

Characterization of Orange Peel Extract and Its Potential Protective Effect against Aluminum Chloride-Induced Alzheimer’s Disease


 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
2.1. Phytochemical and HPLC Analysis
2.2. Biochemical Properties of OPE
2.2.1. Antioxidant Activity
2.2.2. Anti-Acetylcholinesterase Power
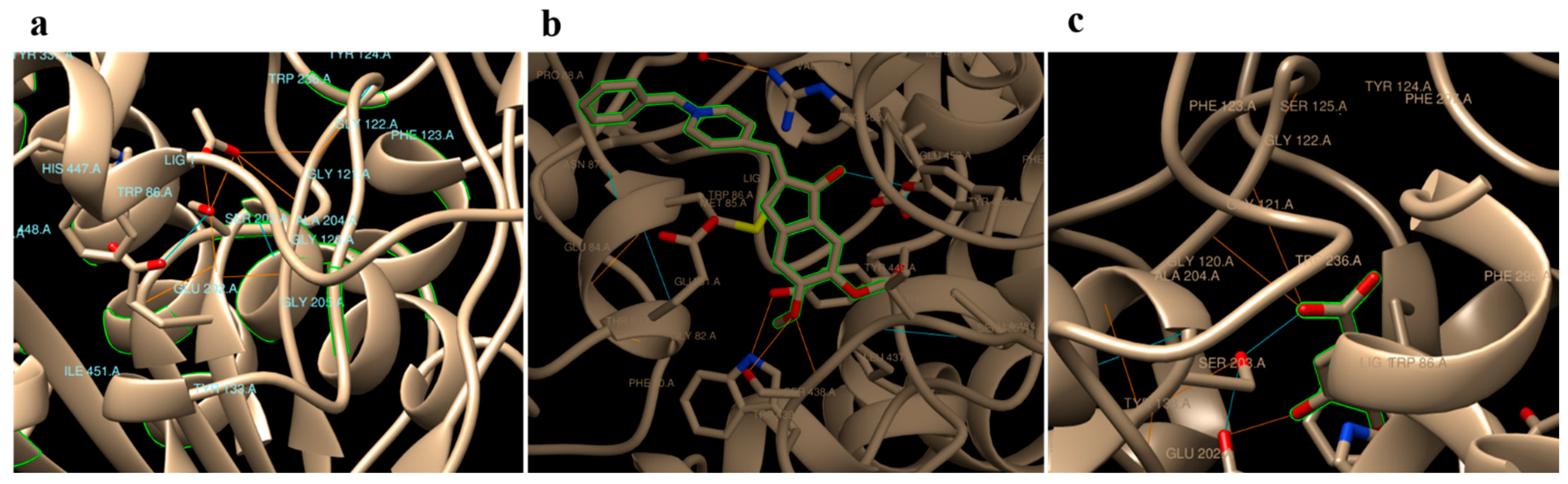
2.3. Computational Docking Analysis
2.4. In Vivo Anti Alzheimer Potentials
2.4.1. Effect of Donepezil, Gallic Acid and OPE Treatment on Brain Aβ42, Total Cholesterol and Phospholipids Levels
2.4.2. Effect of Donepezil, Gallic Acid, and OPE Treatment on Oxidative Stress Parameters in Brain
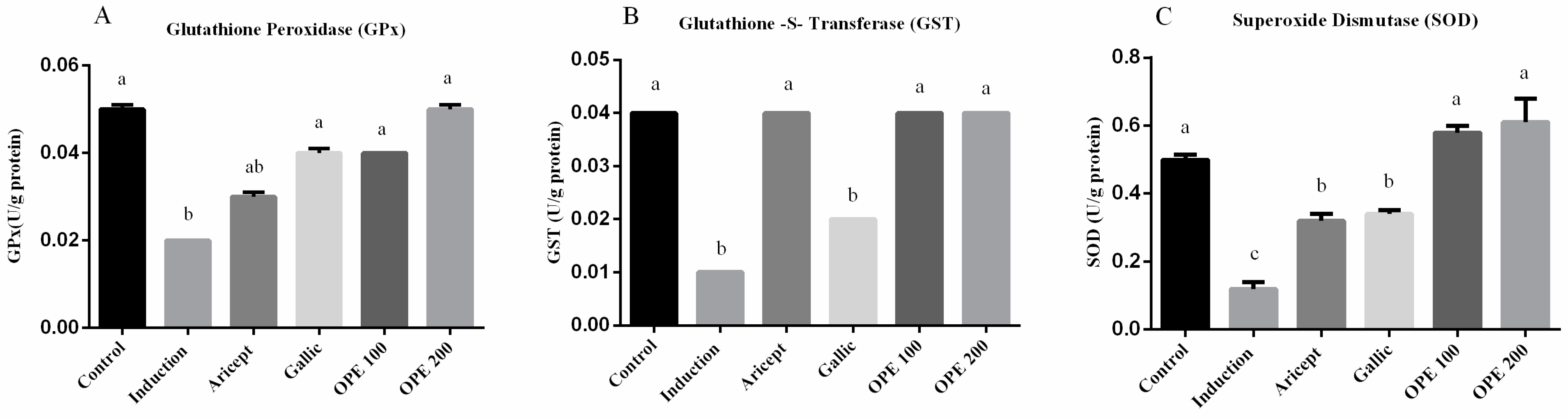
2.4.3. Effect of Donepezil, Gallic Acid, and OPE Treatment on Brain Antioxidant Enzymes
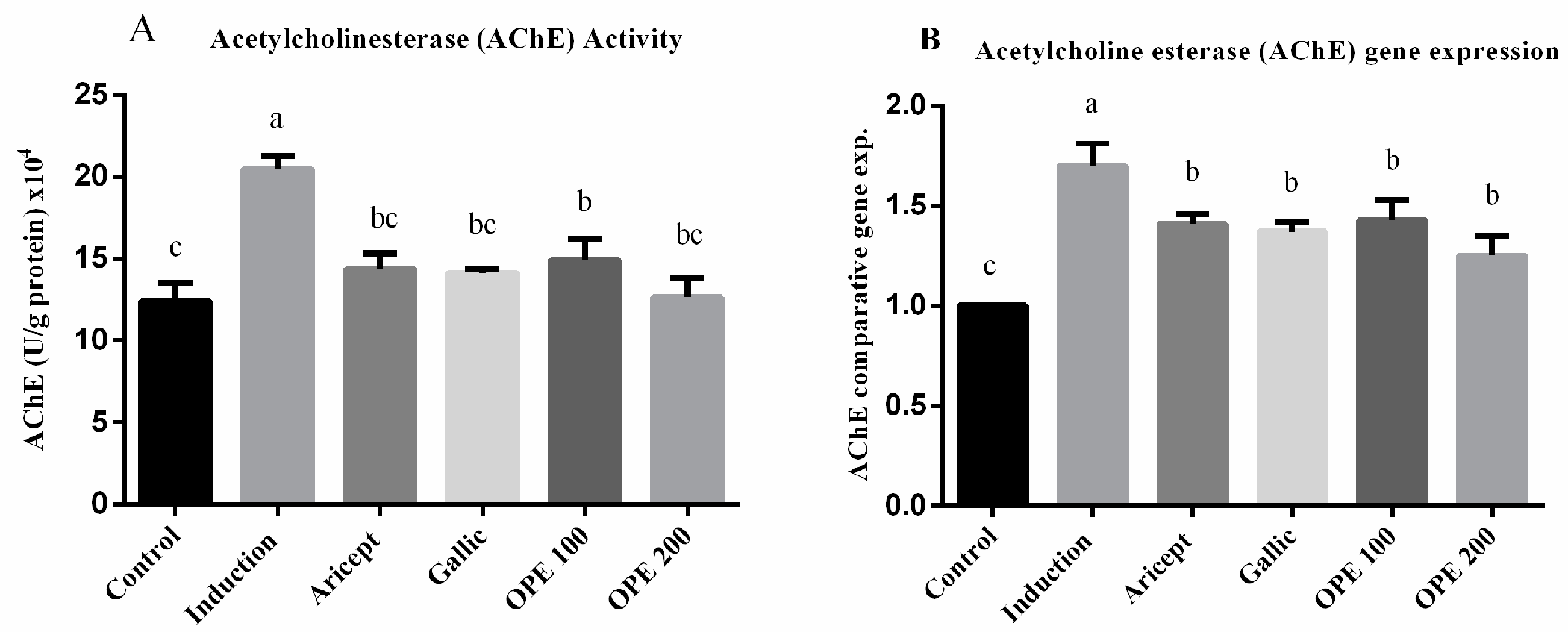
2.4.4. Effect of Donepezil, Gallic Acid, and OPE Treatment on Brain AChE Activity and Gene Expression Levels (Hypothalamus)
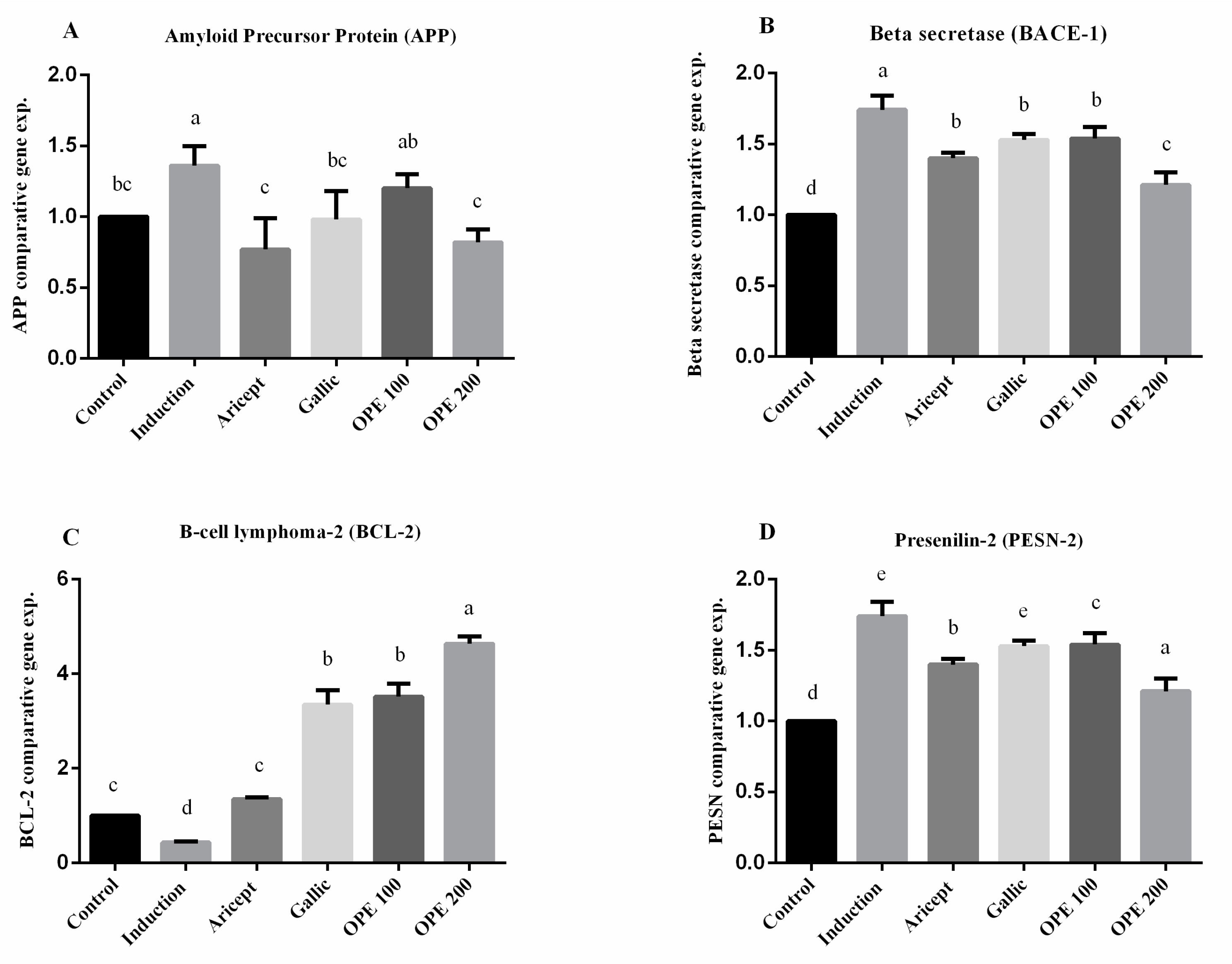
2.4.5. Brain (Hippocampus) Gene Expression
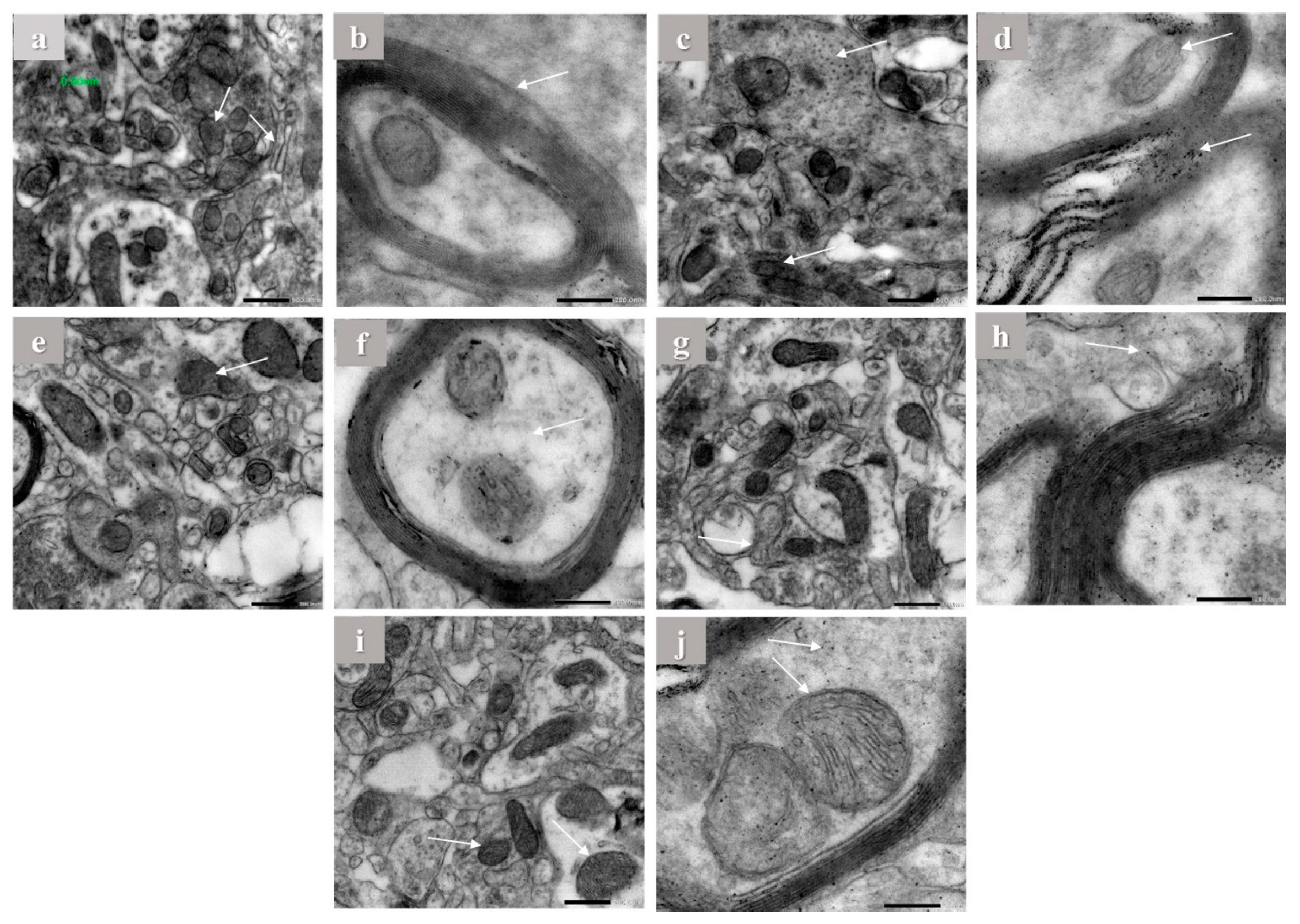
2.5. Electron Microscope Observations
3. Discussion
3.1. Phytochemicals Content and HPLC Analysis
3.2. Characterization of OPE
3.3. The Possible Protective Effect of OPE against AD Rat Model Induced by Aluminum Chloride Injection
3.4. Effect of Donepezil, Gallic Acid, and OPE Treatment on Brain Aβ42 Level, TC, and Phospholipids in the Brain
3.5. Effect of Donepezil, Gallic Acid, and OPE Treatment on Oxidative Stress Parameters in Brain
3.6. Effect of Donepezil, Gallic Acid, and OPE Treatment on Brain Antioxidant Enzymes
3.7. Brain AChE Activity and Its Gene Expression Level on Brain (Hypothalamus)
3.8. Brain (Hippocampus) Gene Expression
3.9. Electron Microscope
4. Material and Methods
4.1. Plant Materials
4.2. Preparation of Orange Peel Extract
Conventional Extraction (CE)
4.3. Phytochemical Analysis
4.3.1. Total Phenolic Content (TPC)
4.3.2. Total Flavonoid Assay (TFC)
4.4. HPLC Polyphenol Identification of Orange Peels Extract
4.5. Characterization of Orange Peels Extract
4.5.1. Antioxidant Activity
4.5.2. Anticholinesterase Potentials
4.6. Molecular Docking Studies
4.7. In Vivo Experiment of OPE Anti-Alzheimer Potentials
4.7.1. Experimental Design
4.7.2. Biochemical Parameters in Brain Homogenate
4.7.3. Hippocampal and Hypothalamic Tissue Separation and Total RNA Purification for qRT-PCR Analysis
4.8. Statistical Analysis of the Data
4.9. Ethical Approval
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tang, Y.; Zhang, D.; Gong, X.; Zheng, J. A mechanistic survey of Alzheimer’s disease. Biophys. Chem. 2022, 281, 106735. [Google Scholar] [CrossRef]
- Alzheimer’s Association. Alzheimer’s disease facts and figures. Alzheimer’s Dement. 2021, 17, 327–406. [Google Scholar]
- World Alzheimer Report. Life after Diagnosis: Navigating Treatment, Care and Support, 2022. Available online: https://www.alzint.org/u/World-Alzheimer-Report-2022.pdf (accessed on 18 November 2022).
- Akwa, Y. Steroids and Alzheimer’s Disease: Changes Associated with Pathology and Therapeutic Potential. Int. J. Mol. Sci. 2022, 7, 21. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, K.; Liu, F.; Gong, C.X. Tau and neurodegenerative disease: The story so far. Nat. Rev. Neurol. 2015, 12, 15–27. [Google Scholar] [CrossRef] [PubMed]
- Du, H. Cyclophilin D deficiency attenuates mitochondrial and neuronal perturbation and ameliorates learning and memory in Alzheimer’s disease. Nat. Med. 2008, 14, 1097–1105. [Google Scholar] [CrossRef]
- Tilokani, L.; Nagashima, S.; Paupe, V.; Prudent, J. Mitochondrial dynamics: Overview of molecular mechanisms. Essays Biochem. 2018, 62, 341–360. [Google Scholar]
- Khan, K.A.; Kumar, N.; Nayak, P.G.; Nampoothiri, M.; Shenoy, R.R.; Krishnadas, N.; Rao, M.; Mudgal, J. Impact of caffeic acid on aluminium chloride-induced dementia in rats. J. Pharm. Pharmacol. 2013, 65, 1745–1752. [Google Scholar] [CrossRef]
- Nie, J. Exposure to aluminum in daily life and Alzheimer’s disease. Adv. Exp. Med. Biol. 2018, 1091, 99–111. [Google Scholar]
- Kawahara, M.; Kato-Negishi, M. Link between aluminum and the pathogenesis of Alzheimer’s disease: The integration of the aluminum and amyloid cascade hypotheses. Int. J. Alzheimers Dis. 2011, 2011, 276393. [Google Scholar] [CrossRef] [Green Version]
- McLachlan, D.R. Aluminium in Neurological and Neurodegenerative Disease. Mol. Neurobiol. 2019, 56, 1531–1538. [Google Scholar] [CrossRef]
- Cummings, J.; Aisen, P.; Apostolova, L.G.; Atri, A.; Salloway, S.; Weiner, M. Aducanumab: Appropriate use recommendations. J. Prev. Alzheimer’s Dis. 2021, 4, 398–410. [Google Scholar] [CrossRef] [PubMed]
- Raschetti, R.; Albanese, E.; Vanacore, N.; Maggini, M. Cholinesterase inhibitors in mild cognitive impairment: A systematic review of randomised trials. PLoS Med. 2007, 4, e338. [Google Scholar] [CrossRef] [PubMed]
- Colizzi, C. The protective effects of polyphenols on Alzheimer’s disease: A systematic review. Review Article. Alzheimer’s Dement. Transl. Res. Clin. Interv. 2019, 5, 184–196. [Google Scholar] [CrossRef]
- Ogunlade, B.; Adelakun, S.A.; Agie, J.A. Nutritional supplementation of gallic acid ameliorates Alzheimer-type hippocampal neurodegeneration and cognitive impairment induced by aluminum chloride exposure in adult Wistar rats. Drug Chem. Toxicol. 2020, 45, 651–662. [Google Scholar] [CrossRef] [PubMed]
- Nour, Y.; Majed, J.; Nawal, A.; Chantal, M. The Immunomodulatory and Anti-Inflammatory Role of Polyphenols. Nutrients 2018, 10, 1618. [Google Scholar]
- Montone, A.M.I.; Papaianni, M.; Malvano, F.; Capuano, F.; Capparelli, R.; Albanese, D. Lactoferrin, Quercetin, and Hydroxyapatite Act Synergistically against Pseudomonas fluorescens. Int. J. Mol. Sci. 2021, 22, 9247. [Google Scholar] [CrossRef]
- Awad, A.M.; Kumar, P.; Ismail-Fitry, M.R.; Jusoh, S.; Aziz, M.F.; Sazili, A.Q. Overview of plant extracts as natural preservatives in meat. J. Food Process. Preserv. 2022, 46, 8. [Google Scholar] [CrossRef]
- Jalilzadeh-Afshari, A.; Fadaei, V. Characterization of flavored milk containing bitter orange peel extract and Gaz-angubin. Food Sci. Nutr. 2020, 9, 164–171. [Google Scholar] [CrossRef]
- Shehata, M.G.; Awad, T.S.; Asker, D.; El Sohaimy, S.A.; Abd El- Aziz, N.M.; Youssef, M.M. Antioxidant and antimicrobial activities and UPLC-ESI-MS/MS polyphenolic profile of sweet orange peel extracts. Curr. Res. Nutr. Food Sci. 2021, 4, 326–335. [Google Scholar] [CrossRef]
- Rafiq, S. Citrus peel as a source of functional ingredient: A review. J. Saudi Soc. Agric. Sci. 2018, 17, 351–358. [Google Scholar] [CrossRef] [Green Version]
- Escobedo-Avellaneda, Z.; Gutiérrez-Uribe, J.; Valdez-Fragoso, A.; Torres, J.A.; Welti-Chanes, J. Phytochemicals and antioxidant activity of juice, flavedo, albedo and comminuted orange. J. Funct. Foods 2014, 6, 470–481. [Google Scholar] [CrossRef]
- Abd El-Aziz, N.M.; Awad, O.M.; Shehata, M.G.; El-Sohaimy, S.A. Antioxidant and anti-acetylcholinesterase potential of artichoke phenolic compounds. Food Biosci. 2021, 41, 101006. [Google Scholar] [CrossRef]
- Makris, D.P.; Sahin, S. Polyphenolic Antioxidants from Agri-Food Waste Biomass. Antioxidants 2019, 8, 624. [Google Scholar] [CrossRef] [Green Version]
- Daglia, M.; Di Lorenzo, A.; Nabavi, S.F.; Talas, Z.S.; Nabavi, S.M. Polyphenols: Well beyond the antioxidant capacity: Gallic acid and related compounds as neuroprotective agents: You are what you eat! Curr. Pharm. Biotechnol. 2014, 15, 362–372. [Google Scholar] [CrossRef] [PubMed]
- Na, J.Y.; Kim, S.; Song, K.; Kwon, J. Rutin alleviates prion Peptide-induced cell death through inhibiting apoptotic pathway activation in dopaminergic neuronal cells. Cell. Mol. Neurobiol. 2014, 34, 107. [Google Scholar] [CrossRef]
- August, P.M.; Gindri dos Santos, B. Chapter 20—Naringin and naringenin in neuroprotection and oxidative stress. In Oxidative Stress and Dietary Antioxidants in Neurological Diseases; Academic Press: Cambridge, MA, USA, 2020; pp. 309–323. [Google Scholar]
- Long, T. Ferulic Acid Exerts Neuroprotective Effects via Autophagy Induction in C. elegans and Cellular Models of Parkinson’s Disease. Oxidative Med. Cell. Longev. 2022, 2022, 3723567. [Google Scholar] [CrossRef]
- Benameur, T.; Soleti, R.; Porro, C. The Potential Neuroprotective Role of Free and Encapsulated Quercetin Mediated by miRNA against Neurological Diseases. Nutrients 2021, 13, 1318. [Google Scholar] [CrossRef] [PubMed]
- Jankobek, J.; Seruga, M.; Krivak, P. The influence of interactions among phenolic compounds on the antiradical activity of chokeberries (Aronia melanocarpa). Int. J. Food Sci. Nutr. 2011, 62, 345–352. [Google Scholar] [CrossRef]
- Altunkaya, A.; Becker, E.M.; Gokmen, V.; Skibsted, L.H. Antioxidant activity of lettuce extract (Lactuca sativa) and synergism with added phenolic antioxidants. Food Chem. 2009, 115, 163–168. [Google Scholar] [CrossRef]
- Ilyasov, I.R.; Beloborodov, V.L.; Selivanova, I.A.; Terekhov, R.P. ABTS/PP Decolorization Assay of Antioxidant Capacity Reaction Pathways. Int. J. Mol. Sci. 2020, 21, 1131. [Google Scholar] [CrossRef] [Green Version]
- Ismail, A.; Marjan, Z.M.; Foong, C. Total antioxidant activity and phenolic content in selected vegetables. Food Chem. 2004, 87, 581–586. [Google Scholar] [CrossRef]
- Hayyan, M.; Hashim, M.A.; Al Nashef, I.M. Superoxide ion: Generation and chemical implications. Chem. Rev. 2016, 116, 3029–3085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dizdaroglu, M.; Jaruga, P. Mechanisms of free radical-induced damage to DNA. Free Rad. Res. 2012, 46, 382–419. [Google Scholar] [CrossRef] [PubMed]
- Treml, J.; Šmejkal, K. Flavonoids as potent scavengers of hydroxyl radicals. Compr. Rev. Food Sci. Food Saf. 2016, 15, 720–738. [Google Scholar] [CrossRef]
- Lopa, S.S. Phytochemical Analysis and Cholinesterase Inhibitory and Antioxidant Activities of Enhydra fluctuans Relevant in the Management of Alzheimer’s Disease. Int. J. Food Sci. 2021, 2021, 8862025. [Google Scholar] [CrossRef]
- Naik, G.H. In vitro antioxidant studies and free radical reactions of triphala, an ayurvedic formulation and its constituents. Phytother. Res. 2005, 19, 582–586. [Google Scholar] [CrossRef]
- Tancheva, L. New mechanisms in preventive effect of ellagic acid on cognition in mice with Alzheimer’s disease type dementia. Bulg. Chem. Commun. 2017, 50, 20–24. [Google Scholar]
- Mohan, V.; Gibbs, A.C.; Cummings, M.D.; Jaeger, E.P.; DesJarlais, R.L. Docking: Successes and challenges. Curr. Pharm. Des. 2005, 11, 323–333. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.E.P. Amelioration of aluminium chloride (AlCl3) induced neurotoxicity by combination of Rivastigmine and Memantine with Artesunate in Albino wistar rats. Biomed. Pharmacol. J. 2019, 12, 703–711. [Google Scholar] [CrossRef]
- Capriello, T.; Grimaldi, M.C.; Cofone, R.; D’Aniello, S.; Ferrandino, I. Effects of aluminium and cadmium on hatching and swimming ability in developing zebrafish. Chemosphere 2019, 222, 243–249. [Google Scholar] [CrossRef]
- Inneh, C.; Eiya, B. Anticholinesterase activity and Antioxidant Effect of Vitamin E in Aluminum Chloride Induced Toxicity in Drosophila Melanogaster. FASEB J. 2022, 36. [Google Scholar] [CrossRef]
- Dey, M.; Singh, R.K. Neurotoxic effects of aluminium exposure as a potential risk factor for Alzheimer’s disease. Pharmacol. Rep. 2022, 74, 439–450. [Google Scholar] [CrossRef]
- Mori, T.; Koyama, N.; Yokoo, T.; Segawa, T.; Maeda, M.; Sawmiller, D.; Tan, J.; Town, T. Gallic acid is a dual α/β-secretase modulator that reverses cognitive impairment and remediates pathology in Alzheimer mice. J. Biol. Chem. 2020, 295, 16251–16266. [Google Scholar] [CrossRef] [PubMed]
- Ferk, F. Gallic acid improves health-associated biochemical parameters and prevents oxidative damage of DNA in type 2 diabetes patients: Results of a placebo-controlled pilot study. Mol. Nutr. Food Res. 2018, 62, 1700482. [Google Scholar] [CrossRef] [PubMed]
- Choubey, S. Medicinal importance of gallic acid and its ester derivatives: A patent review. Pharm. Pat. Anal. 2015, 4, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Hajipour, S. Effect of gallic acid on dementia type of Alzheimer’s disease in rats: Electrophysiological and histological studies. Basic Clin. Neurosci. 2016, 7, 97–106. [Google Scholar] [PubMed]
- Uddin, M.S. Revisiting the Amyloid Cascade Hypothesis: From Anti-A_ Therapeutics to Auspicious NewWays for Alzheimer’s Disease review. Int. J. Mol. Sci. 2020, 21, 5858. [Google Scholar] [CrossRef]
- Volloch, V.; Rits-Volloch, S. The Amyloid Cascade Hypothesis 2.0: On the Possibility of Once-in-a-Lifetime-Only Treatment for Prevention of Alzheimer’s Disease and for Its Potential Cure at Symptomatic Stages. J. Alzheimers Dis. Rep. 2022, 6, 369–399. [Google Scholar] [CrossRef]
- Karran, E.; De Strooper, B. The amyloid hypothesis in Alzheimer disease: New insights from new therapeutics. Nat. Rev. Drug Discov. 2022, 21, 306–318. [Google Scholar] [CrossRef]
- Walsh, D.M.; Selkoe, D.J. Oligomers on the brain: The emerging role of soluble protein aggregates in neurodegeneration. Protein Pept. Lett. 2004, 11, 213–228. [Google Scholar] [CrossRef]
- Williams, T.L.; Serpell, L.C. Membrane and surface inter.actions of Alzheimer’s Abeta peptide–insights into the mechanism of cytotoxicity. FEBS J. 2011, 278, 3905–3917. [Google Scholar] [CrossRef] [PubMed]
- Sakalauskas, A.; Ziaunys, M.; Smirnovas, V. Gallic acid oxidation products alter the formation pathway of insulin amyloid fibrils. Sci. Rep. 2020, 10, 14466. [Google Scholar] [CrossRef] [PubMed]
- Yu, M. Gallic acid disruption of Aβ1–42 aggregation rescues cognitive decline of APP/PS1 double transgenic mouse. Neurobiol. Dis. 2019, 124, 67–80. [Google Scholar] [CrossRef]
- Liu, Y. Gallic acid is the major component of grape seed extract that inhibits amyloid fibril formation. Bioorg. Med. Chem. Lett. 2013, 23, 6336–6340. [Google Scholar] [CrossRef] [PubMed]
- Ghofrani, S. Naringenin improves learning and memory in an Alzheimer’s disease rat model: Insights into the underlying mechanisms. Eur. J. Pharmacol. 2015, 764, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Ono, K.; Hirohata, M.; Yamada, M. Ferulic acid destabilizes preformed beta-amyloid fibrils in vitro. Biochem. Biophys. Res. Commun. 2005, 336, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Wang, N.Y. Ferulic Acid Ameliorates Alzheimer’s Disease-like Pathology and Repairs Cognitive Decline by Preventing Capillary Hypofunction in APP/PS1 Mice. Neurotherapeutics 2021, 18, 1064–1080. [Google Scholar] [CrossRef]
- Islam, M.S. Neuropharmacological Effects of Quercetin: A Literature-Based Review. Front Pharmacol. 2021, 12, 665031. [Google Scholar] [CrossRef]
- Zaplatic, E.; Bule, M.; Shah, S.Z.A.; Uddin, M.S.; Niaz, K. Molecular Mechanisms Underlying Protective Role of Quercetin in Attenuating Alzheimer’s Disease. Life Sci. 2019, 224, 109–119. [Google Scholar] [CrossRef]
- Rudajev, V.; Novotny, J. Cholesterol as a key player in amyloid b-mediated toxicity in Alzheimer’s disease. Front. Mol. Neurosci. 2022, 15, 937056. [Google Scholar] [CrossRef]
- Hirai, M. Preferential intercalation of human amyloid-b peptide into interbilayer region of lipid-raft membrane in macromolecular crowding environment. J. Phys. Chem. B 2018, 122, 9482–9489. [Google Scholar] [CrossRef] [PubMed]
- Yuan, C.Y.; Lee, Y.J.; Hsu, G.S.W. Aluminum overload increases oxidative stress in four functional brain areas of neonatal rats. J. Biomed. Sci. 2012, 19, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Salam, O.M.; El-Shamarka, M.E.; Youness, E.R.; Shaffie, N. Inhibition of aluminum chloride-induced amyloid Aβ peptide accumulation and brain neurodegeneration by Bougainvillea spectabilis flower decoction. Iran. J. Basic Med. Sci. 2021, 24, 1437–1445. [Google Scholar] [PubMed]
- Metodiewa, D.; Kośka, C. Reactive oxygen species and reactive nitrogen species: Relevance to cyto(neuro)toxic events and neurologic disorders. An overview. Neurotox. Res. 2000, 1, 197–233. [Google Scholar] [CrossRef]
- Persson, T.; Popescu, B.O.; Cedazo-Minguez, A. Oxidative stress in Alzheimer’s disease: Why did antioxidant therapy fail? Oxidative Med. Cell. Longev. 2014, 2014, 427318. [Google Scholar] [CrossRef]
- Kaizer, R.R.; Corrêa, M.C.; Spanevello, R.M.; Morsch, V.M.; Mazzanti, C.M.; Gonçalves, J.F.; Schetinger, M.R. Acetylcholinesterase activation and enhanced lipid peroxidation after long-term exposure to low levels of aluminum on di_erent mouse brain regions. J. Inorg. Biochem. 2005, 9, 1865–1870. [Google Scholar] [CrossRef]
- Scali, C.; Casamenti, F.; Bellucci, A.; Costagli, C.; Schmidt, B.; Pepeu, G. Effect of subchronic administration of metrifonate, rivastigmine and donepezil on brain acetylcholine in aged F344 rats. J. Neural Transm. 2002, 109, 1067–1080. [Google Scholar] [CrossRef]
- Marques-Coelho, D. Differential transcript usage unravels gene expression alterations in Alzheimer’s disease human brains. NPJ Aging Mech. Dis. 2021, 7, 2. [Google Scholar] [CrossRef]
- Nguyen, K.V. β-Amyloid precursor protein (APP) and the human diseases. AIMS Neurosci. 2019, 6, 273–281. [Google Scholar] [CrossRef]
- Olsson, F. Characterization of intermediate steps in amyloid beta (Abeta) production under near-native conditions. J. Biol. Chem. 2014, 289, 1540–1550. [Google Scholar] [CrossRef] [Green Version]
- Schneider, L.S.; Mangialasche, F.; Andreasen, N.; Feldman, H.; Giacobini, E.; Jones, R.; Mantua, V.; Mecocci, P.; Pani, L.; Winblad, B.; et al. Clinical trials and late-stage drug development for Alzheimer’s disease: An appraisal from 1984 to 2014. J. Intern. Med. 2014, 275, 251–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hampel, H.; Vassar, R.; De Strooper, B.; Hardy, J.; Willem, M.; Singh, N.; Zhou, J.; Yan, R.; Vanmechelen, E.; de Vos, A.; et al. The β-Secretase BACE1 in Alzheimer’s Disease. Biol. Psychiatry 2021, 89, 745–756. [Google Scholar] [CrossRef] [PubMed]
- Mullard, A. Alzheimer prevention hopes continue to dim. Nat. Rev. Drug Discov. 2020, 19, 226. [Google Scholar] [CrossRef]
- D’Arcy, M.S. Cell death: A review of the major forms of apoptosis, necrosis and autophagy. Cell Biol. Int. 2019, 43, 582–592. [Google Scholar] [CrossRef]
- Thenmozhi, A.J.; Raja, W.T.R.; Manivasagam, T.; Janakiraman, U.; Essa, M.M. Hesperidin ameliorates cognitive dysfunction, oxidative stress and apoptosis against aluminium chloride induced rat model of Alzheimer’s disease. Nutr. Neurosci. 2017, 20, 360–368. [Google Scholar] [CrossRef]
- Kabir, M.T. Exploring the Role of PSEN Mutations in the Pathogenesis of Alzheimer’s Disease. Neurotox. Res. 2020, 38, 833–849. [Google Scholar] [CrossRef] [PubMed]
- Elfiky, A.M.; Mahmoud, A.A.; Elreedy, H.A.; Ibrahim, K.S.; Ghazy, M.A. Quercetin stimulates the non-amyloidogenic pathway via activation of ADAM10 and ADAM17 gene expression in aluminum chloride-induced Alzheimer’s disease rat model. Life Sci. 2021, 285, 119964. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhao, F.; Ma, X.; Perry, G.; Zhu, X. Mitochondria dysfunction in the pathogenesis of Alzheimer’s disease: Recent advances. Mol. Neurodegener. 2020, 15, 30. [Google Scholar] [CrossRef]
- Elena, A.; Asier, R.; Maria, V.S.; Estibaliz, C.; Calos, M. Chapter 14—Polyphenols attenuate mitochondrial dysfunction induced by amyloid peptides. In Mitochondrial Physiology and Vegetal Molecules; Academic Press: Cambridge, MA, USA, 2021; pp. 317–337. [Google Scholar]
- Johnson, E.C.B. Large-scale proteomic analysis of Alzheimer’s disease brain and cerebrospinalfluid reveals early changes in energy metabolism associated with microglia and astrocyte activation. Nat. Med. 2020, 26, 769–780. [Google Scholar] [CrossRef]
- Palop, J.J.; Mucke, L. Amyloid-beta-induced neuronal dysfunction in Alzheimer’s disease: From synapses toward neural networks. Nat. Neurosci. 2010, 13, 812–818. [Google Scholar] [CrossRef] [Green Version]
- Rybka, V. Transmission Electron Microscopy Study of Mitochondria in Aging Brain Synapses. Antioxidants 2019, 11, 171. [Google Scholar] [CrossRef] [Green Version]
- Awad, O.M.E.; El-Sohaimy, S.A.; Ghareeb, D.A.; Aboulenein, A.; Saleh, S.R.; El-Aziz, N.M.A. Phytochemical Analysis and toxicity Assessment of Artichoke By-product Extract. Pak. J. Biol. Sci. 2020, 23, 81–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singleton, V.L.; Rossi, J.J. Colorimetry of total phenolics with phospho-molybdic phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Zhishen, J.; Mengcheng, T.; Jianming, W. The determination of flavonoid contents in mulberry and their scavenging effects on Superoxide radicals. Food Chem. 1999, 64, 555–559. [Google Scholar] [CrossRef]
- Tomaino, A.; Martorana, M.; Arcoraci, T. Antioxidant activity and phenolic profile of pistachio (Pistacia vera L., variety Bronte) seeds and skins. Biochimie 2010, 92, 1115–1122. [Google Scholar] [CrossRef]
- Bandoniene, D.; Murkovic, M.; Pfannhauser, W.; Venskutonis, P.R.; Gruzdiene, D. Detection and activity evaluation of radical scavenging compounds by using DPPH free radical and on-line HPLC–DPPH methods. Eur. Food Res. Technol. 2002, 214, 143–147. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Ozyürek, M.; Bektaşoğlu, B.; Güçlü, K.; Apak, R. Hydroxyl radical scavenging assay of phenolics and flavonoids with a modified cupric reducing antioxidant capacity (CUPRAC) method using catalase for hydrogen peroxide degradation. Anal. Chim. Acta 2008, 2, 196–206. [Google Scholar] [CrossRef]
- Cavdara, H.; Senturkb, M.; Guneyc, M.; Durdagid, S.; Kayikd, G.; Supuran, C.T.; Ekinci, D. Inhibition of acetylcholinesterase and butyrylcholinesterase with uracil derivatives: Kinetic and computational studies. J. Enzym. Inhib. Med. Chem. 2019, 34, 429–437. [Google Scholar] [CrossRef] [Green Version]
- Ali, A.A.; Ahmed, H.I.; Abu-Elfotuh, K. Modeling Stages Mimic Alzheimer’s Disease Induced by Different Doses of Aluminum in Rats: Focus on Progression of the Disease in Response to Time. J. Alzheimers Dis. Park. 2016, 1, 2. [Google Scholar]
- Kaur, R.; Parveen, S.; Mehan, S.; Khanna, D.; Kalra, S. Neuroprotective Effect of Ellagic Acid Against Chronically Scopolamine Induced Alzheimer’s Type Memory and Cognitive Dysfunctions: Possible Behavioural and Biochemical Evidences. Int. J. Prev. Med. 2015, 1, 45–64. [Google Scholar]
- Haug, K.H.; Myhrer, T.; Fonnum, F. The combination of donepezil and procyclidine protects against soman-induced seizures in rats. Toxicol. Appl. Pharmacol. 2007, 220, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantification of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Pejman, T.; Farzam, J.; Muhammad, S. Three-Dimensional Stochastic Characterization of Shale SEM Images. Transp. Porous Media 2015, 110, 521–531. [Google Scholar]
- Tappel, A.L.; Zalkin, H. Inhibition of lipid peroxidation in mitochondria by vitamin E. Arch. Biochem. Biophys. 1959, 80, 333. [Google Scholar] [CrossRef]
- Montgomery, H.A.C.; Dymock, J.F. The determination of nitrite in water. Analyst 1961, 86, 414–416. [Google Scholar]
- Ellman, G.L. Tissue sulfhydryl groups. Arch. Biochem. Biophys. 1959, 82, 70–77. [Google Scholar] [CrossRef]
- Paglia, E.; Valentine, N. Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. J. Lab. Med. 1967, 70, 158–169. [Google Scholar]
- Habig, W.H.; Pabst, M.J.; Jakoby, W.B. Glutathione S-transferases. The first enzymatic step in mercapturic acid formation. J. Biol. Chem. 1974, 249, 7130–7139. [Google Scholar] [CrossRef]
- Marklund, S.; Marklund, G. Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxidedismutase. Eur. J. Biochem. 1974, 47, 469–474. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Featherstone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Piotr, C.; Nicoletta, S. The single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction: Twenty-something years on. Nat. Protoc. 2006, 1, 581–585. [Google Scholar]
- Wang, W.; Barnaby, J.Y.; Tada, Y.; Li, H.; Tör, M.; Caldelari, D.; Lee, D.-U.; Fu, X.-D.; Dong, X. Timing of plant immune responses by a central circadian regulator. Nature 2011, 470, 110–114. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, L.A.; Feeney, B.C. A Simple Guide to IBM SPSS Statistics for Version 20.0; Wadsworth Publishing: Belmont, CA, USA, 2013. [Google Scholar]
- Kotz, S.; Balakrishnan, N.; Read, C.B.; Vidakovic, B. Encyclopedia of Statistical Sciences, 2nd ed.; Wiley-Interscience: Hoboken, NJ, USA, 2006; ISBN 978-0-471-15044-2-9686. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Results |
|---|---|
| 255.86 ± 1.77 |
| 52.06 ± 1.74 |
| |
| DPPH | 67.90 ± 1.05 |
| ABTS | 60.48 ± 0.91 |
| Hydroxyl radical scavenging activity | 63.70 ± 0.30 |
| |
| OPE | 0.87 ± 0.025 b |
| Gallic acid | 2.45± 0.001 c |
| Compounds | Chemical Name | Formula | R.T. | Conc. (μg/100 g) |
|---|---|---|---|---|
| Gallic acid | 3,4,5-trihydroxybenzoic acid | C7H6O5 | 3.13 | 3388.60 |
| Chlorogenic acid | (1S,3R,4R,5R)-3-[(E)-3-(3,4-dihydroxyphenyl)prop-2-enoyl]oxy-1,4,5-trihydroxycyclohexane-1-carboxylic acid | C16H18O9 | 3.49 | 874.25 |
| Catechin | (2R,3S)-2-(3,4-dihydroxyphenyl)-3,4-dihydro-2H-chromene-3,5,7-triol | C15H14O6 | 3.84 | ND |
| Caffeine | 1,3,7-trimethylpurine-2,6-dione, guaranine, methyltheobromine | C8H10N4O2 | 4.03 | ND |
| Coffeic acid | (2E)-3-(3,4-dihydroxyphenyl) prop-2-enoic acid, 3,4-dihydroxy-trans-cinnamate | C9H8O4 | 4.97 | ND |
| Syringic acid | 4-hydroxy-3,5 dimethoxy benzoic acid, gallic acid 3,5-dimethyl ether | C9H10O5 | 5.37 | 330.91 |
| Rutin | 2-(3,4-dihydroxyphenyl)-5,7-dihydroxy-3-[(2S,3R,4S,5S,6R)-3,4,5-trihydroxy-6-[[(2R,3R,4R,5R,6S)-3,4,5-trihydroxy-6- methyloxan-2-yl] oxymethyl] oxan-2-yl] oxychromen-4-one | C27H30O16 | 5.70 | 1952.64 |
| Pyro catechol | 3-(methylaminomethyl)benzene-1,2-diol;hydrochloride | C8H12ClNO2 | 5.82 | 0.00 |
| Ellagic acid | 3,7,8-tetrahydroxy-[1]-benzopyrano[5,4,3,-cde]-[1]-benzopyran-5,10-dione | C14H6O8 | 6.83 | 336.16 |
| O-coumaric acid | (E)-3-(2-hydroxy phenyl) prop-2-enoic acid, 2-hydroxycinnamic acid | C9H8O3 | 7.72 | 183.16 |
| Vanillin | 4-hydroxy-3-methoxybenzaldehyde vanillic aldehyde | C8H8O3 | 8.38 | ND |
| Ferulic acid | (2E)-3-(4-hydroxy-3-methoxyphenyl) prop-2-enoic acid, 3-methoxy-4-hydroxycinnamic acid | C10H10O4 | 8.80 | 1335.96 |
| Naringenin | 5,7-dihydroxy-2-(4-hydroxyphenyl)-2,3-dihydrochromen-4-one | C15H12O5 | 9.43 | 1967.43 |
| Propyl gallate | propyl 3,4,5-trihydroxybenzoate | C10H12O5 | 10.22 | 1212.96 |
| 4′,7-Dihydroxy isoFlavone | 8-[(2S,3R,4S,5S,6R)-4,5-dihydroxy-6-(hydroxymethyl)-3-[(2S,3R,4S,5R)-3,4,5-trihydroxyoxan-2-yl]oxyoxan-2-yl]-7-hydroxy-3-(4-hydroxyphenyl)chromen-4-one | C26H28O13 | 10.49 | 277.49 |
| Querectin | 2-(3,4-dihydroxyphenyl)-3,5,7-trihydroxychromen-4-one;dihydrate | C₁₅H₁₀O₇ | 10.69 | 1266.97 |
| Cinnamic acid | (2E)-3-phenylprop-2-enoic acid | C9H8O2 | 11.22 | 47.08 |
| Ligand Name | Molecular Formula | Molecular Weight (g/mol) | Binding Energy (ΔG, Kcal/mol) | Binding Sites |
|---|---|---|---|---|
| Gallic acid | C7H6O5 | 170.12 | −3.72 | GLU 202, SER 203, GLY 121, GLY 122 |
| Acetylcholine | C7H16NO2 | 146.21 | −5.69 | SER 203, GLY 121, GLY 122, ALA 204, ARG 296, and PHE 295 |
| Aricept | C24H29NO3 | 379.5 | −9.47 | TYR 465, TYR 124, HIS 447, and SER 203 |
| Exp. Groups | Control | Induction | Aricept Protective Group | Gallic Protective Group | OPE 100 Protective Group | OPE 200 Protective Group |
|---|---|---|---|---|---|---|
| Body weight gain | 54.56 ± 2.05 a | 34.14 ± 2.05 c | 39.47 ± 1.13 b | 34.66 ± 1.28 c | 34.65 ± 1.88 c | 40.00 ± 1.80 b |
| Brain structure | ||||||
| Aβ42 (Pg/g tissue) | 5.32 ± 0.98 f | 19.18 ± 1.15 a | 7.97 ± 0.63 e | 13.86 ± 1.70 c | 16.73 ± 0.97 b | 11.55 ± 1.02 d |
| Total cholesterol (mg/mg protein) | 46.11 ± 0.93 e | 94.88 ± 0.83 a | 70.65 ± 1.72 b | 51.43 ± 1.46 d | 58.83 ± 1.04 c | 44.45 ± 0.90 e |
| Phospholipids levels (mg/mg protein) | 72.33 ± 1.19 a | 33.50 ± 0.86 e | 60.73 ± 1.55 c | 38.20 ± 1.25 d | 67.53 ± 1.50 b | 71.30 ± 1.41 a |
| Total cholesterol/ phospholipids | 0.63 ± 0.00 e | 2.83 ± 0.07 a | 1.16 ± 0.02 c | 1.34 ± 0.08 b | 0.87 ± 0.03 d | 0.62 ± 0.005 e |
| Parameters of oxidative stress | ||||||
| TBARS (umol/g × 10−3) | 1.32 ± 0.15 d | 5.43 ± 0.83 a | 3.67 ± 0.59 b | 2.41 ± 0.21 c | 4.25 ± 0.38 b | 2.34 ± 0.22 c |
| NO level (µm/mg protein) | 12.43 ± 0.96 c | 32.76 ± 1.47 a | 23.35 ± 1.31 b | 23.54 ± 0.73 b | 22.58 ± 0.79 b | 21.86 ± 1.68 b |
| GSH level (mg/mg protein × 10−2) | 0.43 ± 0.03 a | 0.24 ± 0.12 b | 0.60 ± 0.10 a | 0.58 ± 0.09 a | 0.55 ± 0.04 a | 0.62 ± 0.16 a |
| Groups | Control | Induction | Aricept | Gallic Acid | OPE 200 |
|---|---|---|---|---|---|
| Mit. Length (nm) | 435.67 ± 1.24 b | 186.30 ± 0.65 e | 368.27 ± 0.75 d | 407 ± 1.00 c | 450.33 ± 0.35 a |
| Mit. Width (nm) | 261 ± 1.00 b | 157 ± 2 d | 248.87 ± 0.90 c | 263.66 ± 2.54 b | 300.78 ± 1.09 a |
| Primer Name | Primer Sequence from 5′–3′ | Anealing Temp. °C |
|---|---|---|
| Acetylcholinesterase (AChE) | R-CCACCGATCCTCTGGACGAG F-CGCTCCTGCTTGCTATAGTG | 60 |
| B-cell lymphoma-2 (BCL-2) | F-ATGTGTGTGGAGAGCGTCAACC R-TGAGCAGAGTCTTCAGAGACAGCC | 63 |
| Presenilin-2 (PSEN-2) | F-GAGCAGAGCCAA ATCAAA GG R-GGGAGA AAGAACAGCTCGTG | 60 |
| B–site APP-cleaving enzyme (BACE-1) | F-CGGGAGTGGTATTATGAAGTG R-AGGATGGTGATGCGGAAG | 60 |
| Amyloid precursor protein (APP) | F-AGAGGTCTACCCTGAACTGC R-ATCGCT TACAAACTCACCAAC | 54.9 |
| Glycerol-3-phosphate dehydrogenase (GPDH) | F-ATTGACCACTACCTGGGCAA R-GAGATACACTTCAACACTTTGACCT | 60–65 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abd El-Aziz, N.M.; Shehata, M.G.; Alsulami, T.; Badr, A.N.; Elbakatoshy, M.R.; Ali, H.S.; El-Sohaimy, S.A. Characterization of Orange Peel Extract and Its Potential Protective Effect against Aluminum Chloride-Induced Alzheimer’s Disease. Pharmaceuticals 2023, 16, 12. https://doi.org/10.3390/ph16010012
Abd El-Aziz NM, Shehata MG, Alsulami T, Badr AN, Elbakatoshy MR, Ali HS, El-Sohaimy SA. Characterization of Orange Peel Extract and Its Potential Protective Effect against Aluminum Chloride-Induced Alzheimer’s Disease. Pharmaceuticals. 2023; 16(1):12. https://doi.org/10.3390/ph16010012
Chicago/Turabian StyleAbd El-Aziz, Nourhan Mohammad, Mohamed Gamal Shehata, Tawfiq Alsulami, Ahmed Noah Badr, Marwa Ramadan Elbakatoshy, Hatem Salama Ali, and Sobhy Ahmed El-Sohaimy. 2023. "Characterization of Orange Peel Extract and Its Potential Protective Effect against Aluminum Chloride-Induced Alzheimer’s Disease" Pharmaceuticals 16, no. 1: 12. https://doi.org/10.3390/ph16010012
APA StyleAbd El-Aziz, N. M., Shehata, M. G., Alsulami, T., Badr, A. N., Elbakatoshy, M. R., Ali, H. S., & El-Sohaimy, S. A. (2023). Characterization of Orange Peel Extract and Its Potential Protective Effect against Aluminum Chloride-Induced Alzheimer’s Disease. Pharmaceuticals, 16(1), 12. https://doi.org/10.3390/ph16010012