



Binding-Induced Diversity of a Human Telomeric G-Quadruplex Stability Phase Space

Abstract
:1. Introduction
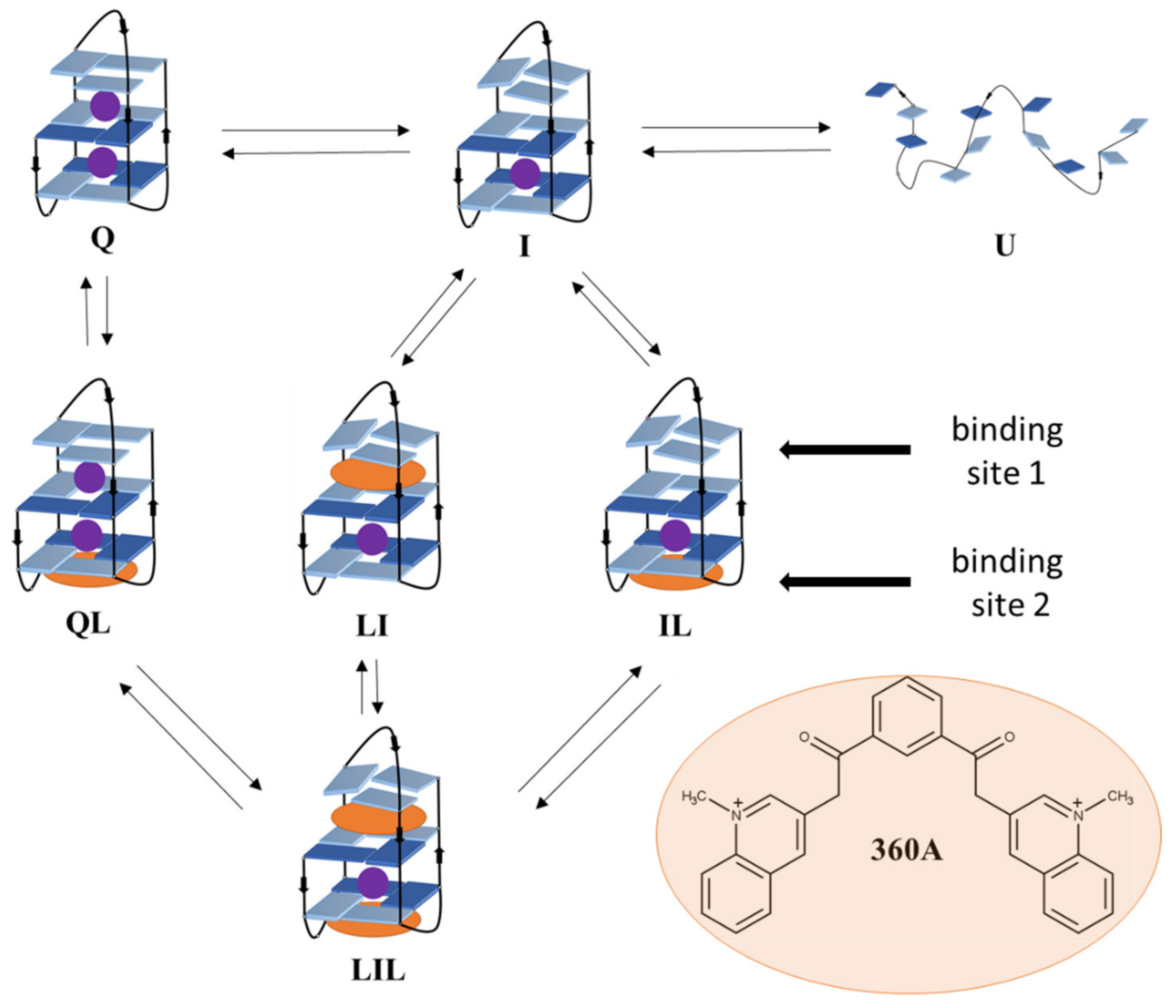
2. Results and Discussion
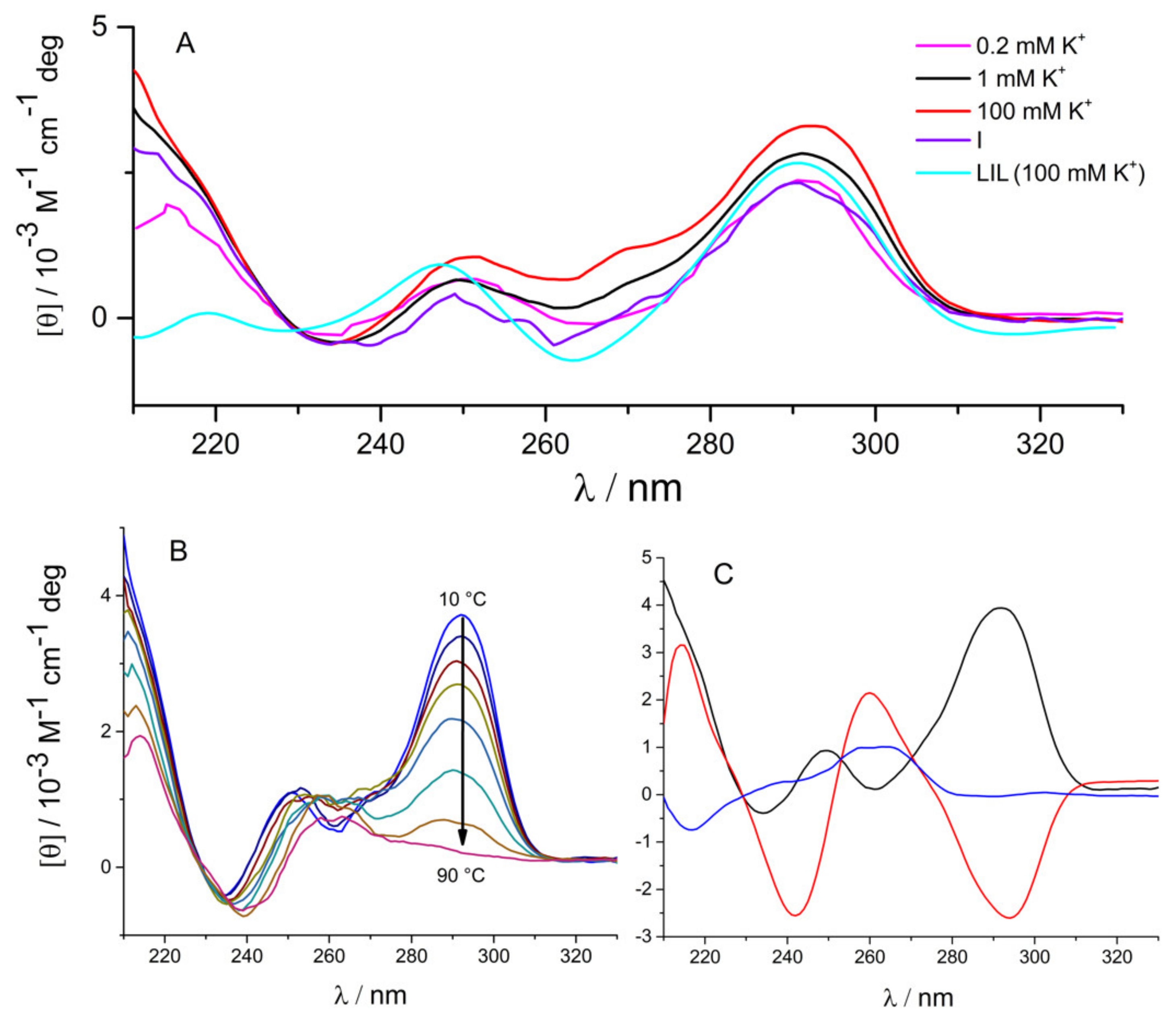
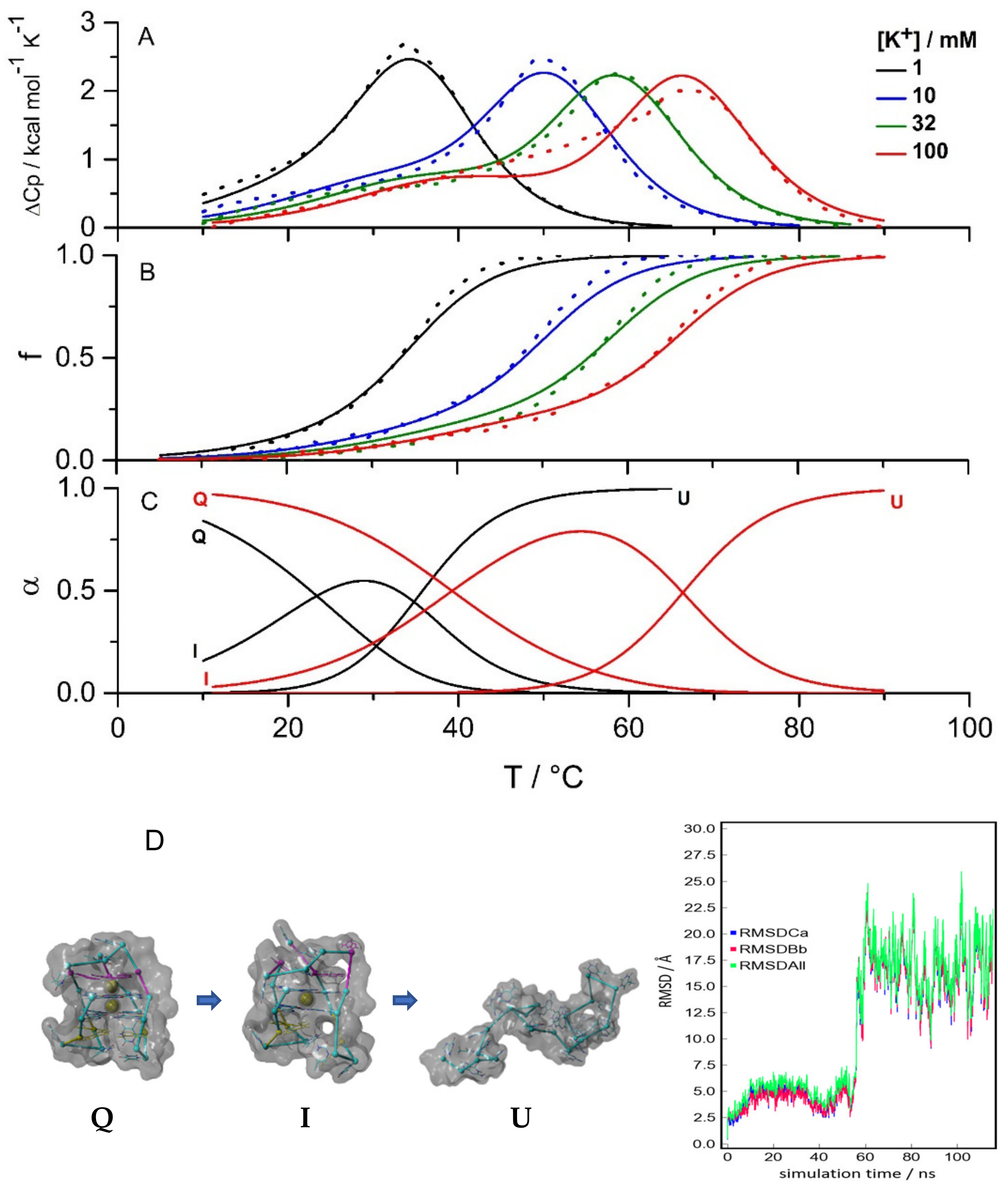
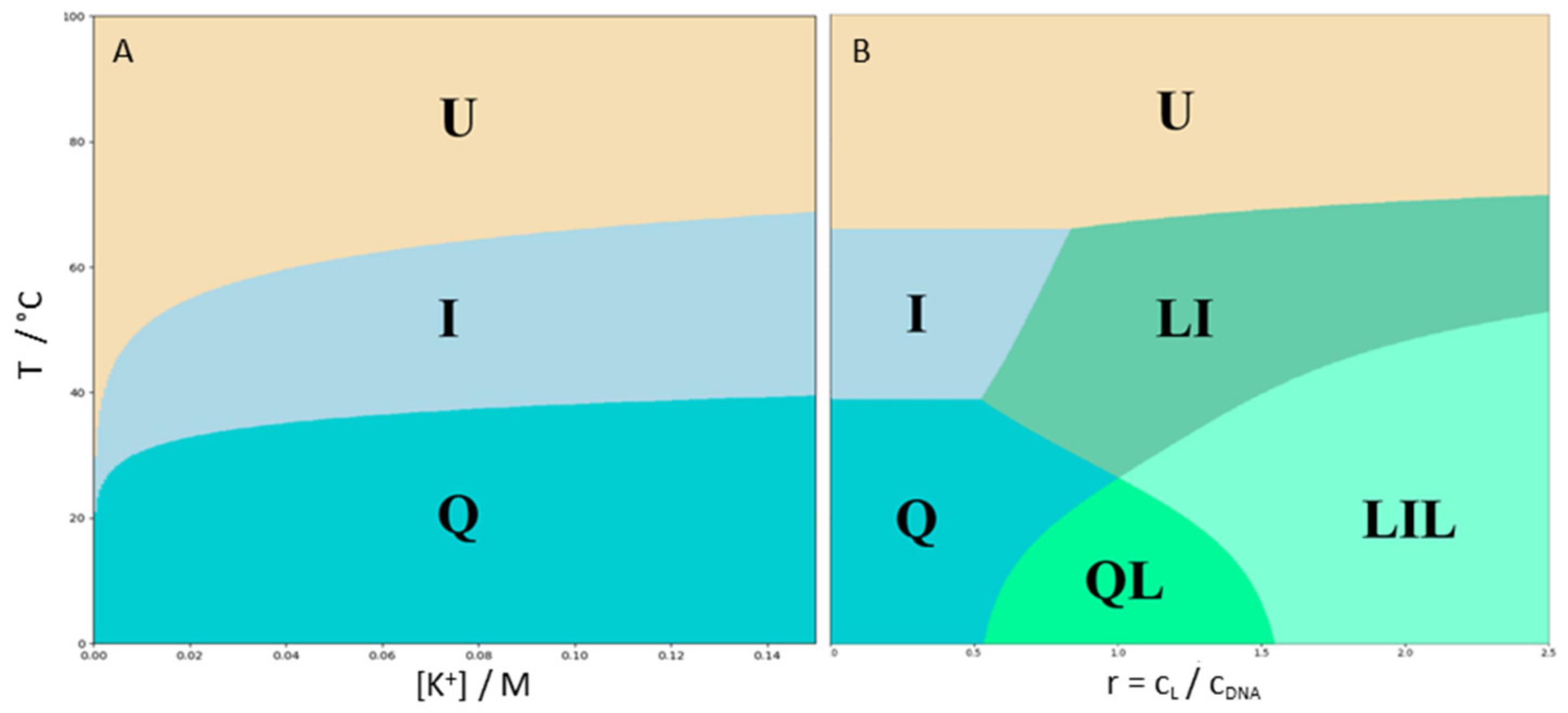
2.1. Three Significant Species Are Populated during Melting in the Absence of Ligand
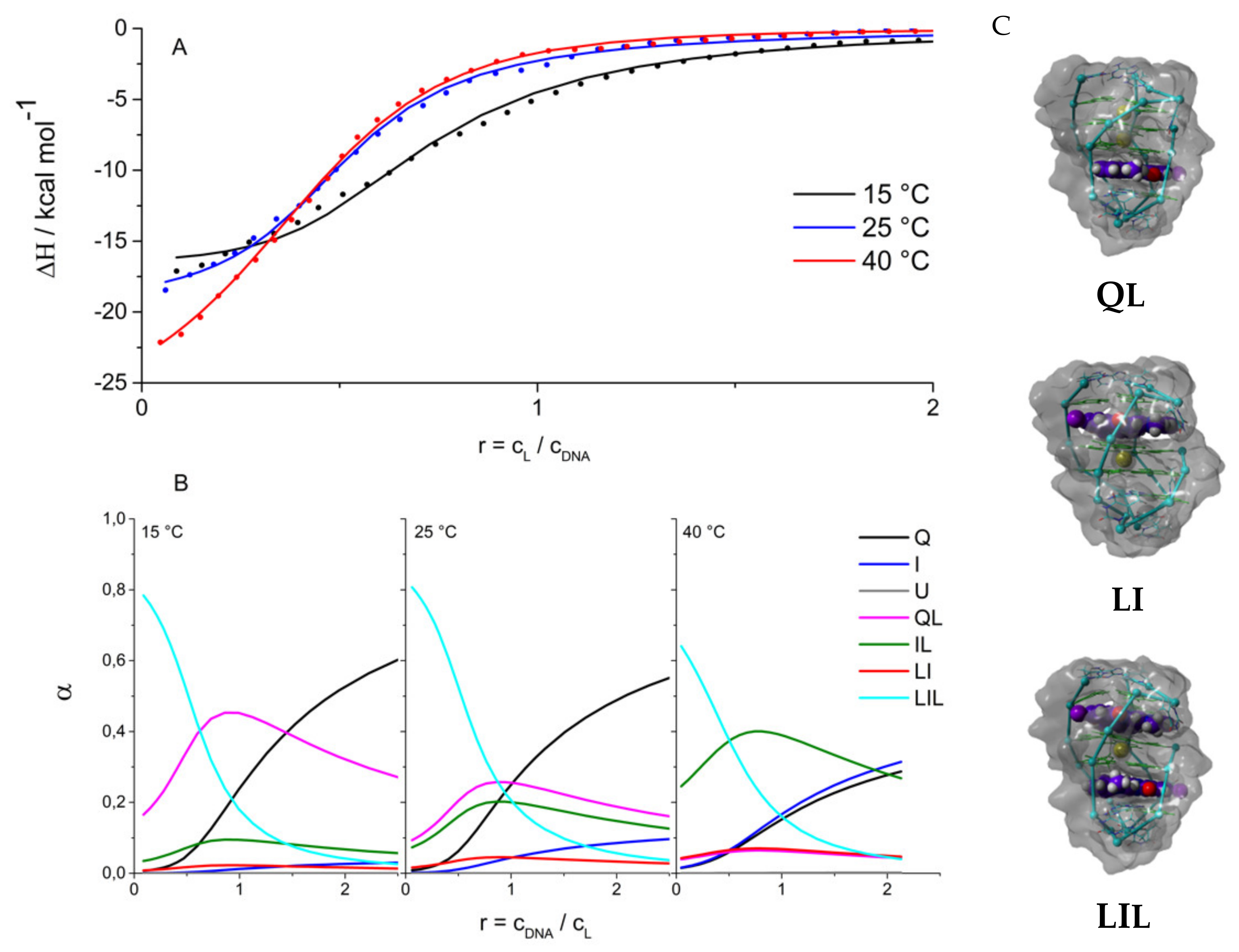
2.2. Driving Forces of Ligand Binding
2.3. Ligand Binding Alters the Stability Phase Space of 22GT
3. Materials and Methods
3.1. Sample Preparation
3.2. Circular Dichroism Spectroscopy
3.3. Differential Scanning Calorimetry
3.4. Isothermal Titration Calorimetry
3.5. Molecular Modelling
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Crick, F.; Watson, J. Molecular Structure of Nucleic Acids: A Structure for Deoxyribose Nucleic Acid. Nature 1953, 171, 737–738. [Google Scholar]
- Gellert, M.; Lipsett, M.N.; Davies, D.R. Helix Formation by Guanylic Acid. Proc. Natl. Acad. Sci. USA 1962, 48, 2013–2018. [Google Scholar] [CrossRef] [PubMed]
- Arnott, S.; Chandrasekaran, K.; Marttila, C.M. Structures for Polyinosinic Acid and Polyguanylic Acid. Biochem. J. 1974, 141, 537–543. [Google Scholar] [CrossRef] [PubMed]
- Sen, D.; Gilbert, W. Formation of Parallel Four-Stranded Complexes by Guanine-Rich Motifs in DNA and Its Implications for Meiosis. Nature 1988, 334, 364–366. [Google Scholar] [CrossRef] [PubMed]
- Huppert, J.L.; Balasubramanian, S. G-Quadruplexes in Promoters throughout the Human Genome. Nucleic Acids Res. 2007, 35, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Sundquist, W.I.; Klug, A. Telomeric DNA Dimerizes by Formation of Guanine Tetrads between Hairpin Loops. Nature 1989, 342, 825–829. [Google Scholar] [CrossRef]
- Brooks, T.A.; Kendrick, S.; Hurley, L. Making Sense of G-Quadruplex and i-Motif Functions in Oncogene Promoters. FEBS J. 2010, 277, 3459–3469. [Google Scholar] [CrossRef]
- Collie, G.W.; Parkinson, G.N. The Application of DNA and RNA G-Quadruplexes to Therapeutic Medicines. Chem. Soc. Rev. 2011, 40, 5867–5892. [Google Scholar] [CrossRef]
- Day, H.A.; Pavlou, P.; Waller, Z.A.E. I-Motif DNA: Structure, Stability and Targeting with Ligands. Bioorg. Med. Chem. 2014, 22, 4407–4418. [Google Scholar] [CrossRef]
- Benabou, S.; Aviñó, A.; Eritja, R.; González, C.; Gargallo, R. Fundamental Aspects of the Nucleic Acid I-Motif Structures. RSC Adv. 2014, 4, 26956–26980. [Google Scholar] [CrossRef]
- Alba, J.J.; Sadurní, A.; Gargallo, R. Nucleic Acid I-Motif Structures in Analytical Chemistry. Crit. Rev. Anal. Chem. 2016, 46, 443–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biffi, G.; Tannahill, D.; McCafferty, J.; Balasubramanian, S. Quantitative Visualization of DNA G-Quadruplex Structures in Human Cells. Nat. Chem. 2013, 5, 182–186. [Google Scholar] [CrossRef] [PubMed]
- Zeraati, M.; Langley, D.B.; Schofield, P.; Moye, A.L.; Rouet, R.; Hughes, W.E.; Bryan, T.M.; Dinger, M.E.; Christ, D. I-Motif DNA Structures Are Formed in the Nuclei of Human Cells. Nat. Chem. 2018, 10, 631–637. [Google Scholar] [CrossRef]
- Kocman, V.; Plavec, J. A Tetrahelical DNA Fold Adopted by Tandem Repeats of Alternating GGG and GCG Tracts. Nat. Commun. 2014, 5, 5831. [Google Scholar] [CrossRef]
- Kocman, V.; Plavec, J. Tetrahelical Structural Family Adopted by AGCGA-Rich Regulatory DNA Regions. Nat. Commun. 2017, 8, 15355. [Google Scholar] [CrossRef]
- Hadži, S.; Kocman, V.; Oblak, D.; Plavec, J.; Lah, J. Energetic Basis of AGCGA-Rich DNA Folding into a Tetrahelical Structure. Angew. Chem. Int. Ed. 2019, 58, 2387–2391. [Google Scholar] [CrossRef]
- Zahner, A.M.; Williamson, J.R.; Cech, T.R.; Prescott, D.M. Inhibition of Telomerase by G-Quartet DNA Structures. Nature 1991, 350, 718–720. [Google Scholar]
- Agarwal, T.; Pradhan, D.; Géci, I.; El-Madani, A.M.; Petersen, M.; Pedersen, E.B.; Maiti, S. Improved Inhibition of Telomerase by Short Twisted Intercalating Nucleic Acids under Molecular Crowding Conditions. Nucleic Acid Ther. 2012, 22, 380–390. [Google Scholar] [CrossRef]
- Phan, A.T.; Kuryavyi, V.; Gaw, H.Y.; Patel, D.J. Small-Molecule Interaction with a Five-Guanine-Tract g-Quadruplex Structure from the Human Myc Promoter. Nat. Chem. Biol. 2005, 1, 167–173. [Google Scholar] [CrossRef]
- De Cian, A.; DeLemos, E.; Mergny, J.L.; Teulade-Fichou, M.P.; Monchaud, D. Highly Efficient G-Quadruplex Recognition by Bisquinolinium Compounds. J. Am. Chem. Soc. 2007, 129, 1856–1857. [Google Scholar] [CrossRef]
- Balasubramanian, S.; Hurley, L.H.; Neidle, S. Targeting G-Quadruplexes in Gene Promoters: A Novel Anticancer Strategy? Nat. Rev. Drug Discov. 2011, 10, 261–275. [Google Scholar] [CrossRef]
- Eddy, J.; Maizels, N. Gene Function Correlates with Potential for G4 DNA Formation in the Human Genome. Nucleic Acids Res. 2006, 34, 3887–3896. [Google Scholar] [CrossRef] [Green Version]
- Riou, J.F. G-Quadruplex Interacting Agents Targetting the Telomeric G-Overhang Are More than Simple Telomerase Inhibitors. Curr. Med. Chem. Anti-Cancer Agents 2004, 4, 439–443. [Google Scholar] [CrossRef]
- Gomez, D.; Wenner, T.; Brassart, B.; Douarre, C.; O’Donohue, M.F.; El Khoury, V.; Shin-Ya, K.; Morjani, H.; Trentesaux, C.; Riou, J.F. Telomestatin-Induced Telomere Uncapping Is Modulated by POT1 through G-Overhang Extension in HT1080 Human Tumor Cells. J. Biol. Chem. 2006, 281, 38721–38729. [Google Scholar] [CrossRef]
- Rodriguez, R.; Müller, S.; Yeoman, J.A.; Trentesaux, C.; Riou, J.F.; Balasubramanian, S. A Novel Small Molecule That Alters Shelterin Integrity and Triggers a DNA-Damage Response at Telomeres. J. Am. Chem. Soc. 2008, 130, 15758–15759. [Google Scholar] [CrossRef]
- Pagano, B.; Amato, J.; Iaccarino, N.; Cingolani, C.; Zizza, P.; Biroccio, A.; Novellino, E.; Randazzo, A. Looking for Efficient G-Quadruplex Ligands: Evidence for Selective Stabilizing Properties and Telomere Damage by Drug-like Molecules. ChemMedChem 2015, 10, 640–649. [Google Scholar] [CrossRef]
- Alcaro, S.; Musetti, C.; Distinto, S.; Casatti, M.; Zagotto, G.; Artese, A.; Parrotta, L.; Moraca, F.; Costa, G.; Ortuso, F.; et al. Identification and Characterization of New DNA G-Quadruplex Binders Selected by a Combination of Ligand and Structure-Based Virtual Screening Approaches. J. Med. Chem. 2013, 56, 843–855. [Google Scholar] [CrossRef]
- Webba Da Silva, M. Geometric Formalism for DNA Quadruplex Folding. Chemistry 2007, 13, 9738–9745. [Google Scholar] [CrossRef]
- Neidle, S.; Parkinson, G. Telomere Maintenance as a Target for Anticancer Drug Discovery. Nat. Rev. Drug Discov. 2002, 1, 383–393. [Google Scholar] [CrossRef]
- Ambrus, A.; Chen, D.; Dai, J.; Bialis, T.; Jones, R.A.; Yang, D. Human Telomeric Sequence Forms a Hybrid-Type Intramolecular G-Quadruplex Structure with Mixed Parallel/Antiparallel Strands in Potassium Solution. Nucleic Acids Res. 2006, 34, 2723–2735. [Google Scholar] [CrossRef]
- Gray, R.D.; Petraccone, L.; Trent, J.O.; Chaires, J.B. Characterization of a K+-Induced Conformational Switch in a Human Telomeric DNA Oligonucleotide Using 2-Aminopurine Fluorescence. Biochemistry 2010, 49, 179–194. [Google Scholar] [CrossRef]
- Phan, A.T.; Kuryavyi, V.; Luu, K.N.; Patel, D.J. Structure of Two Intramolecular G-Quadruplexes Formed by Natural Human Telomere Sequences in K+ Solution. Nucleic Acids Res. 2007, 35, 6517–6525. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Patel, D.J. Solution Structure of the Human Telomeric Repeat d[AG3(T2AG3)3] G-Tetraplex. Structure 1993, 1, 263–282. [Google Scholar] [CrossRef]
- Gray, R.D.; Buscaglia, R.; Chaires, J.B. Populated Intermediates in the Thermal Unfolding of the Human Telomeric Quadruplex. J. Am. Chem. Soc. 2012, 134, 16834–16844. [Google Scholar] [CrossRef]
- Bončina, M.; Lah, J.; Prislan, I.; Vesnaver, G. Energetic Basis of Human Telomeric DNA Folding into G-Quadruplex Structures. J. Am. Chem. Soc. 2012, 134, 9657–9663. [Google Scholar] [CrossRef]
- Bončina, M.; Vesnaver, G.; Chaires, J.B.; Lah, J. Unraveling the Thermodynamics of the Folding and Interconversion of Human Telomere G-Quadruplexes. Angew. Chem. Int. Ed. 2016, 55, 10340–10344. [Google Scholar] [CrossRef]
- Marchand, A.; Granzhan, A.; Iida, K.; Tsushima, Y.; Ma, Y.; Nagasawa, K.; Teulade-Fichou, M.P.; Gabelica, V. Ligand-Induced Conformational Changes with Cation Ejection upon Binding to Human Telomeric DNA G-Quadruplexes. J. Am. Chem. Soc. 2015, 137, 750–756. [Google Scholar] [CrossRef]
- Piazza, A.; Boulé, J.B.; Lopes, J.; Mingo, K.; Largy, E.; Teulade-Fichou, M.P.; Nicolas, A. Genetic Instability Triggered by G-Quadruplex Interacting Phen-DC Compounds in Saccharomyces Cerevisiae. Nucleic Acids Res. 2010, 38, 4337–4348. [Google Scholar] [CrossRef]
- Bončina, M.; Podlipnik, Č.; Piantanida, I.; Eilmes, J.; Teulade-Fichou, M.P.; Vesnaver, G.; Lah, J. Thermodynamic Fingerprints of Ligand Binding to Human Telomeric G-Quadruplexes. Nucleic Acids Res. 2015, 43, 10376–10386. [Google Scholar] [CrossRef]
- Bončina, M.; Hamon, F.; Islam, B.; Teulade-Fichou, M.P.; Vesnaver, G.; Haider, S.; Lah, J. Dominant Driving Forces in Human Telomere Quadruplex Binding-Induced Structural Alterations. Biophys. J. 2015, 108, 2903–2911. [Google Scholar] [CrossRef]
- Karsisiotis, A.I.; Hessari, N.M.A.; Novellino, E.; Spada, G.P.; Randazzo, A.; Webba Da Silva, M. Topological Characterization of Nucleic Acid G-Quadruplexes by UV Absorption and Circular Dichroism. Angew. Chem. Int. Ed. 2011, 50, 10645–10648. [Google Scholar] [CrossRef]
- Lim, K.W.; Amrane, S.; Bouaziz, S.; Xu, W.; Mu, Y.; Patel, D.J.; Luu, K.N.; Phan, A.T. Structure of the Human Telomere in K+ Solution: A Stable Basket-Type G-Quadruplex with Only Two G-Tetrad Layers. J. Am. Chem. Soc. 2009, 131, 4301–4309. [Google Scholar] [CrossRef] [Green Version]
- Marchand, A.; Gabelica, V. Folding and Misfolding Pathways of G-Quadruplex DNA. Nucleic Acids Res. 2016, 44, 10999–11012. [Google Scholar] [CrossRef]
- Gray, R.D.; Chaires, J.B. Analysis of Multidimensional G-Quadruplex Melting Curves. Curr. Protoc. Nucleic Acid Chem. 2011, 45, 1–17. [Google Scholar] [CrossRef]
- Pagano, B.; Randazzo, A.; Fotticchia, I.; Novellino, E.; Petraccone, L.; Giancola, C. Differential Scanning Calorimetry to Investigate G-Quadruplexes Structural Stability. Methods 2013, 64, 43–51. [Google Scholar] [CrossRef]
- Marchand, A.; Rosu, F.; Zenobi, R.; Gabelica, V. Thermal Denaturation of DNA G-Quadruplexes and Their Complexes with Ligands: Thermodynamic Analysis of the Multiple States Revealed by Mass Spectrometry. J. Am. Chem. Soc. 2018, 140, 12553–12565. [Google Scholar] [CrossRef]
- Jana, J.; Weisz, K. Thermodynamic Stability of G-Quadruplexes: Impact of Sequence and Environment. ChemBioChem 2021, 22, 2848–2856. [Google Scholar] [CrossRef]
- Santos, T.; Lopes-Nunes, J.; Alexandre, D.; Miranda, A.; Figueiredo, J.; Silva, M.S.; Mergny, J.L.; Cruz, C. Stabilization of a DNA Aptamer by Ligand Binding. Biochimie 2022, 200, 8–18. [Google Scholar] [CrossRef]
- Marchand, A.; Strzelecka, D.; Gabelica, V. Selective and Cooperative Ligand Binding to Antiparallel Human Telomeric DNA G-Quadruplexes. Chem. Eur. J. 2016, 22, 9551–9555. [Google Scholar] [CrossRef]
- Santos, T.; Salgado, G.F.; Cabrita, E.J.; Cruz, C. G-Quadruplexes and Their Ligands: Biophysical Methods to Unravel g-Quadruplex/Ligand Interactions. Pharmaceuticals 2021, 14, 769. [Google Scholar] [CrossRef]
- Liao, T.C.; Ma, T.Z.; Chen, S.B.; Cilibrizzi, A.; Zhang, M.J.; Li, J.H.; Zhou, C.Q. Human Telomere Double G-Quadruplex Recognition by Berberine-Bisquinolinium Imaging Conjugates in Vitro and in Cells. Int. J. Biol. Macromol. 2020, 158, 1299–1309. [Google Scholar] [CrossRef]
- Spolar, R.S.; Record, M.T. Coupling of Local Folding to Site-Specific Binding of Proteins to DNA. Science 1994, 263, 777–784. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, R.L. Temperature Dependence of the Hydrophobic Interaction in Protein Folding. Proc. Natl. Acad. Sci. USA 1986, 83, 8069–8072. [Google Scholar] [CrossRef]
- Lah, J.; Hadži, S. Conditional Cooperativity in DNA Minor-Groove Recognition by Oligopeptides. Molecules 2021, 26, 5188. [Google Scholar] [CrossRef]
- Cantor, C.R.; Warshaw, M.M.; Shapiro, H. Dichroism Studies of the Conformation of Deoxyoligonucleotides. Biopolymers 1970, 9, 1059–1077. [Google Scholar] [CrossRef]
- Krieger, E.; Vriend, G. New Ways to Boost Molecular Dynamics Simulations. J. Comput. Chem. 2015, 36, 996–1007. [Google Scholar] [CrossRef]
- Pérez, A.; Marchán, I.; Svozil, D.; Sponer, J.; Cheatham, T.E.; Laughton, C.A.; Orozco, M. Refinement of the AMBER Force Field for Nucleic Acids: Improving the Description of α/γ Conformers. Biophys. J. 2007, 92, 3817–3829. [Google Scholar] [CrossRef]
- MacKerell, A.D.; Bashford, D.; Bellott, M.; Dunbrack, R.L.; Evanseck, J.D.; Field, M.J.; Fischer, S.; Gao, J.; Guo, H.; Ha, S.; et al. All-Atom Empirical Potential for Molecular Modeling and Dynamics Studies of Proteins. J. Phys. Chem. B 1998, 102, 3586–3616. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2009, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 22GT | 22AG a | 22GT | 22AGa | 22GT | 22AG a | |
| /kcal mol−1 | −4.7 ± 0.7 | −3.7 ± 0.1 | −1.0 ± 0.2 | −1.4 ± 0.1 | −5.7 ± 0.9 | −5.1 ± 0.2 |
| /kcal mol−1 | −34 ± 4 | −23.6 ± 0.6 | −22 ± 5 | −22.4 ± 0.1 | −56 ± 9 | −46.0 ± 1.2 |
| /kcal mol−1 K−1 | −0.22 ± 0.1 | −0.42 ± 0.02 | −0.1 ± 0.1 | −0.0 ± 0.2 | −0.32± 0.2 | −0.42 ± 0.2 |
| /kcal mol−1 | −29 ± 4 | −19.9 ± 0.6 | −21 ± 5 | −21.0 ± 0.1 | −50 ± 9 | −40.9 ± 0.7 |
| −1.3 ± 0.1 | −1.4 ± 0.2 | −0.4 ± 0.2 | −0.7 ± 0.2 | −1.7 ± 0.3 | −2.1 ± 0.4 | |
| Site 1 | Site 2 | |
|---|---|---|
| /kcal mol−1 K−1 | −0.02 ± 0.01 | −0.16 ± 0.04 |
| /kcal mol−1 | −18 ± 4 | −20 ± 5 |
| /kcal mol−1 | −8 ± 4 | −11 ± 5 |
| /kcal mol−1 | −9.6 ± 0.6 | −8.7 ± 0.5 |
| ∆Gint/kcal mol−1 | −18 ± 4 | −20 ± 5 |
| ∆Ghyd/kcal mol−1 | −1.6 ± 0.2 | −12.8 ± 1.5 |
| ∆Gother/kcal mol−1 | 10 ± 4 | 24 ± 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oblak, D.; Hadži, S.; Podlipnik, Č.; Lah, J. Binding-Induced Diversity of a Human Telomeric G-Quadruplex Stability Phase Space. Pharmaceuticals 2022, 15, 1150. https://doi.org/10.3390/ph15091150
Oblak D, Hadži S, Podlipnik Č, Lah J. Binding-Induced Diversity of a Human Telomeric G-Quadruplex Stability Phase Space. Pharmaceuticals. 2022; 15(9):1150. https://doi.org/10.3390/ph15091150
Chicago/Turabian StyleOblak, Domen, San Hadži, Črtomir Podlipnik, and Jurij Lah. 2022. "Binding-Induced Diversity of a Human Telomeric G-Quadruplex Stability Phase Space" Pharmaceuticals 15, no. 9: 1150. https://doi.org/10.3390/ph15091150
APA StyleOblak, D., Hadži, S., Podlipnik, Č., & Lah, J. (2022). Binding-Induced Diversity of a Human Telomeric G-Quadruplex Stability Phase Space. Pharmaceuticals, 15(9), 1150. https://doi.org/10.3390/ph15091150