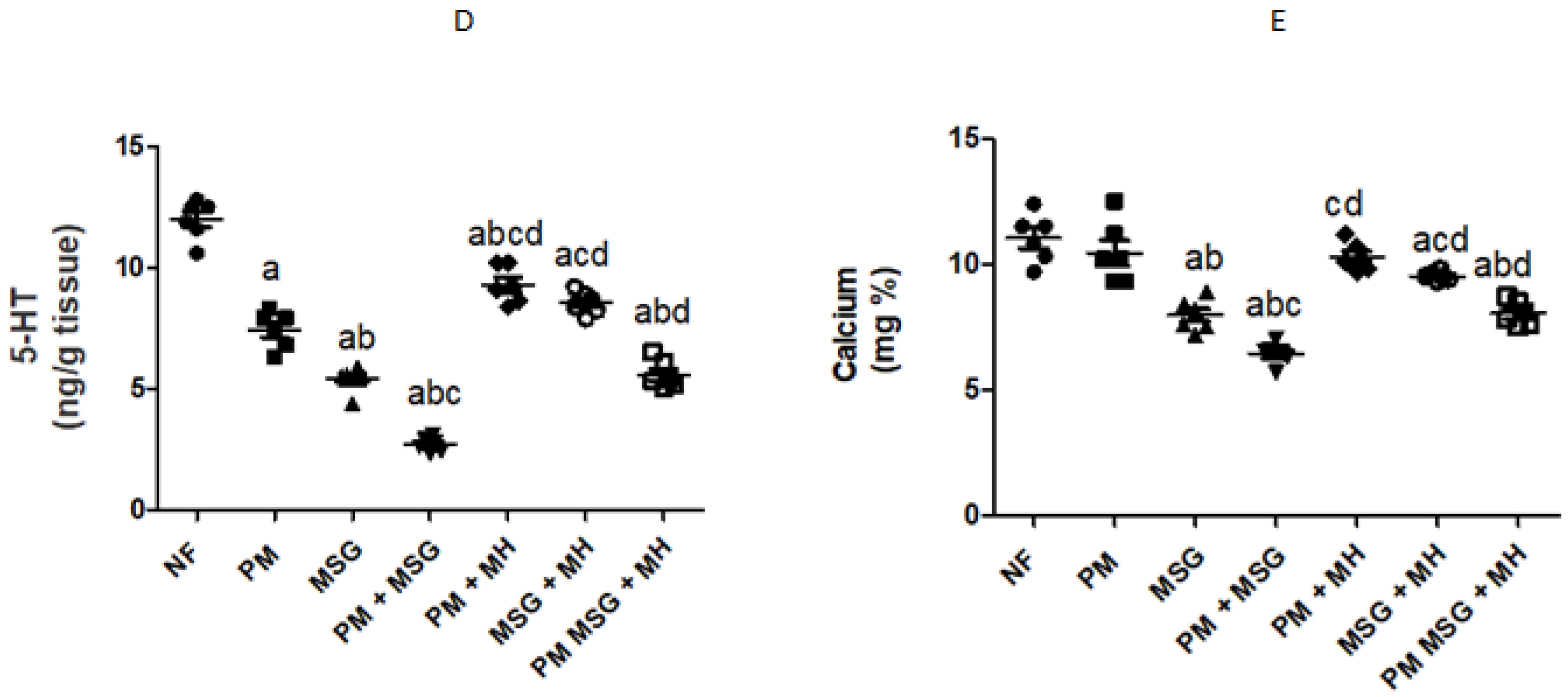
2.4. MH Balances Monoamines Neurotransmittors of PM and/or MSG—Fed Rat Pups
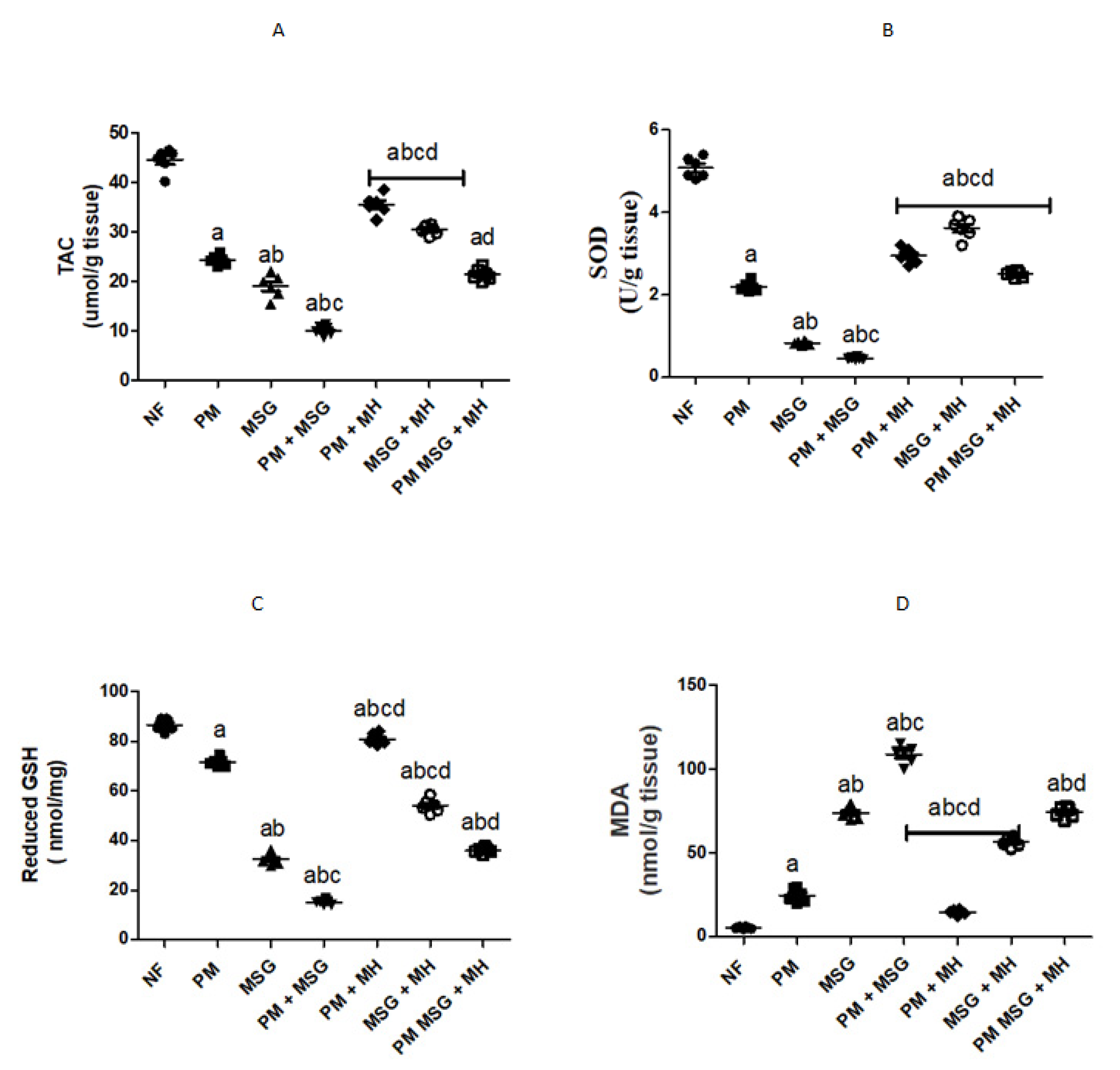
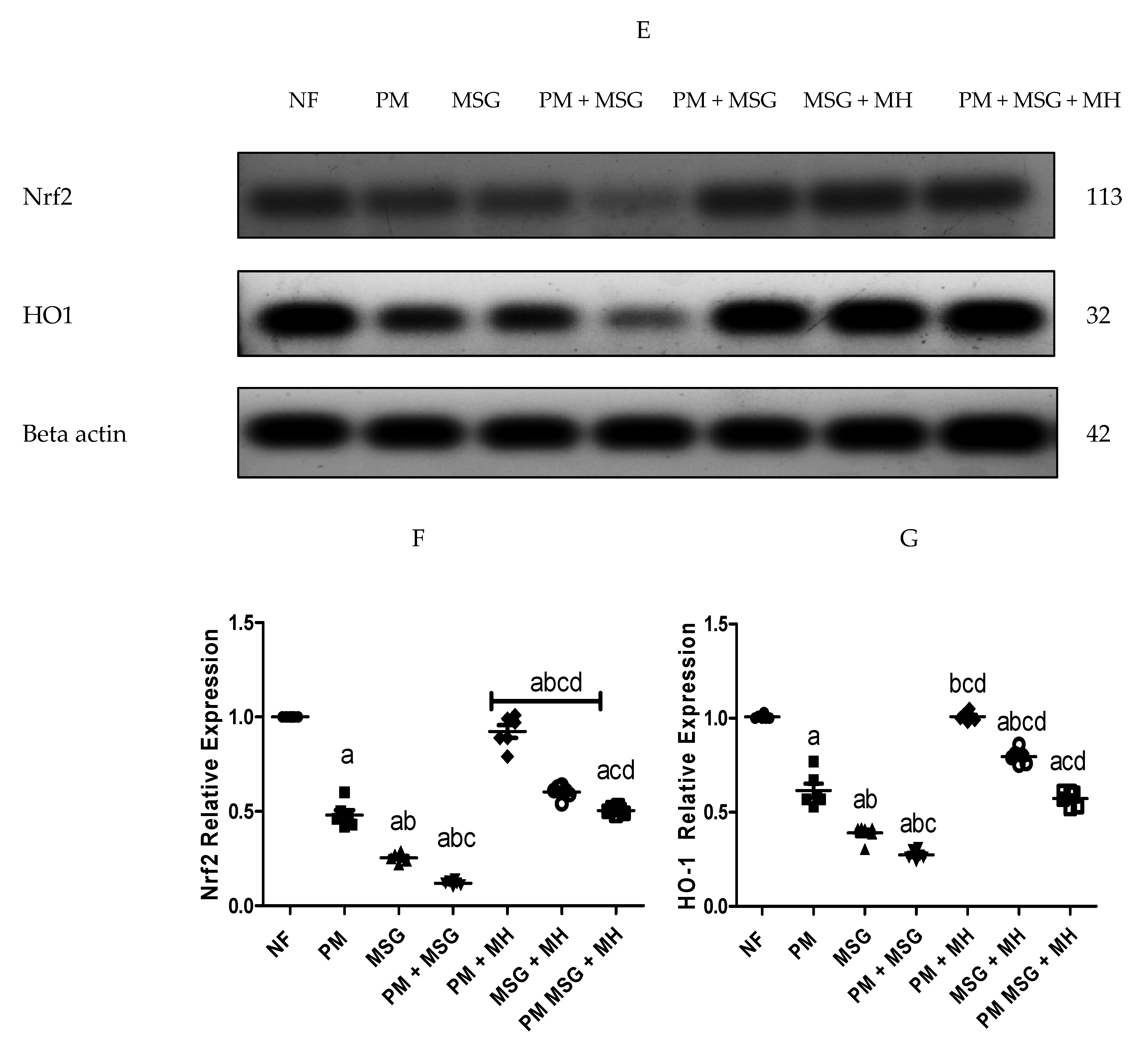
2.5. MH Modify Brain Oxidative Stress Biomarkers of PM and/or MSG—Fed Rat Pups
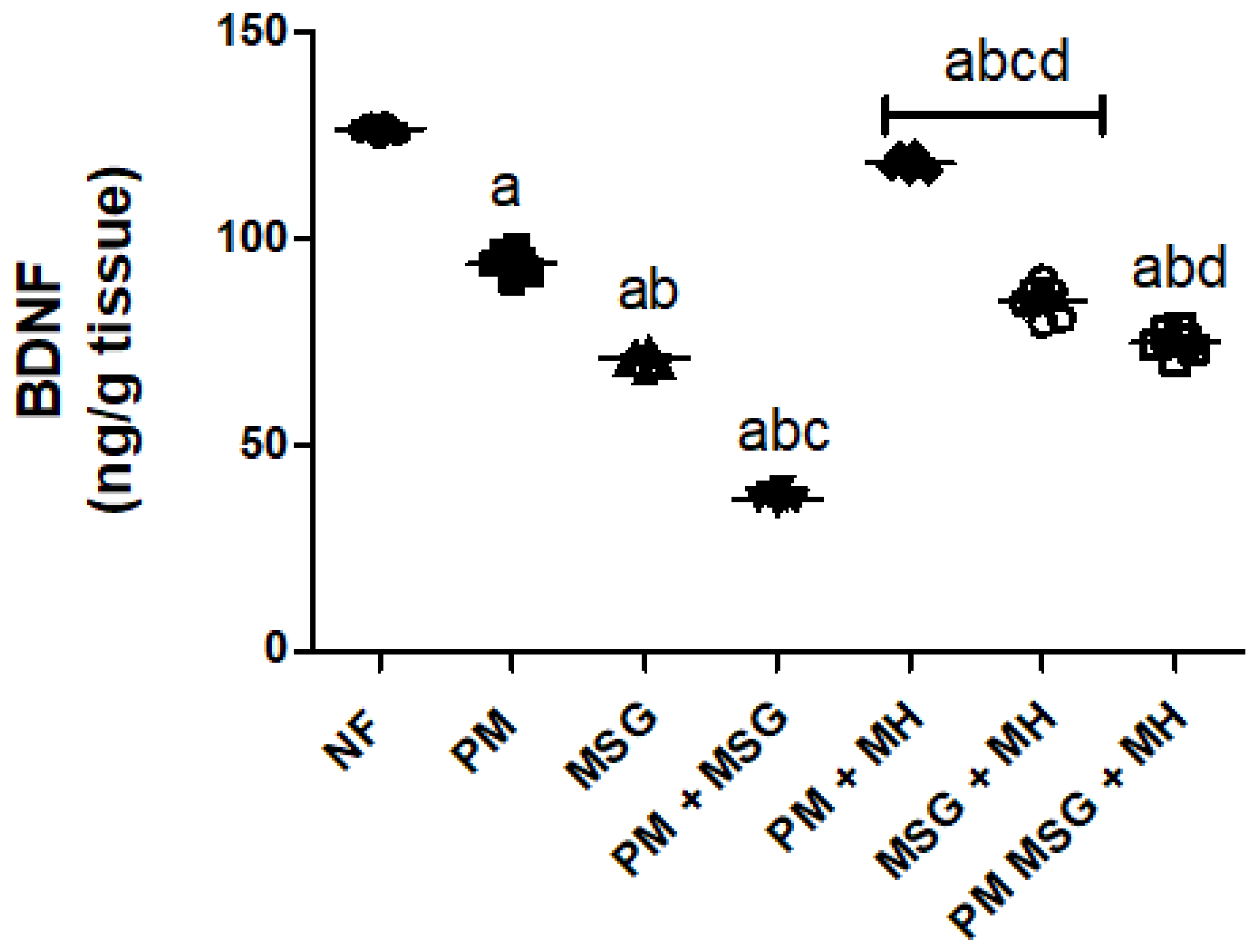
2.6. MH Adjust BDNF Content of PM and/or MSG—Fed Rat Pups
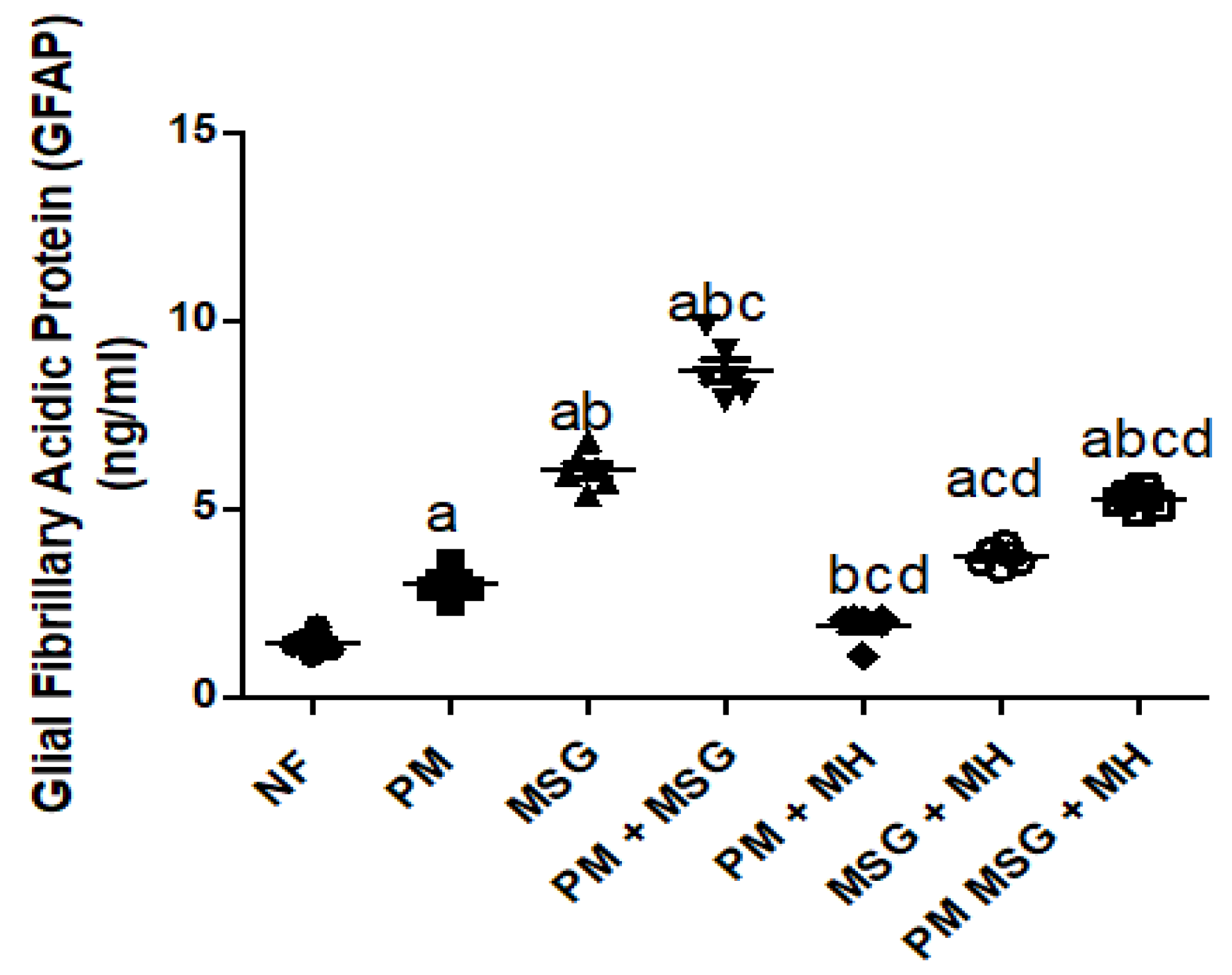
2.7. MH Correct GFAP Brain Content of PM and/or MSG—Fed Rat Pups
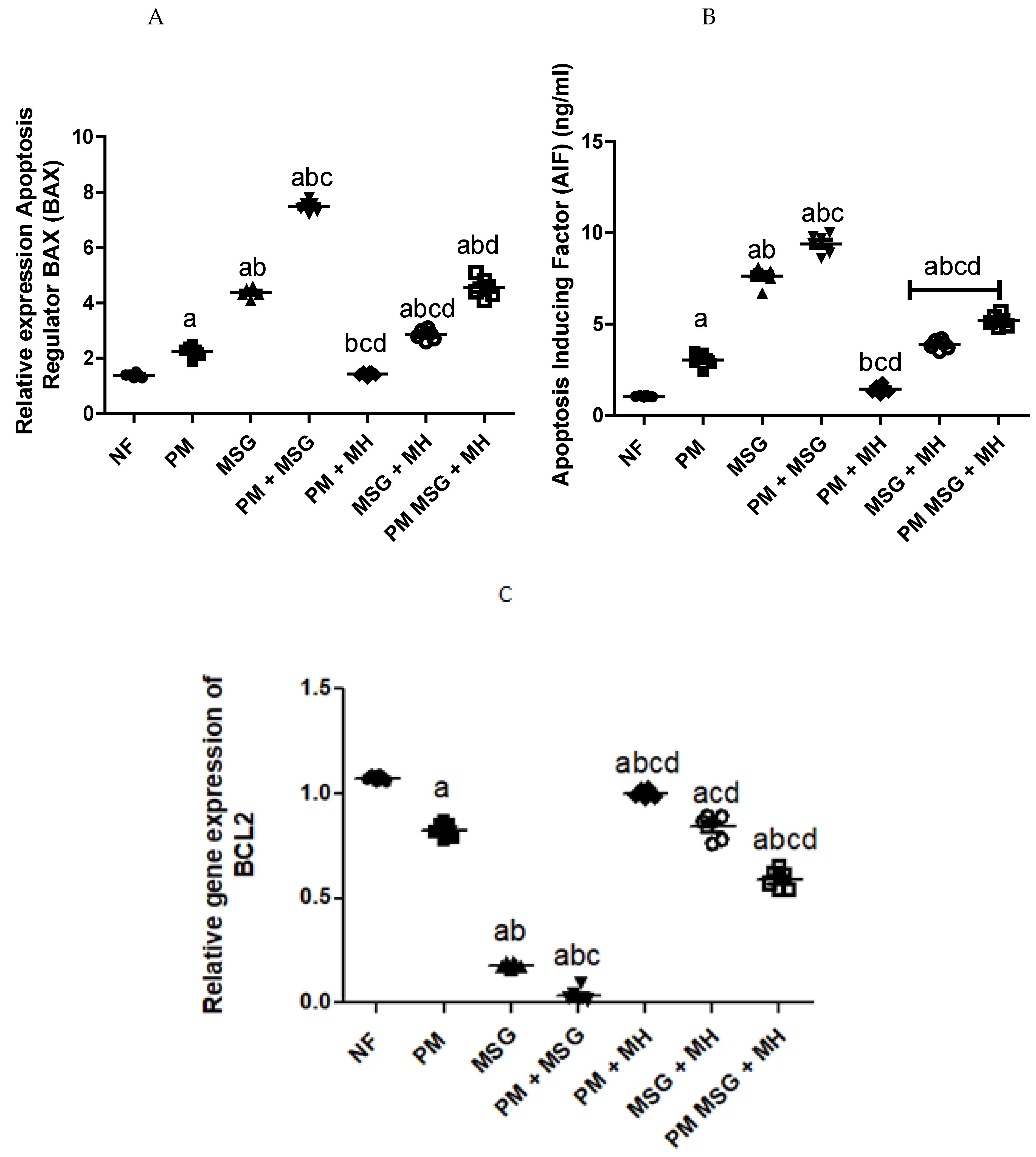
2.8. MH Reverts Apoptotic/Anti-Apoptotic Markers of PM and/or MSG—Fed Rat Pups
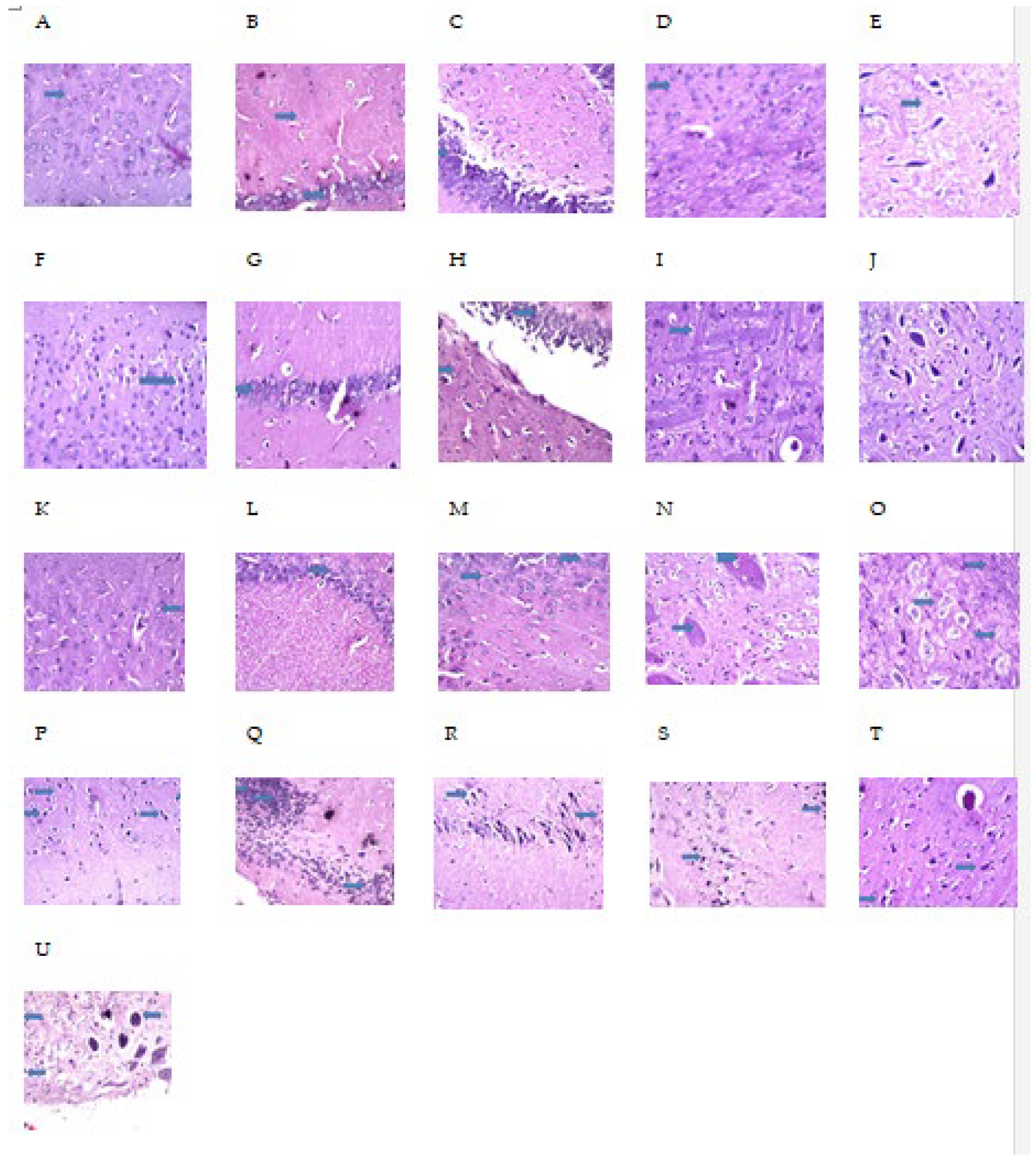
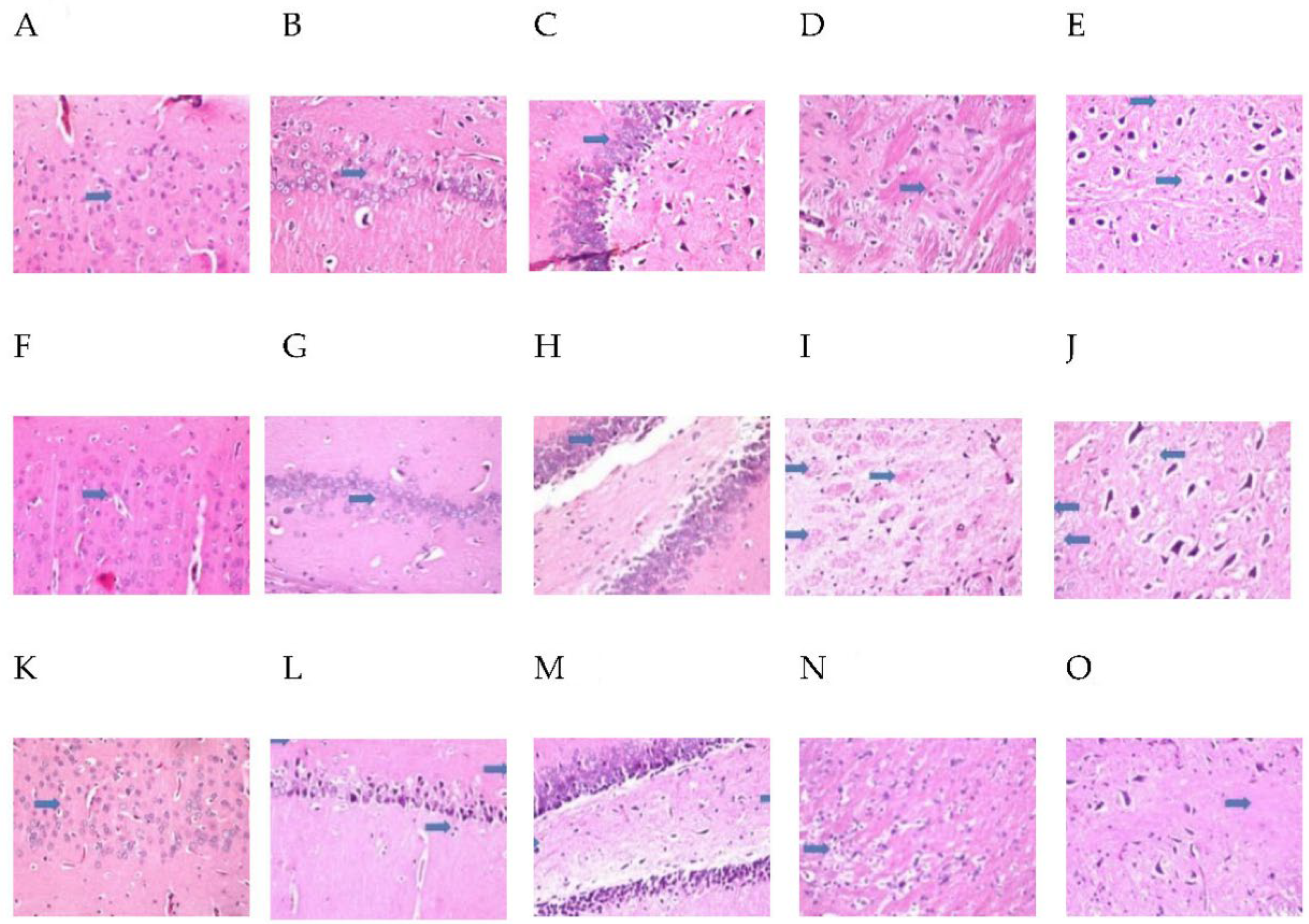
2.9. MH Alter Histopathological Brain Specimen of PM and/or MSG—Fed Rat Pups
3. Discussion
4. Materials and Methods
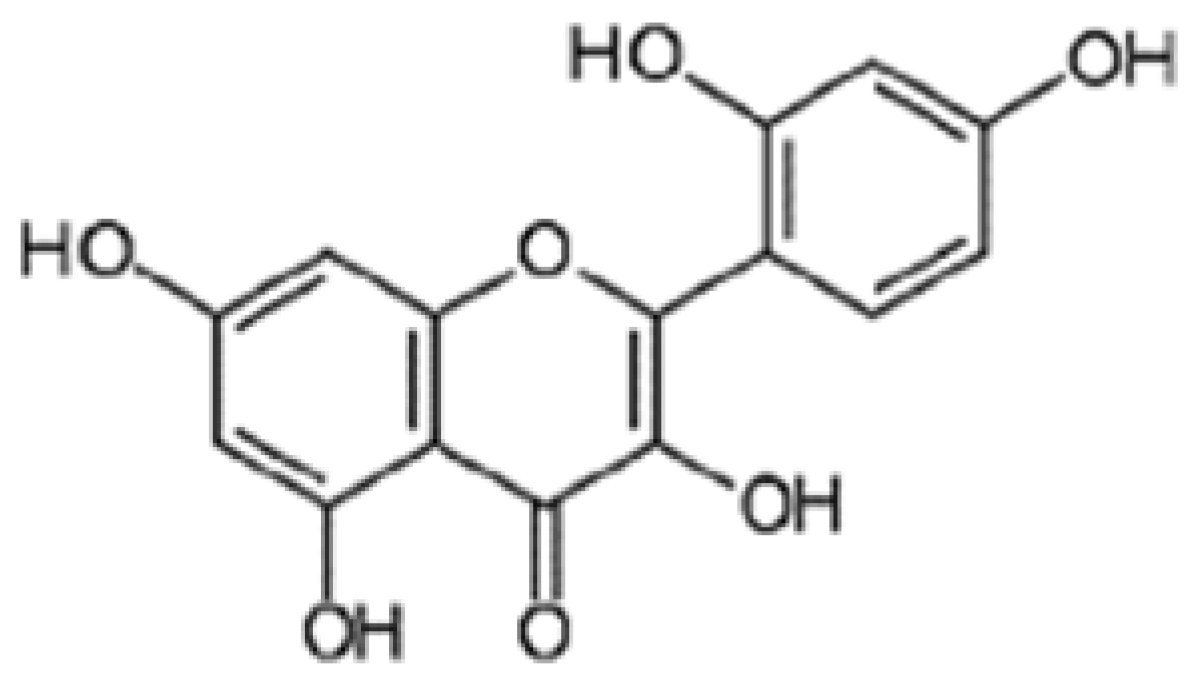
4.1. Drugs and Chemicals
4.2. Animals
4.3. Diet
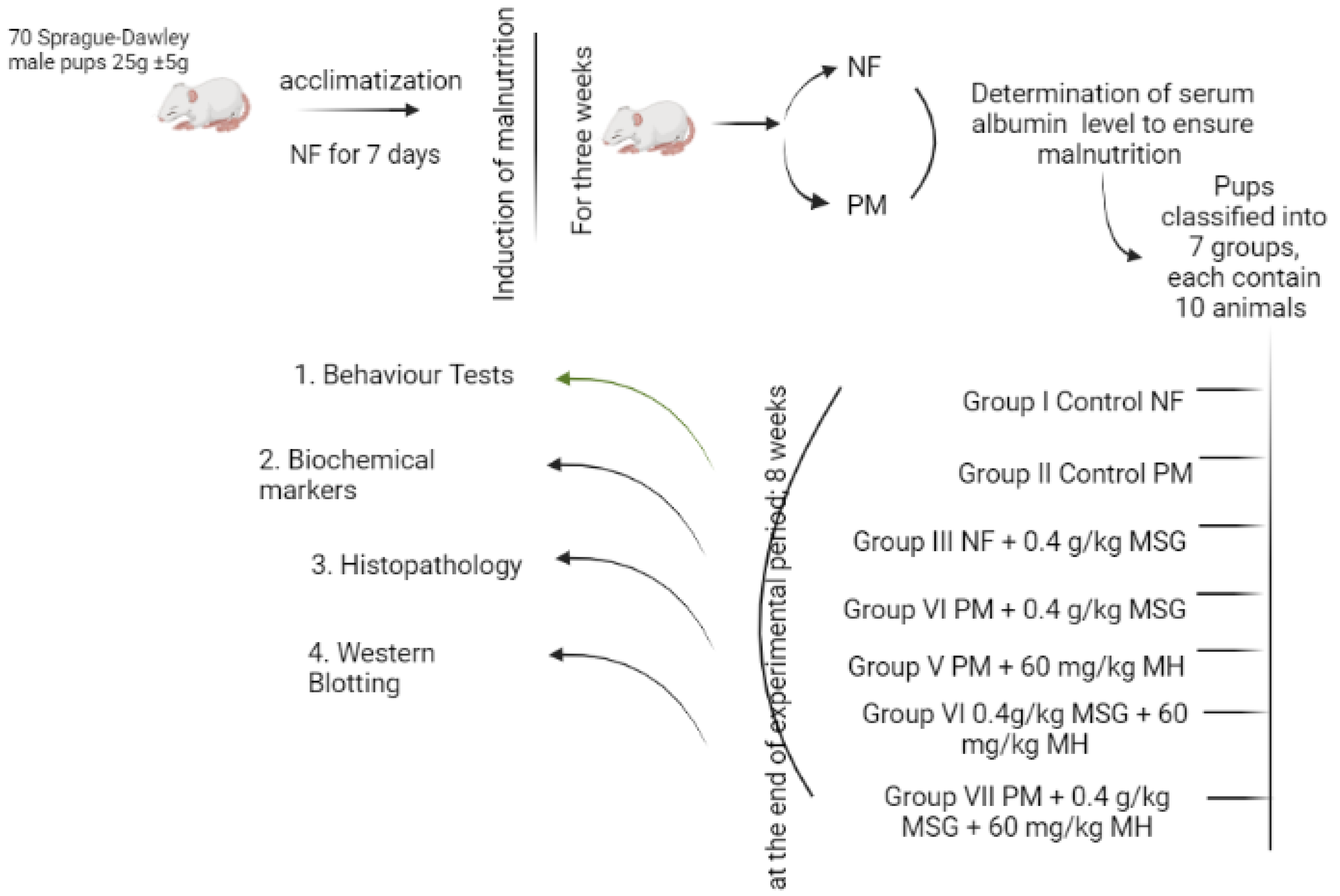
4.4. Experimental Design
4.5. Determination of Serum Albumin Content
4.6. Behavior Assessment
4.6.1. Y-Maze Task (for Assessment of Spatial Working Memory)
4.6.2. Swimming Test (for Assessment of Motor Coordination) Reflecting the Cognitive Function)
4.6.3. Open-Field Test (OFT)
4.7. Enzyme-Linked Immunosorbent Assays (ELISA)
4.8. Colorimetrical Analysis
4.9. Western Blotting Analysis
4.10. Gene Expression Measurement of Bcl-2 and Bax in Brain Tissue by Quantitative Real-Time PCR (qPCR) Analysis
4.11. Histopathological Analysis
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| PM | Low protein diet |
| NF | Normal protein diet |
| ADHD | Attention deficit hyperactivity disorder |
| MSG | Monosodium glutamate |
| Nrf2 | nuclear factor erythroid-2 related factor |
| HO-1 | heme oxygenase-1 |
| ROS | Reactive oxygen species |
| MH | Morin hydrate |
| ELISA | Enzyme-Linked Immunosorbent Assays |
| GFAP | Glial fibrillary acidic protein |
| AIF | Apoptosis inducing factor |
| H&E | Hematoxylin and eosin |
| OFT | Open field test |
| 5-HT | Serotonin |
| TNF-α | Tumor necrosis factor-α |
| IL-1β | Interleukin-1 β |
| NFκB | nuclear factor kapa B |
| TLR-4 | Toll-like receptor-4 |
| GSH | Reduced glutathione |
| TAC | Total antioxidant count |
| Bcl-2 | B-cell lymphoma-2 |
| Bax | Bcl-2 associated X |
| BDNF | Brain-derived neurotrophic factor |
| NA | Noradrenaline |
| MDA | Malondialdehyde |
| Nlrp-3 | NOD-like receptor protein-3 |
References
- Chertoff, M. Protein Malnutrition and Brain Development. Brain Disord. Ther. 2015, 4, 1–6. [Google Scholar] [CrossRef]
- Hazzaa, S.M.; Abdelaziz, S.A.M.; Eldaim, M.A.A.; AbdelDaim, M.M.; Elgarawany, G.E. Neuroprotective potential of Allium sativum against monosodium glutamate-induced excitotoxicity: Impact on short-term memory, gliosis and oxidative stress. Nutrients 2020, 12, 1028. [Google Scholar] [CrossRef] [PubMed]
- Kazmi, Z.; Fatima, I.; Perveen, S.; Malik, S.S. Monosodium glutamate: Review on clinical reports. Int. J. Food Prop. 2017, 20, 1807–1815. [Google Scholar] [CrossRef]
- GonzaÂlez-Burgosa, I.; PeÂrez-Vegaa, M.I.; Beas-ZaÂrate, C. Neonatal exposure to monosodium glutamate induces cell death and dendritic hypotrophy in rat prefrontocortical pyramidal neurons. Neurosci. Lett. 2001, 297, 69–72. [Google Scholar] [CrossRef]
- Brandes, M.S.; Gray, N.E. NRF2 as a therapeutic target in neurodegenerative diseases. ASN Neuro. 2020, 12, 1–23. [Google Scholar] [CrossRef]
- Zhao, X.; Sun, G.; Zhang, J.; Ting, S.; Gonzales, N.; Aronowski, J. Dimethyl fumarate protects brain from damage produced by intracerebral hemorrhage by mechanism involving Nrf2. Stroke 2015, 46, 1923–1928. [Google Scholar] [CrossRef]
- Schipper, H.M.; Song, W.; Tavitian, A.; Cressatti, M. The sinister face of heme oxygenase-1 in brain aging and disease. Prog. Neurobiol. 2019, 172, 40–70. [Google Scholar] [CrossRef]
- Zhang, R.; Kang, K.A.; Piao, M.J.; Maeng, Y.H.; Lee, K.H.; Chang, W.Y.; You, H.J.; Kim, J.S.; Kang, S.S.; Hyun, J.W. Cellular protection of morin against the oxidative stress induced by hydrogen peroxide. Chem. Biol. Interact. 2009, 177, 21–27. [Google Scholar] [CrossRef]
- Kuo, H.M.; Chang, L.S.; Lin, Y.L.; Lu, H.F.; Yang, J.S.; Lee, J.H.; Chung, J.-G. Morin inhibits the growth of human leukemia HL-60 cells via cell cycle arrest and induction of apoptosis through mitochondria dependent pathway. Anticancer Res. 2007, 27, 395–406. [Google Scholar]
- Kapoor, R.; Kakkar, P. Protective Role of Morin, a Flavonoid, against High Glucose Induced Oxidative Stress Mediated Apoptosis in Primary Rat Hepatocytes. PLoS ONE 2012, 7, e41663. [Google Scholar] [CrossRef]
- Bera, T.K.; Kar, S.K.; Yadav, P.K.; Mukherjee, P.; Yadav, S.; Joshi, B. Effects of monosodium glutamate on human health: A systematic review. World J. Pharm. Res. 2017, 5, 139–144. [Google Scholar]
- Ali, M.M.; Bawari, M.; Misra, U.; Babu, G. Locomotor and learning deficits in adult rats exposed to monosodium-L-glutamate during early life. Neurosci. Lett. 2000, 284, 57–60. [Google Scholar] [CrossRef]
- Olvera-Cortes, E.; Lopez-Vazquez, M.; Beas-Zarate, C.; Gonzalez-Burgos, I. Neonatal exposure to monosodium glutamate disrupts place learning ability in adult rats. Pharmacol. Biochem. Behav. 2005, 82, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Compton, D.M. Behavior strategy learning in rat: Effects of lesions of the dorsal striatum or dorsal hippocampus. Behav. Processes. 2004, 67, 335–342. [Google Scholar] [CrossRef]
- Husain, M.; Mehta, M.A. Cognitive enhancement by drugs in health and disease. TiCS 2011, 15, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Rodriguez, J.J.; Molina-Gil, S.; Rey-Brea, R.; Berrocoso, E.; Garcia-Alloza, M. Specific serotonergic denervation affects tau pathology and cognition without altering senile plaques deposition in APP/PS1 mice. PLoS ONE 2013, 8, e79947. [Google Scholar] [CrossRef]
- Gonçalves Rosa, S.; Quines, C.B.; Stangherlin, E.C.; Nogueira, C.W. Diphenyl diselenide ameliorates monosodium glutamate induced anxiety-like behavior in rats by modulating hippocampal BDNF-Akt pathway and uptake of GABA and serotonin neurotransmitters. Physiol. Behav. 2016, 155, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ciranna, L. Serotonin as a Modulator of Glutamate- and GABA-Mediated Neurotransmission: Implications in Physiological Functions and in Pathology. Curr. Neuropharmacol. 2006, 4, 101–114. [Google Scholar] [CrossRef]
- Quines, C.B.; Rosa, S.G.; Da Rocha, J.T.; Gai, B.M.; Bortolatto, C.F.; Duarte, M.M.M.; Nogueira, C.W. Monosodium glutamate, a food additive, induces depressive-like and anxiogenic-like behaviors in young rats. Life Sci. 2014, 107, 27–31. [Google Scholar] [CrossRef]
- De Pittà, M.; Goldberg, M.; Volman, V.; Berry, H.; Ben-Jacob, E. Glutamate regulation of calcium and IP3 oscillating and pulsating dynamics in astrocytes. J. Biol. Phys. 2009, 35, 383–411. [Google Scholar] [CrossRef]
- Madreiter-Sokolowski, C.T.; Thomas, C.; Ristow, M. Interrelation between ROS and Ca2+ in aging and age-related diseases. Redox Biol. 2020, 36, 78–135. [Google Scholar] [CrossRef] [PubMed]
- Onaolapo, O.J.; Aremu, O.S.; Onaolapo, A.Y. Monosodium glutamate-associated alterations in open field, anxiety-related and conditioned place preference behaviours in mice. Naunyn Schmiedebergs Arch. Pharmacol. 2017, 390, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Pauls, D.L.; Abramovitch, A.; Rauch, S.L.; Geller, D.A. Obsessive compulsive disorder: An integrative genetic and neurobiological perspective. Nat. Rev. Neurosci. 2014, 15, 410–424. [Google Scholar] [CrossRef] [PubMed]
- Farombi, E.O.; Onyema, O.O. Monosodium glutamate-induced oxidative damage and genotoxicity in the rat: Modulatory role of vitamin C, vitamin E and quercetin. Hum. Exp. Toxicol. 2006, 25, 251–259. [Google Scholar] [CrossRef]
- Singh, K.; Ahluwalia, P. Effect of monosodium glutamate on lipid peroxidation and certain antioxidant enzymes in cardiac tissue of alcoholic adult male mice. J. Cardiovasc. Dis. Res. 2012, 3, 12–18. [Google Scholar] [CrossRef]
- Zhang, M.; An, C.; Gao, Y.; Leak, R.K.; Chen, J.; Zhang, F. Emerging roles of Nrf2 and phase II antioxidant enzymes in neuroprotection. Prog. Neurobiol. 2013, 100, 30–47. [Google Scholar] [CrossRef]
- Ranjan, K.; Pathak, C. FADD regulates NF-κB activation and promotes ubiquitination of cFLIPL to induce apoptosis. Sci. Rep. 2016, 6, 22787–22801. [Google Scholar] [CrossRef]
- Abbott, N.J.; Rönnbäck, L.; Hansson, E. Astrocyte-endothelial interactions at the blood-brain barrier. Nat. Rev. Neurosci. 2006, 7, 41–53. [Google Scholar] [CrossRef]
- Caselli, A.; Cirri, P.; Santi, A.; Paoli, P. Morin: A promising natural drug. Curr. Med. Chem. 2016, 23, 774–791. [Google Scholar] [CrossRef]
- Lee, K.M.; Lee, Y.; Chun, H.J.; Kim, A.H.; Kim, J.Y.; Lee, J.Y.; Ishigami, A.; Lee, J. Neuroprotective and anti-inflammatory effects of morin in a murine model of Parkinson’s disease. J. Neurosci. Res. 2016, 94, 865–878. [Google Scholar] [CrossRef]
- Naowaboot, J.; Wannasiri, S.; Pannangpetch, P. Morin attenuates hepatic insulin resistance in high-fat-diet-induced obese mice. J. Physiol. Biochem. 2016, 72, 269–280. [Google Scholar] [CrossRef]
- Rizvi, F.; Mathur, A.; Kakkar, P. Morin mitigates acetaminophen induced liver injury by potentiating Nrf2 regulated survival mechanism through molecular intervention in PHLPP2-Akt-Gsk3beta axis. Apoptosis 2015, 20, 1296–1306. [Google Scholar] [CrossRef] [PubMed]
- Gu, M.; Zhang, Y.; Liu, C.; Wang, D.; Feng, L.; Fan, S.; Yang, B.; Tong, Q.; Ji, G.; Huang, C. Morin, a novel liver X receptor alpha/beta dual antagonist, has potent therapeutic efficacy for nonalcoholic fatty liver diseases. Br. J. Pharmacol. 2017, 174, 3032–3044. [Google Scholar] [CrossRef] [PubMed]
- Perumal, N.; Perumal, M.; Halagowder, D.; Sivasithamparam, N. Morin attenuates diethylnitrosamine-induced rat liver fibrosis and hepatic stellate cell activation by co-ordinated regulation of Hippo/Yap and TGFbeta1/Smad signaling. Biochimie 2017, 140, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.; Li, Z.; Shen, B.; Zhang, Q.; Feng, H. Protective effects of morin on lipopolysaccharide/d-galactosamine-induced acute liver injury by inhibiting TLR4/NF-kappaB and activating Nrf2/HO-1 signaling pathways. Int. Immunopharmacol. 2017, 45, 148–155. [Google Scholar] [CrossRef]
- Li, X.; Jin, Q.; Yao, Q.; Xu, B.; Li, L.; Zhang, S.; Tu, C. The flavonoid quercetin ameliorates liver inflammation and fibrosis by regulating hepatic macrophages activation and polarization in mice. Front. Pharmacol. 2018, 9, 72. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, D.M.; Gu, T.T.; Ding, X.Q.; Fan, C.Y.; Zhu, Q.; Shi, Y.; Hong, Y.; Kong, L. Morin reduces hepatic inflammation-associated lipid accumulation in high fructose-fed rats via inhibiting sphingosine kinase 1/sphingosine 1-phosphate signaling pathway. Biochem. Pharmacol. 2013, 86, 1791–1804. [Google Scholar] [CrossRef]
- Elizabeth, A.; Adegbuyi, A.; Olusegun, A.; Benneth, B.; Anthony, E.; Abayomi, A.; Solomon, U. Morin hydrate attenuates chronic stress-induced memoryimpairment and degeneration of hippocampal subfields in mice: The role of oxidative, nitrergic and neuroinflammatory pathways. Metab. Brain Dis. 2020, 35, 1145–1156. [Google Scholar] [CrossRef]
- Zhou, Y.; Cao, Z.; Wang, H.; Cheng, Y.; Yu, L.; Zhang, X.; Sun, Y.; Guo, X. The anti-inflammatory effects of Morin hydrate in atherosclerosis is associated with autophagy induction through cAMP signaling. Mol. Nutr. Food Res. 2017, 61, 1–10. [Google Scholar] [CrossRef]
- Onaolapo, A.Y.; Odetunde, I.; Akintola, A.S.; Ogundeji, M.O.; Ajao, A.; Obelawo, A.Y.; Onaolapo, O.J. Dietary composition modulates impact of food-added monosodium glutamate on behaviour, metabolic status and cerebral cortical morphology in mice. Biomed. Pharmacother. 2019, 109, 417–428. [Google Scholar] [CrossRef]
- Kiruthika, R.; Hema, T.; Justin Thenmozhi, A. Neuroprotective effect of morin against unpredictable chronic mild stress induced oxidative stress and behavioural deficits in Wistar rats. Int. J. Res. Pharm. Sci. 2020, 11, 1148–1154. [Google Scholar] [CrossRef]
- Ali, A.A.; Ezzeldin, E. Evaluation of the effect of caffeine and nicotine on postnatal neuorobehavioral and functional development in normally-fed and protein malnourished rats. ESCTJ 2004, 31, 77–89. [Google Scholar]
- Khaled Abd-Elhaleim El Azazy, M.; Kamel Mohamed, E.A.; Ismail Abo El-Fadl, H.M.; Abd El-Razik, F.H.; Abu Elfotuh, K. Omega-3 Rich Oils Attenuate ADHD-Like Behaviour Induced by Dietary Monosodium Glutamate in Rats. Pak. J. Biol. Sci. 2021, 24, 868–880. [Google Scholar] [CrossRef] [PubMed]
- Beppe, G.J.; Dongmo, A.B.; Foyet, H.S.; Tsabang, N.; Olteanu, Z.; Cioanca, O.; Hancianu, M.; Dimo, T.; Hritcu, L. Memory-enhancing activities of the aqueous extract of Albizia adianthifolia leaves in the 6-hydroxydopamine-lesion rodent model of parkinson’s disease. BMC Complement. Med. Ther. 2014, 14, 142. [Google Scholar] [CrossRef] [PubMed]
- Vorhess, C.V.; Klein, K.L.; Scott, W.I. Asprin-induced psychoteratogenesis in rats as a function of embryonic age. Teratog. Carcinog. Mutagen. 1982, 2, 77–84. [Google Scholar] [CrossRef]
- Stanford, S.C. The Open Field Test: Reinventing the wheel. J. Psychopharmacol. 2007, 21, 134–145. [Google Scholar] [CrossRef]
- Valrance, M.E.; Brunet, A.H.; Welsh, J. Vitamin D receptor-dependent inhibition of mammary tumor growth by EB1089 and ultraviolet radiation in vivo. Endocrinology 2007, 148, 4887–4894. [Google Scholar] [CrossRef]
- Alisik, M.; Neselioglu, S.; Erel, O. A colorimetric method to measure oxidized, reduced and total glutathione levels in erythrocytes. Lab. Med. 2019, 43, 269–277. [Google Scholar] [CrossRef]
- Glare, E.; Divjak, M.; Bailey, M.; Walters, E.H. beta-Actin and GAPDH housekeeping gene expression in asthmatic airways is variable and not suitable for normalizing mRNA levels. Thorax 2002, 57, 765–770. [Google Scholar] [CrossRef]
- Stahlberg, A.; Hakansson, J.; Xian, X.; Semb, H.; Kubista, M. Properties of the reverse transcription reaction in mRNA quantification. Clin. Chem. 2004, 50, 509–515. [Google Scholar] [CrossRef]
- Briton-Jones, C.; Lok, I.H.; Po, A.L.S.; Cheung, C.K.; Chiu, T.T.Y.; Haines, C. Changes in the ratio of Bax and Bcl-2 mRNA expression and their cellular localization throughout the ovulatory cycle in the human oviduct. J. Assist. Reprod. Genet. 2006, 23, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques; Churchill Livingstone: London, UK, 2008. [Google Scholar]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Groups | Bax (Arbitrary Units) | Bcl-2 (Arbitrary Units) | Bax/Bcl-2 ratio |
|---|---|---|---|
| NF | 1.383 | 1.074 | 1.28 |
| PM | 2.250 | 0.827 | 2.72 |
| MSG | 4.367 | 0.181 | 24.12 |
| PM + MSG | 7.483 | 0.036 | 207.86 |
| PM + MH | 1.433 | 1.003 | 1.43 |
| MSG + MH | 2.850 | 0.841 | 3.39 |
| PM + MSG + MH | 4.550 | 0.588 | 8.42 |
| Gene | Forward and Backward Primer Sequence |
|---|---|
| Bax | Forward: 5′-ATGTTTTCTGACGGCAACTTC-3′ Reverse: 5′-AGTCCAATGTCCAGCCCAT-3′ |
| Bcl-2 | Forward: 5′-CTACGAGTGGGATGCTGGAG-3′ Reverse: 5′- TTCTTCACGATGGTGAGCG-3′ |
| β-actin | Forward: 5′-GGTCGGTGTGAACGGATTTGG-3′ Reverse: 5′-ATGTAGGCCATGAGGTCCACC-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salem, H.A.; Elsherbiny, N.; Alzahrani, S.; Alshareef, H.M.; Abd Elmageed, Z.Y.; Ajwah, S.M.; Hamdan, A.M.E.; Abdou, Y.S.; Galal, O.O.; El Azazy, M.K.A.; et al. Neuroprotective Effect of Morin Hydrate against Attention-Deficit/Hyperactivity Disorder (ADHD) Induced by MSG and/or Protein Malnutrition in Rat Pups: Effect on Oxidative/Monoamines/Inflammatory Balance and Apoptosis. Pharmaceuticals 2022, 15, 1012. https://doi.org/10.3390/ph15081012
Salem HA, Elsherbiny N, Alzahrani S, Alshareef HM, Abd Elmageed ZY, Ajwah SM, Hamdan AME, Abdou YS, Galal OO, El Azazy MKA, et al. Neuroprotective Effect of Morin Hydrate against Attention-Deficit/Hyperactivity Disorder (ADHD) Induced by MSG and/or Protein Malnutrition in Rat Pups: Effect on Oxidative/Monoamines/Inflammatory Balance and Apoptosis. Pharmaceuticals. 2022; 15(8):1012. https://doi.org/10.3390/ph15081012
Chicago/Turabian StyleSalem, Hoda A., Nehal Elsherbiny, Sharifa Alzahrani, Hanan M. Alshareef, Zakaria Y. Abd Elmageed, Sadeem M. Ajwah, Ahmed M. E. Hamdan, Yahia S. Abdou, Omneya O. Galal, Marwa K. A. El Azazy, and et al. 2022. "Neuroprotective Effect of Morin Hydrate against Attention-Deficit/Hyperactivity Disorder (ADHD) Induced by MSG and/or Protein Malnutrition in Rat Pups: Effect on Oxidative/Monoamines/Inflammatory Balance and Apoptosis" Pharmaceuticals 15, no. 8: 1012. https://doi.org/10.3390/ph15081012
APA StyleSalem, H. A., Elsherbiny, N., Alzahrani, S., Alshareef, H. M., Abd Elmageed, Z. Y., Ajwah, S. M., Hamdan, A. M. E., Abdou, Y. S., Galal, O. O., El Azazy, M. K. A., & Abu-Elfotuh, K. (2022). Neuroprotective Effect of Morin Hydrate against Attention-Deficit/Hyperactivity Disorder (ADHD) Induced by MSG and/or Protein Malnutrition in Rat Pups: Effect on Oxidative/Monoamines/Inflammatory Balance and Apoptosis. Pharmaceuticals, 15(8), 1012. https://doi.org/10.3390/ph15081012