

Advances in Anti-Diabetic Cognitive Dysfunction Effect of Erigeron Breviscapus (Vaniot) Hand-Mazz

Abstract
:1. Introduction
2. Active Ingredients and Pharmacological Effects of EBHM
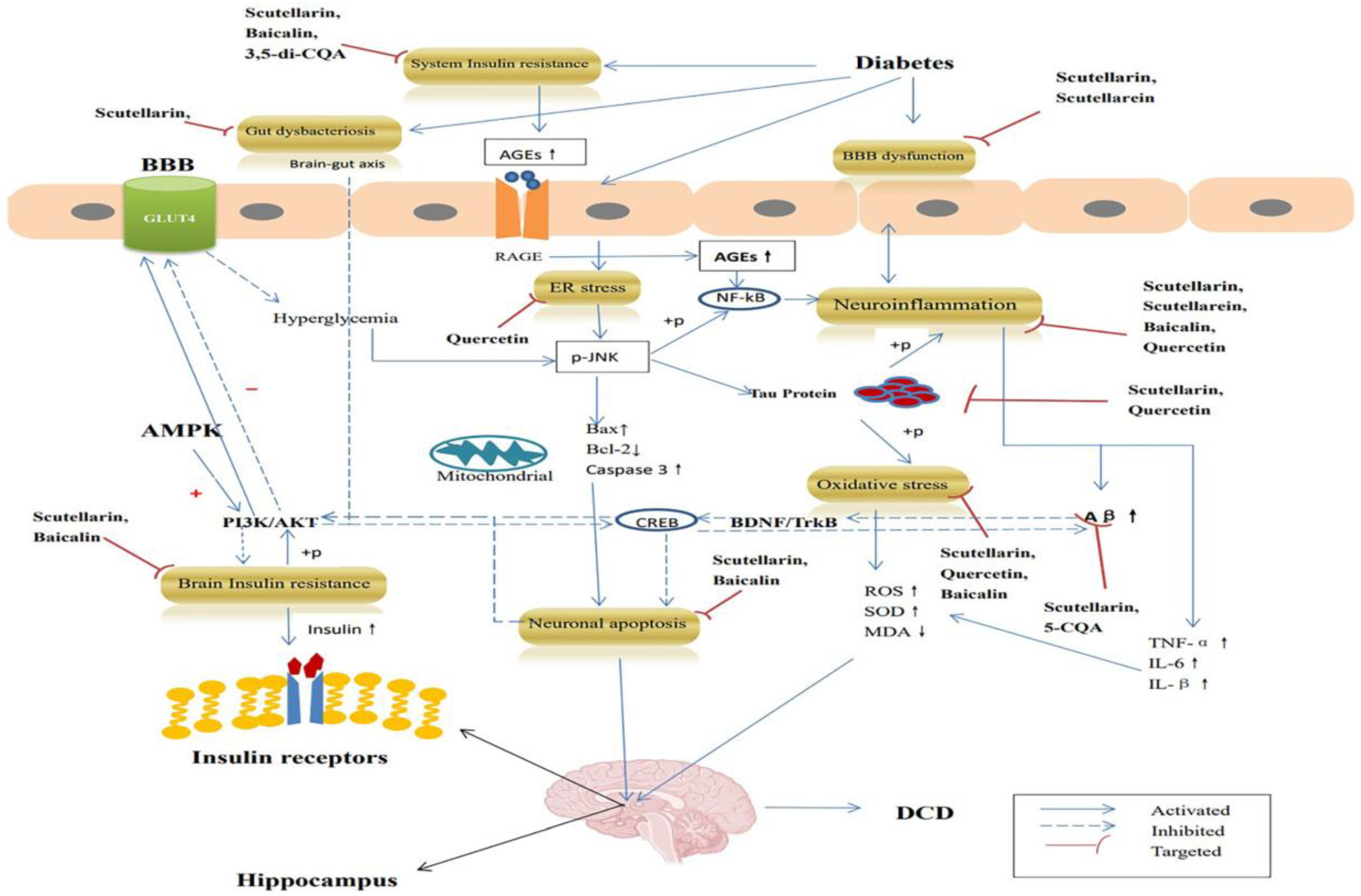
3. Anti-DCD Effects of EBHM
3.1. Targeted Insulin Resistance
3.2. Targeted Amyloid β-Peptide
3.3. Targeted Tau Protein
3.4. Targeted Neuroinflammation
3.5. Targeted Oxidative Stress
3.6. Targeted Endoplasmic Reticulum Stress
3.7. Targeted Anti-Apoptosis
3.8. Targeted Blood–Brain Barrier Injury
3.9. Targeted Gut Dysbacteriosis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- De la Monte, S.M.; Wands, J.R. Alzheimer’s Disease is Type 3 Diabetes—Evidence Reviewed. J. Diabetes Sci. Technol. 2008, 2, 1101–1113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biessels, G.J.; Despa, F. Cognitive decline and dementia in diabetes mellitus: Mechanisms and clinical implications. Nat. Rev. Endocrinol. 2018, 14, 591–604. [Google Scholar] [CrossRef] [PubMed]
- Giorda, C.B.; Roberto, G.; Barbara, T.; Roberta, M.; Alessandro, M.; Lucia, F.; Silvia, F.; Ivana, R. Increased incidence of type 1 diabetes in 2 years of COVID-19 pandemic. Acta Diabetol. 2022, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Biessels, G.J.; Staekenborg, S.; Brunner, E.; Brayne, C.; Scheltens, P. Risk of dementia in diabetes mellitus: A systematic review. Lancet Neurol. 2006, 5, 64–74. [Google Scholar] [CrossRef] [PubMed]
- Ennis, G.E.; Saelzler, U.; Umpierrez, G.E.; Moffat, S.D. Prediabetes and working memory in older adults. Brain Neurosci. Adv. 2020, 4, 2398212820961725. [Google Scholar] [CrossRef]
- Willmann, C.; Brockmann, K.; Wagner, R.; Kullmann, S.; Preissl, H.; Schnauder, G.; Maetzler, W.; Gasser, T.; Berg, D.; Eschweiler, G.W.; et al. Insulin sensitivity predicts cognitive decline in individuals with prediabetes. BMJ Open Diabetes Res. Care 2020, 8, e001741. [Google Scholar] [CrossRef]
- Xue, M.; Xu, W.; Ou, Y.-N.; Cao, X.-P.; Tan, M.-S.; Tan, L.; Yu, J.-T. Diabetes mellitus and risks of cognitive impairment and dementia: A systematic review and meta-analysis of 144 prospective studies. Ageing Res. Rev. 2019, 55, 100944. [Google Scholar] [CrossRef]
- You, Y.; Liu, Z.; Chen, Y.; Xu, Y.; Qin, J.; Guo, S.; Huang, J.; Tao, J. The prevalence of mild cognitive impairment in type 2 diabetes mellitus patients: A systematic review and meta-analysis. Acta Diabetol. 2021, 58, 671–685. [Google Scholar] [CrossRef]
- Ehtewish, H.; Arredouani, A.; El-Agnaf, O. Diagnostic, Prognostic, and Mechanistic Biomarkers of Diabetes Mellitus-Associated Cognitive Decline. Int. J. Mol. Sci. 2022, 23, 6144. [Google Scholar] [CrossRef]
- Luo, A.; Xie, Z.; Wang, Y.; Wang, X.; Li, S.; Yan, J.; Zhan, G.; Zhou, Z.; Zhao, Y.; Li, S. Type 2 diabetes mellitus-associated cognitive dysfunction: Advances in potential mechanisms and therapies. Neurosci. Biobehav. Rev. 2022, 137, 104642. [Google Scholar] [CrossRef]
- Wang, K.M.; Jiang, Z.; Yu, X.; Shao, Y.; Liu, H.M.; Wu, S.M.; Kong, L.M.; Wang, Z. Comparative efficacy and safety of traditional Chinese patent medicine for cognitive dysfunction in diabetic cognitive dysfunction. Medicine 2022, 101, e28946. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.D.; Sun, G.B.; Wu, Y.M.; Ye, T.Y.; Meng, X.B.; Sun, X.B. Pathogenesis of diabetic cognitive dysfunction and intervention by traditional Chinese medicine. Chin. Pharmacol. Bull. 2018, 34, 303–308. [Google Scholar]
- Madhavadas, S.; Subramanian, S. Cognition enhancing effect of the aqueous extract of Cinnamomum zeylanicum on non-transgenic Alzheimer’s disease rat model: Biochemical, histological, and behavioural studies. Nutr. Neurosci. 2016, 20, 526–537. [Google Scholar] [CrossRef]
- Rahman, M.A.; Hossain, S.; Abdullah, N.; Aminudin, N. Lingzhi or Reishi Medicinal Mushroom, Ganoderma lucidum (Agaricomycetes), Ameliorates Nonspatial Learning and Memory Deficits in Rats with Hypercholesterolemia and Alzheimer’s Disease. Int. J. Med. Mushrooms 2020, 22, 1067–1078. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Park, S.; Kang, J.; Kim, J.; Yoo, S.; Kim, D.-O.; Kim, G.-H.; Heo, H. Mixture of Phlorotannin and Fucoidan from Ecklonia cava Prevents the Aβ-Induced Cognitive Decline with Mitochondrial and Cholinergic Activation. Mar. Drugs 2021, 19, 434. [Google Scholar] [CrossRef]
- Fan, H.; Lin, P.; Kang, Q.; Zhao, Z.-L.; Wang, J.; Cheng, J.-Y. Metabolism and Pharmacological Mechanisms of Active Ingredients in Erigeron breviscapus. Curr. Drug Metab. 2021, 22, 24–39. [Google Scholar] [CrossRef]
- Wu, R.; Liang, Y.; Xu, M.; Fu, K.; Zhang, Y.; Wu, L.; Wang, Z. Advances in Chemical Constituents, Clinical Applications, Pharmacology, Pharmacokinetics and Toxicology of Erigeron breviscapus. Front. Pharmacol. 2021, 12, 656335. [Google Scholar] [CrossRef]
- Dong, X.; Qu, S. Erigeron breviscapus (Vant.) Hand-Mazz.: A Promising Natural Neuroprotective Agent for Alzheimer’s Disease. Front. Pharmacol. 2022, 13, 877872. [Google Scholar] [CrossRef]
- Zeng, X.Y.; Chen, S.H.; Shi, Z.T.; Tang, B.X.; Liu, Y.W. Study on influence factors of extraction technology of Erigeron breviscapus. Hubei J. Tradit. Chin. Med. 2016, 38, 63–65. [Google Scholar]
- Liu, M.Q.; Wei, D.F.; Liu, Z.Q.; Zhang, Z.J. Research Progress of Scutellarin’S Effects on Cognitive Impairment and Its Mechanism. J. Basic Chin. Med. 2019, 25, 139–142. [Google Scholar]
- Zhu, Y.N.; Yang, Q.M.; Zhang, S.; Zhang, M.; Gao, X.L. Advances in Neuroprotective Mechanisms of Baicalin and Baicalein. Lishizhen Med. Mater. Med. Res. 2020, 31, 921–925. [Google Scholar]
- Ma, N.; Li, Y.J.; Fan, J.P. Research Progress on Pharmacological Action of Quercetin. J. Liaoning Univ. Tradit. Chin. Med. 2018, 20, 221–224. [Google Scholar] [CrossRef]
- Tong, J.; Ma, B.; Ge, L.; Mo, Q.; Zhou, G.; He, J.; Wang, Y. Dicaffeoylquinic Acid-Enriched Fraction of Cichorium glandulosum Seeds Attenuates Experimental Type 1 Diabetes via Multipathway Protection. J. Agric. Food Chem. 2015, 63, 10791–10802. [Google Scholar] [CrossRef] [PubMed]
- Ishida, K.; Misawa, K.; Nishimura, H.; Hirata, T.; Yamamoto, M.; Ota, N. 5-Caffeoylquinic Acid Ameliorates Cognitive Decline and Reduces Aβ Deposition by Modulating Aβ Clearance Pathways in APP/PS2 Transgenic Mice. Nutrients 2020, 12, 494. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Lv, Y.L.; Wei, Y.L.; Zhang, Y.P.; Li, J.; Dou, Z.J. Research progress on mechanism of diabetic cognitive dysfunction. J. Chengde Med. Univ. 2022, 39, 155–159. [Google Scholar] [CrossRef]
- Xiu, J.Y.; Jiang, T.; Xu, K.; Wu, Y.Q. Molecular Mechanism and Treatment Strategy for Diabetes-Associated Cognitive Decline. Chin. J. Cell Biol. 2020, 42, 1884–1893. [Google Scholar]
- Zhang, S.; Wei, D.; Lv, S.; Wang, L.; An, H.; Shao, W.; Wang, Y.; Huang, Y.; Peng, D.; Zhang, Z. Scutellarin Modulates the Microbiota-Gut-Brain Axis and Improves Cognitive Impairment in APP/PS1 Mice. J. Alzheimer’s Dis. 2022, 89, 955–975. [Google Scholar] [CrossRef]
- Liu, G.; Liang, Y.; Xu, M.; Sun, M.; Sun, W.; Zhou, Y.; Huang, X.; Song, W.; Wang, Z. Protective mechanism of Erigeron breviscapus injection on blood–brain barrier injury induced by cerebral ischemia in rats. Sci. Rep. 2021, 11, 18451. [Google Scholar] [CrossRef]
- Wang, M.N.; Mei, Q.Y.; Liu, B.; Zhang, T.Y.; Zhang, Y.; Lu, B.; Sheng, Y.C. Scutellarein alleviates dysfunction of blood-brain-barrier initiated by hyperglycemia-stimulated microglia cells. Chin. Pharmacol. Bull. 2020, 36, 1542–1547. [Google Scholar]
- Gao, L.; Tang, H.; Zeng, Q.; Tang, T.; Chen, M.; Pu, P. The anti-insulin resistance effect of scutellarin may be related to antioxidant stress and AMPKα activation in diabetic mice. Obes. Res. Clin. Pract. 2020, 14, 368–374. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, J.; Wei, D.; An, H.; Liu, W.; Lai, Y.; Yang, T.; Shao, W.; Huang, Y.; Wang, L.; et al. Dengzhan Shengmai capsules and their active component scutellarin prevent cognitive decline in APP/PS1 mice by accelerating Aβ aggregation and reducing oligomers formation. Biomed. Pharmacother. 2019, 121, 109682. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, J.; Zhang, X.; Wang, L.; Hao, T.; Cheng, Y.; Wang, D. Scutellarin Exerts Hypoglycemic and Renal Protective Effects in db/db Mice via the Nrf2/HO-1 Signaling Pathway. Oxidative Med. Cell. Longev. 2019, 2019, 1354345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, E.N. Inhibitory Effect and Its Mechanism of Scutellarin on Apoptosis of Hippocampal Neuronal and Cognitive Dysfunction in Type 2 Diabetes Mellitus. Master’s Thesis, Kunming Medical University, Kunming, China, 2021. [Google Scholar] [CrossRef]
- Zhou, Y.; Ji, H.; Lin, B.Q.; Jiang, Y.; Li, P. The effects of five alkaloids from Bulbus Fritillariae on the concentration of cAMP in HEK cells transfected with muscarinic M2 receptor plasmid. Am. J. Chin. Med. 2006, 34, 901–910. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Teng, S.; He, J.; Sun, X.; Du, M.; Kou, L.; Wang, X. Pharmacological basis for application of scutellarin in Alzheimer’s disease: Antioxidation and antiapoptosis. Mol. Med. Rep. 2018, 18, 4289–4296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, Y.-Q.; Cui, Y.-B.; Gu, J.-H.; Liang, C.; Zhou, X.-F. Scutellarin Mitigates Aβ-Induced Neurotoxicity and Improves Behavior Impairments in AD Mice. Molecules 2018, 23, 869. [Google Scholar] [CrossRef] [PubMed]
- Baluchnejadmojarad, T.; Zeinali, H.; Roghani, M. Scutellarin alleviates lipopolysaccharide-induced cognitive deficits in the rat: Insights into underlying mechanisms. Int. Immunopharmacol. 2018, 54, 311–319. [Google Scholar] [CrossRef]
- Song, Z.; He, C.; Yu, W.; Yang, M.; Li, Z.; Li, P.; Zhu, X.; Xiao, C.; Cheng, S. Baicalin Attenuated Aβ1-42-Induced Apoptosis in SH-SY5Y Cells by Inhibiting the Ras-ERK Signaling Pathway. BioMed. Res. Int. 2022, 2022, 9491755. [Google Scholar] [CrossRef]
- Li, C.; Sui, C.; Wang, W.; Yan, J.; Deng, N.; Du, X.; Cheng, F.; Ma, X.; Wang, X.; Wang, Q. Baicalin Attenuates Oxygen–Glucose Deprivation/Reoxygenation–Induced Injury by Modulating the BDNF-TrkB/PI3K/Akt and MAPK/Erk1/2 Signaling Axes in Neuron–Astrocyte Cocultures. Front. Pharmacol. 2021, 12, 599543. [Google Scholar] [CrossRef]
- Ma, L.; Wu, F.; Shao, Q.; Chen, G.; Xu, L.; Lu, F. Baicalin Alleviates Oxidative Stress and Inflammation in Diabetic Nephropathy via Nrf2 and MAPK Signaling Pathway. Drug Des. Dev. Ther. 2021, 15, 3207–3221. [Google Scholar] [CrossRef]
- Noh, J.-W.; Kwon, O.-J.; Lee, B.-C. The Immunomodulating Effect of Baicalin on Inflammation and Insulin Resistance in High-Fat-Diet-Induced Obese Mice. Evidence-Based Complement. Altern. Med. 2021, 2021, 5531367. [Google Scholar] [CrossRef]
- Jin, X.; Liu, M.-Y.; Zhang, D.-F.; Zhong, X.; Du, K.; Qian, P.; Yao, W.-F.; Gao, H.; Wei, M.-J. Baicalin mitigates cognitive impairment and protects neurons from microglia-mediated neuroinflammation via suppressing NLRP3 inflammasomes and TLR4/NF-κB signaling pathway. CNS Neurosci. Ther. 2019, 25, 575–590. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Li, X.; Gao, P.; Tu, Y.; Zhao, M.; Li, J.; Zhang, S.; Liang, H. Baicalin attenuates Alzheimer-like pathological changes and memory deficits induced by amyloid β1–42 protein. Metab. Brain Dis. 2014, 30, 537–544. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.F.; Peng, H.L. Effect of baicalin on serum glucose, cholesterol and activation of AMPK in streptozotocin-induced type 2 diabetic rats. Chin. J. Pharmacol. Toxicol. 2011, 25, 145–150. [Google Scholar]
- Hu, T.; Shi, J.-J.; Fang, J.; Wang, Q.; Chen, Y.-B.; Zhang, S.-J. Quercetin ameliorates diabetic encephalopathy through SIRT1/ER stress pathway in db/db mice. Aging 2020, 12, 7015–7029. [Google Scholar] [CrossRef]
- Tang, L.; Li, K.; Zhang, Y.; Li, H.; Li, A.; Xu, Y.; Wei, B. Quercetin liposomes ameliorate streptozotocin-induced diabetic nephropathy in diabetic rats. Sci. Rep. 2020, 10, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Hu, T.; Lu, X.; Shi, J.; Liu, X.; Chen, Q.; Wang, Q.; Chen, Y.; Zhang, S. Quercetin protects against diabetic encephalopathy via SIRT1/NLRP3 pathway in db/db mice. J. Cell Mol. Med. 2020, 24, 3449–3459. [Google Scholar] [CrossRef]
- Park, D.-J.; Kang, J.-B.; Shah, M.-A.; Koh, P.-O. Quercetin alleviates the injury-induced decrease of protein phosphatase 2A subunit B in cerebral ischemic animal model and glutamate-exposed HT22 cells. J. Veter Med. Sci. 2019, 81, 1047–1054. [Google Scholar] [CrossRef] [Green Version]
- Shen, X.; Luo, T.; Li, S.; Ting, O.; He, F.; Xu, J.; Wang, H. Quercetin inhibits okadaic acid-induced tau protein hyperphosphorylation through the Ca2+-calpain-p25-CDK5 pathway in HT22 cells. Int. J. Mol. Med. 2017, 41, 1138–1146. [Google Scholar] [CrossRef] [Green Version]
- Zhu, X.; Cheng, Y.-Q.; Lu, Q.; Du, L.; Yin, X.-X.; Liu, Y.-W. Enhancement of glyoxalase 1, a polyfunctional defense enzyme, by quercetin in the brain in streptozotocin-induced diabetic rats. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2018, 391, 1237–1245. [Google Scholar] [CrossRef]
- Ramos-Rodriguez, J.J.; Jimenez-Palomares, M.; Murillo-Carretero, M.I.; Infante-Garcia, C.; Berrocoso, E.; Hernandez-Pacho, F.; Lechuga-Sancho, A.M.; Cozar-Castellano, I.; Garcia-Alloza, M. Central vascular disease and exacerbated pathology in a mixed model of type 2 diabetes and Alzheimer’s disease. Psychoneuroendocrinology 2015, 62, 69–79. [Google Scholar] [CrossRef]
- Wang, J.-Q.; Yin, J.; Song, Y.-F.; Zhang, L.; Ren, Y.-X.; Wang, D.-G.; Gao, L.-P.; Jing, Y.-H. Brain Aging and AD-Like Pathology in Streptozotocin-Induced Diabetic Rats. J. Diabetes Res. 2014, 2014, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benedict, C.; Grillo, C. Insulin Resistance as a Therapeutic Target in the Treatment of Alzheimer’s Disease: A State-of-the-Art Review. Front. Neurosci. 2018, 12, 215. [Google Scholar] [CrossRef]
- Leto, D.; Saltiel, A.R. Regulation of glucose transport by insulin: Traffic control of GLUT4. Nat. Rev. Mol. Cell Biol. 2012, 13, 383–396. [Google Scholar] [CrossRef] [PubMed]
- Soumaya, K. Molecular Mechanisms of Insulin Resistance in Diabetes. Diabetes 2012, 771, 240–251. [Google Scholar] [CrossRef]
- Hardie, D.G.; Ross, F.A.; Hawley, S.A. AMPK: A nutrient and energy sensor that maintains energy homeostasis. Nat. Rev. Mol. Cell Biol. 2012, 13, 251–262. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Hu, X.; Selvakumar, P.; Russell, R.R.; Cushman, S.W.; Holman, G.D.; Young, L.H. Role of the nitric oxide pathway in AMPK-mediated glucose uptake and GLUT4 translocation in heart muscle. Am. J. Physiol. Metab. 2004, 287, E834–E841. [Google Scholar] [CrossRef] [PubMed]
- Mihaylova, M.M.; Shaw, R.J. The AMPK signalling pathway coordinates cell growth, autophagy and metabolism. Nature 2011, 13, 1016–1023. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Jiang, H.; Cao, S.; Chen, Q.; Cui, M.; Wang, Z.; Li, D.; Zhou, J.; Qiu, F.; Kang, N. Baicalin and its metabolites suppresses gluconeogenesis through activation of AMPK or AKT in insulin resistant HepG-2 cells. Eur. J. Med. Chem. 2017, 141, 92–100. [Google Scholar] [CrossRef]
- Pivovarova, O.; Höhn, A.; Grune, T.; Pfeiffer, A.F.; Rudovich, N. Insulin-degrading enzyme: New therapeutic target for diabetes and Alzheimer’s disease? Ann. Med. 2016, 48, 614–624. [Google Scholar] [CrossRef]
- Li, H.; Wu, J.; Zhu, L.; Sha, L.; Yang, S.; Wei, J.; Ji, L.; Tang, X.; Mao, K.; Cao, L.; et al. Insulin degrading enzyme contributes to the pathology in a mixed model of Type 2 diabetes and Alzheimer’s disease: Possible mechanisms of IDE in T2D and AD. Biosci. Rep. 2018, 38, BSR20170862. [Google Scholar] [CrossRef] [Green Version]
- Lane, C.A.; Hardy, J.; Schott, J.M. Alzheimer’s disease. Eur. J. Neurol. 2017, 25, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Panza, F.; Lozupone, M.; Logroscino, G.; Imbimbo, B.P. A critical appraisal of amyloid-β-targeting therapies for Alzheimer disease. Nat. Rev. Neurol. 2019, 15, 73–88. [Google Scholar] [CrossRef]
- Selkoe, D.J.; Hardy, J. The amyloid hypothesis of Alzheimer’s disease at 25 years. EMBO Mol. Med. 2016, 8, 595–608. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Atluri, V.; Kaushik, A.; Yndart, A.; Nair, M. Alzheimer’s disease: Pathogenesis, diagnostics, and therapeutics. Int. J. Nanomed. 2019, 14, 5541–5554. [Google Scholar] [CrossRef] [Green Version]
- Stancu, I.-C.; Ferraiolo, M.; Terwel, D.; Dewachter, I. Tau Interacting Proteins: Gaining Insight into the Roles of Tau in Health and Disease. Tau Biol. 2019, 1184, 145–166. [Google Scholar] [CrossRef]
- El Khoury, N.B.; Gratuze, M.; Papon, M.-A.; Bretteville, A.; Planel, E. Insulin dysfunction and Tau pathology. Front. Cell. Neurosci. 2014, 8, 22. [Google Scholar] [CrossRef]
- Van der Harg, J.M.; Eggels, L.; Bangel, F.N.; Ruigrok, S.R.; Zwart, R.; Hoozemans, J.J.; la Fleur, S.E.; Scheper, W. Insulin deficiency results in reversible protein kinase A activation and tau phosphorylation. Neurobiol. Dis. 2017, 103, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Congdon, E.E.; Sigurdsson, E.M. Tau-targeting therapies for Alzheimer disease. Nat. Rev. Neurol. 2018, 14, 399–415. [Google Scholar] [CrossRef]
- Valente, T.; Gella, A.; Fernàndez-Busquets, X.; Unzeta, M.; Durany, N. Immunohistochemical analysis of human brain suggests pathological synergism of Alzheimer’s disease and diabetes mellitus. Neurobiol. Dis. 2010, 37, 67–76. [Google Scholar] [CrossRef]
- Donath, M.Y.; Shoelson, S.E. Type 2 diabetes as an inflammatory disease. Nat. Rev. Immunol. 2011, 11, 98–107. [Google Scholar] [CrossRef]
- Baker, R.G.; Hayden, M.S.; Ghosh, S. NF-κB, Inflammation, and Metabolic Disease. Cell Metab. 2011, 13, 11–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Huo, F.; Liu, B.; Liu, J.; Chen, T.; Li, J.; Zhu, Z.; Lv, B. Crocin Inhibits Oxidative Stress and Pro-inflammatory Response of Microglial Cells Associated with Diabetic Retinopathy Through the Activation of PI3K/Akt Signaling Pathway. J. Mol. Neurosci. 2017, 61, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Pugazhenthi, S.; Qin, L.; Reddy, P.H. Common neurodegenerative pathways in obesity, diabetes, and Alzheimer’s disease. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2017, 1863, 1037–1045. [Google Scholar] [CrossRef]
- Choe, J.-Y.; Kim, S.-K. Quercetin and Ascorbic Acid Suppress Fructose-Induced NLRP3 Inflammasome Activation by Blocking Intracellular Shuttling of TXNIP in Human Macrophage Cell Lines. Inflammation 2017, 40, 980–994. [Google Scholar] [CrossRef]
- Zhou, R.; Yazdi, A.S.; Menu, P.; Tschopp, J. A role for mitochondria in NLRP3 inflammasome activation. Nature 2011, 469, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Wan, Z.; Fan, Y.; Liu, X.; Xue, J.; Han, Z.; Zhu, C.; Wang, X. NLRP3 inflammasome promotes diabetes-induced endothelial inflammation and atherosclerosis. Diabetes Metab. Syndr. Obes. 2019, 12, 1931–1942. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.L.; Shen, X.C. Scutellarin improves LPS+ATP induced inflammation and pyroptosis of endothelial cells by inhibiting NLRP3/caspase-1 signaling pathway, Chin. Pharmacol. Bull. 2022, 38, 1196–1201. [Google Scholar]
- Thornalley, P.J. Dicarbonyl Intermediates in the Maillard Reaction. Ann. N. Y. Acad. Sci. 2005, 1043, 111–117. [Google Scholar] [CrossRef]
- Thornalley, P.J. Endogenous α-Oxoaldehydes and Formation of Protein and Nucleotide Advanced Glycation Endproducts in Tissue Damage. In Acetaldehyde-Related Pathology: Bridging the Trans-Disciplinary Divide: Novartis Foundation; Chadwick, D.J., Goode, J., Eds.; John Wiley & Sons, Ltd.: Chichester, UK, 2007; Volume 285, pp. 229–246, discussion 243–246. [Google Scholar] [CrossRef]
- Shinohara, M.; Thornalley, P.J.; Giardino, I.; Beisswenger, P.; Thorpe, S.R.; Onorato, J.; Brownlee, M. Overexpression of glyoxalase-I in bovine endothelial cells inhibits intracellular advanced glycation endproduct formation and prevents hyperglycemia-induced increases in macromolecular endocytosis. J. Clin. Investig. 1998, 101, 1142–1147. [Google Scholar] [CrossRef]
- Thornalley, P. Glyoxalase I—Structure, function and a critical role in the enzymatic defence against glycation. Biochem. Soc. Trans. 2003, 31, 1343–1348. [Google Scholar] [CrossRef]
- Zanotto, C.; Hansen, F.; Galland, F.; Batassini, C.; Federhen, B.C.; da Silva, V.F.; Leite, M.C.; Nardin, P.; Gonçalves, C.-A. Glutamatergic Alterations in STZ-Induced Diabetic Rats Are Reversed by Exendin-4. Mol. Neurobiol. 2018, 56, 3538–3551. [Google Scholar] [CrossRef] [PubMed]
- Newsholme, P.; Cruzat, V.F.; Keane, K.N.; Carlessi, R.; Homem de Bittencourt, P.I., Jr. Molecular mechanisms of ROS production and oxidative stress in diabetes. Biochem. J. 2016, 473, 4527–4550. [Google Scholar] [CrossRef] [PubMed]
- Sireesh, D.; Ganesh, M.-R.; Dhamodharan, U.; Sakthivadivel, M.; Sivasubramanian, S.; Gunasekaran, P.; Ramkumar, K.M. Role of pterostilbene in attenuating immune mediated devastation of pancreatic beta cells via Nrf2 signaling cascade. J. Nutr. Biochem. 2017, 44, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.L.; Goldfine, I.D.; Maddux, B.A.; Grodsky, G.M. Are Oxidative Stress−Activated Signaling Pathways Mediators of Insulin Resistance and β-Cell Dysfunction? Diabetes 2003, 52, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Siti, H.N.; Kamisah, Y.; Kamsiah, J. The role of oxidative stress, antioxidants and vascular inflammation in cardiovascular disease (a review). Vasc. Pharmacol. 2015, 71, 40–56. [Google Scholar] [CrossRef]
- Smith, M.; Wilkinson, S. ER homeostasis and autophagy. Essays Biochem. 2017, 61, 625–635. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, F.; Fu, M.; Wang, C.; Quon, M.J.; Yang, P. Cellular Stress, Excessive Apoptosis, and the Effect of Metformin in a Mouse Model of Type 2 Diabetic Embryopathy. Diabetes 2015, 64, 2526–2536. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Wang, F.; Reece, E.A.; Yang, P. Curcumin ameliorates high glucose-induced neural tube defects by suppressing cellular stress and apoptosis. Am. J. Obstet. Gynecol. 2015, 212, 802.e1–802.e8. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Kaufman, R.J. From endoplasmic-reticulum stress to the inflammatory response. Nature 2008, 454, 455–462. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Wu, C.; Ye, L.; Wang, B.; Yuan, Y.; Liu, Y.; Zheng, P.; Xiong, J.; Li, Y.; Jiang, T.; et al. Exogenous fibroblast growth factor 1 ameliorates diabetes-induced cognitive decline via coordinately regulating PI3K/AKT signaling and PERK signaling. Cell Commun. Signal. 2020, 18, 81. [Google Scholar] [CrossRef]
- Baur, J.A. Biochemical effects of SIRT1 activators. Biochim. Biophys. Acta (BBA) Proteins Proteom. 2010, 1804, 1626–1634. [Google Scholar] [CrossRef] [Green Version]
- Feng, K.; Chen, Z.; Pengcheng, L.; Zhang, S.; Wang, X. Quercetin attenuates oxidative stress-induced apoptosis via SIRT1/AMPK-mediated inhibition of ER stress in rat chondrocytes and prevents the progression of osteoarthritis in a rat model. J. Cell. Physiol. 2019, 234, 18192–18205. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.-J.; Xu, T.-T.; Li, L.; Xu, Y.-M.; Qu, Z.-L.; Wang, X.-C.; Huang, S.-Q.; Luo, Y.; Luo, N.-C.; Lu, P.; et al. Bushen-Yizhi formula ameliorates cognitive dysfunction through SIRT1/ER stress pathway in SAMP8 mice. Oncotarget 2017, 8, 49338–49350. [Google Scholar] [CrossRef] [Green Version]
- Lourenco, M.V.; Clarke, J.R.; Frozza, R.L.; Bomfim, T.R.; Forny-Germano, L.; Batista, A.F.; Sathler, L.B.; Brito-Moreira, J.; Amaral, O.B.; Silva, C.A.; et al. TNF-α Mediates PKR-Dependent Memory Impairment and Brain IRS-1 Inhibition Induced by Alzheimer’s β-Amyloid Oligomers in Mice and Monkeys. Cell Metab. 2013, 18, 831–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagatys, E.; Garrett, C.R.; Boulware, D.; Kelley, S.; Malafa, M.; Cheng, J.Q.; Sebti, S.; Coppola, D. Activation of the serine/threonine protein kinase Akt during the progression of Barrett neoplasia. Hum. Pathol. 2007, 38, 1526–1531. [Google Scholar] [CrossRef]
- Vanhaesebroeck, B.; Guillermet-Guibert, J.; Graupera, M.; Bilanges, B. The emerging mechanisms of isoform-specific PI3K signalling. Nat. Rev. Mol. Cell Biol. 2010, 11, 329–341. [Google Scholar] [CrossRef] [PubMed]
- Brazil, D.P.; Yang, Z.-Z.; Hemmings, B.A. Advances in protein kinase B signalling: AKTion on multiple fronts. Trends Biochem. Sci. 2004, 29, 233–242. [Google Scholar] [CrossRef]
- Kanaan, N.M.; Morfini, G.A.; Lapointe, N.E.; Pigino, G.F.; Patterson, K.R.; Song, Y.; Andreadis, A.; Fu, Y.; Brady, S.T.; Binder, L.I. Pathogenic Forms of Tau Inhibit Kinesin-Dependent Axonal Transport through a Mechanism Involving Activation of Axonal Phosphotransferases. J. Neurosci. 2011, 31, 9858–9868. [Google Scholar] [CrossRef]
- Zakharenko, S.S.; Patterson, S.L.; Dragatsis, I.; Zeitlin, S.O.; Siegelbaum, S.A.; Kandel, E.R.; Morozov, A. Presynaptic BDNF Required for a Presynaptic but Not Postsynaptic Component of LTP at Hippocampal CA1-CA3 Synapses. Neuron 2003, 39, 975–990. [Google Scholar] [CrossRef] [Green Version]
- Tyler, W.; Pozzo-Miller, L.D. BDNF Enhances Quantal Neurotransmitter Release and Increases the Number of Docked Vesicles at the Active Zones of Hippocampal Excitatory Synapses. J. Neurosci. 2001, 21, 4249–4258. [Google Scholar] [CrossRef] [Green Version]
- Ying, S.W.; Futter, M.; Rosenblum, K.; Webber, M.J.; Hunt, S.P.; Bliss, T.V.P.; Bramham, C.R. Brain-derived neurotrophic factor induces long-term potentiation in intact adult hippocampus: Requirement for ERK activation coupled to CREB and upregulation of Arc synthesis. J. Neurosci. 2002, 22, 1532–1540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, A.L.; Caldeira, M.; Santos, S.; Duarte, C.B. Role of the brain-derived neurotrophic factor at glutamatergic synapses. J. Cereb. Blood Flow Metab. 2008, 153, S310–S324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, T.-J.; Wang, D.-C.; Chen, S.-S. Amyloid-β interrupts the PI3K-Akt-mTOR signaling pathway that could be involved in brain-derived neurotrophic factor-induced Arc expression in rat cortical neurons. J. Neurosci. Res. 2009, 87, 2297–2307. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Wang, X.; O’Connor, M.; Wang, G.; Han, F. Brain-Derived Neurotrophic Factor and Its Potential Therapeutic Role in Stroke Comorbidities. Neural Plast. 2020, 2020, 1–13. [Google Scholar] [CrossRef]
- Hill, J.; Rom, S.; Ramirez, S.H.; Persidsky, Y. Emerging Roles of Pericytes in the Regulation of the Neurovascular Unit in Health and Disease. J. Neuroimmune Pharmacol. 2014, 9, 591–605. [Google Scholar] [CrossRef] [Green Version]
- Bogush, M.; Heldt, N.A.; Persidsky, Y. Blood Brain Barrier Injury in Diabetes: Unrecognized Effects on Brain and Cognition. J. Neuroimmune Pharmacol. 2017, 12, 593–601. [Google Scholar] [CrossRef]
- Takechi, R.; Lam, V.; Brook, E.; Giles, C.; Fimognari, N.; Mooranian, A.; Al-Salami, H.; Coulson, S.H.; Nesbit, M.; Mamo, J. Blood-Brain Barrier Dysfunction Precedes Cognitive Decline and Neurodegeneration in Diabetic Insulin Resistant Mouse Model: An Implication for Causal Link. Front. Aging Neurosci. 2017, 9, 399. [Google Scholar] [CrossRef] [Green Version]
- Rom, S.; Zuluaga-Ramirez, V.; Gajghate, S.; Seliga, A.; Winfield, M.; Heldt, N.A.; Kolpakov, M.A.; Bashkirova, Y.V.; Sabri, A.K.; Persidsky, Y. Hyperglycemia-Driven Neuroinflammation Compromises BBB Leading to Memory Loss in Both Diabetes Mellitus (DM) Type 1 and Type 2 Mouse Models. Mol. Neurobiol. 2018, 56, 1883–1896. [Google Scholar] [CrossRef]
- Lv, S.; Song, H.-L.; Zhou, Y.; Li, L.-X.; Cui, W.; Wang, W.; Liu, P. Tumour necrosis factor-α affects blood-brain barrier permeability and tight junction-associated occludin in acute liver failure. Liver Int. 2010, 30, 1198–1210. [Google Scholar] [CrossRef]
- Geng, J.; Wang, L.; Zhang, L.; Qin, C.; Song, Y.; Ma, Y.; Chen, Y.; Chen, S.; Wang, Y.; Zhang, Z.; et al. Blood-Brain Barrier Disruption Induced Cognitive Impairment Is Associated With Increase of Inflammatory Cytokine. Front. Aging Neurosci. 2018, 10, 129. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, A.; Khera, A.; Singh, I.; Sandhir, R. S-nitrosoglutathione prevents blood-brain barrier disruption associated with increased matrix metalloproteinase-9 activity in experimental diabetes. J. Neurochem. 2014, 132, 595–608. [Google Scholar] [CrossRef] [PubMed]
- Enciu, A.-M.; Gherghiceanu, M.; Popescu, B.O. Triggers and Effectors of Oxidative Stress at Blood-Brain Barrier Level: Relevance for Brain Ageing and Neurodegeneration. Oxidative Med. Cell. Longev. 2013, 2013, 297512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, E.K.; Olson, D.M.; Bernlohr, D.A. High-fat diet induces changes in adipose tissue trans-4-oxo-2-nonenal and trans-4-hydroxy-2-nonenal levels in a depot-specific manner. Free. Radic. Biol. Med. 2013, 63, 390–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ju, W.Z.; Zhang, J.; Tan, H.S.; Jiang, M.; Chen, M.; Xiong, N. Determination of Scutellarin in Human Plasma by LC-MS Method and its Clinical Pharmacokinetics in Chinese Healthy Volunteers. Chin. J. Clin. Pharmacol. 2005, 3, 298–301. [Google Scholar]
- Wang, L.; Ma, Q. Clinical benefits and pharmacology of scutellarin: A comprehensive review. Pharmacol. Ther. 2018, 190, 105–127. [Google Scholar] [CrossRef] [PubMed]
- Mamadalieva, N.Z.; Herrmann, F.; El-Readi, M.Z.; Tahrani, A.; Hamoud, R.; Egamberdieva, D.R.; Azimova, S.S.; Wink, M. Flavonoids in Scutellaria immaculata and S. ramosissima (Lamiaceae) and their biological activity. J. Pharm. Pharmacol. 2011, 63, 1346–1357. [Google Scholar] [CrossRef]
- Han, W.Z.; Li, Y.T.; Hu, B.; Yu, Y.; Wang, W. Research progress of intestinal flora imbalance involved in cognitive impairment in type 2 diabetes mellitus. Chin. J. Neuroanat. 2020, 36, 583–586. [Google Scholar] [CrossRef]
- Welcome, M.O. Gut Microbiota Disorder, Gut Epithelial and Blood–Brain Barrier Dysfunctions in Etiopathogenesis of Dementia: Molecular Mechanisms and Signaling Pathways. NeuroMolecular Med. 2019, 21, 205–226. [Google Scholar] [CrossRef]
- Rogers, G.B.; Keating, D.; Young, R.; Wong, M.-L.; Licinio, J.; Wesselingh, S. From gut dysbiosis to altered brain function and mental illness: Mechanisms and pathways. Mol. Psychiatry 2016, 21, 738–748. [Google Scholar] [CrossRef] [Green Version]
- Heijtz, R.D.; Wang, S.; Anuar, F.; Qian, Y.; Björkholm, B.; Samuelsson, A.; Hibberd, M.L.; Forssberg, H.; Pettersson, S. Normal gut microbiota modulates brain development and behavior. Proc. Natl. Acad. Sci. USA 2011, 108, 3047–3052. [Google Scholar] [CrossRef] [Green Version]
- Bonaz, B.; Bazin, T.; Pellissier, S. The Vagus Nerve at the Interface of the Microbiota-Gut-Brain Axis. Front. Neurosci. 2018, 12, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalile, B.; Van Oudenhove, L.; Vervliet, B.; Verbeke, K. The role of short-chain fatty acids in microbiota–gut–brain communication. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 461–478. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Zhang, F.; Ding, X.; Wu, G.; Lam, Y.Y.; Wang, X.; Fu, H.; Xue, X.; Lu, C.; Ma, J.; et al. Gut bacteria selectively promoted by dietary fibers alleviate type 2 diabetes. Science 2018, 359, 1151–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Tremaroli, V.; Schmidt, C.; Lundqvist, A.; Olsson, L.M.; Krämer, M.; Gummesson, A.; Perkins, R.; Bergström, G.; Bäckhed, F. The Gut Microbiota in Prediabetes and Diabetes: A Population-Based Cross-Sectional Study. Cell Metab. 2020, 32, 379–390.e3. [Google Scholar] [CrossRef] [PubMed]
- Zheng, H.; Xu, P.; Jiang, Q.; Xu, Q.; Zheng, Y.; Yan, J.; Ji, H.; Ning, J.; Zhang, X.; Li, C.; et al. Correction to: Depletion of acetate-producing bacteria from the gut microbiota facilitates cognitive impairment through the gut-brain neural mechanism in diabetic mice. Microbiome 2021, 9, 164. [Google Scholar] [CrossRef]
- Fung, T.C.; Olson, C.A.; Hsiao, E.Y. Interactions between the microbiota, immune and nervous systems in health and disease. Nat. Neurosci. 2017, 20, 145–155. [Google Scholar] [CrossRef]
- Erny, D.; Hrabě de Angelis, A.L.; Jaitin, D.; Wieghofer, P.; Staszewski, O.; David, E.; Keren-Shaul, H.; Mahlakoiv, T.; Jakobshagen, K.; Buch, T.; et al. Host microbiota constantly control maturation and function of microglia in the CNS. Nat. Neurosci. 2015, 18, 965–977. [Google Scholar] [CrossRef]
- Wang, J.-F.; Fu, S.-P.; Li, S.-N.; Hu, Z.-M.; Xue, W.-J.; Li, Z.-Q.; Huang, B.-X.; Lv, Q.-K.; Liu, J.-X.; Wang, W. Short-Chain Fatty Acids Inhibit Growth Hormone and Prolactin Gene Transcription via cAMP/PKA/CREB Signaling Pathway in Dairy Cow Anterior Pituitary Cells. Int. J. Mol. Sci. 2013, 14, 21474–21488. [Google Scholar] [CrossRef] [Green Version]
- Srikanth, V.; Sinclair, A.J.; Hill-Briggs, F.; Moran, C.; Biessels, G.J. Type 2 diabetes and cognitive dysfunction—Towards effective management of both comorbidities. Lancet Diabetes Endocrinol. 2020, 8, 535–545. [Google Scholar] [CrossRef]
- Chen, M.; Huang, N.; Liu, J.; Huang, J.; Shi, J.; Jin, F. AMPK: A bridge between diabetes mellitus and Alzheimer’s disease. Behav. Brain Res. 2020, 400, 113043. [Google Scholar] [CrossRef]
- Longnus, S.L.; Wambolt, R.B.; Parsons, H.L.; Brownsey, R.W.; Allard, M.F. 5-Aminoimidazole-4-carboxamide 1-β-d-ribofuranoside (AICAR) stimulates myocardial glycogenolysis by allosteric mechanisms. Am. J. Physiol. Integr. Comp. Physiol. 2003, 284, R936–R944. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.J.; Xu, Z.G.; Liu, Z.M.; Shijie, T.; Xianli, Z. Determination of quercetin in Erigeron Breviscapus by reflux extraction under vacuum coupled with high performance liquid chromatograph. Hua Hsueh Yen Chiu 2015, 26, 158–161. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, Y.J.; Sheng, Y.M.; Zhang, Y.; Luo, W.Z. Study on the neuroprotection components of ethnomedicine Erigeron breviscapus. West China J. Pharm. Sci. 2011, 26, 208–211. [Google Scholar] [CrossRef]



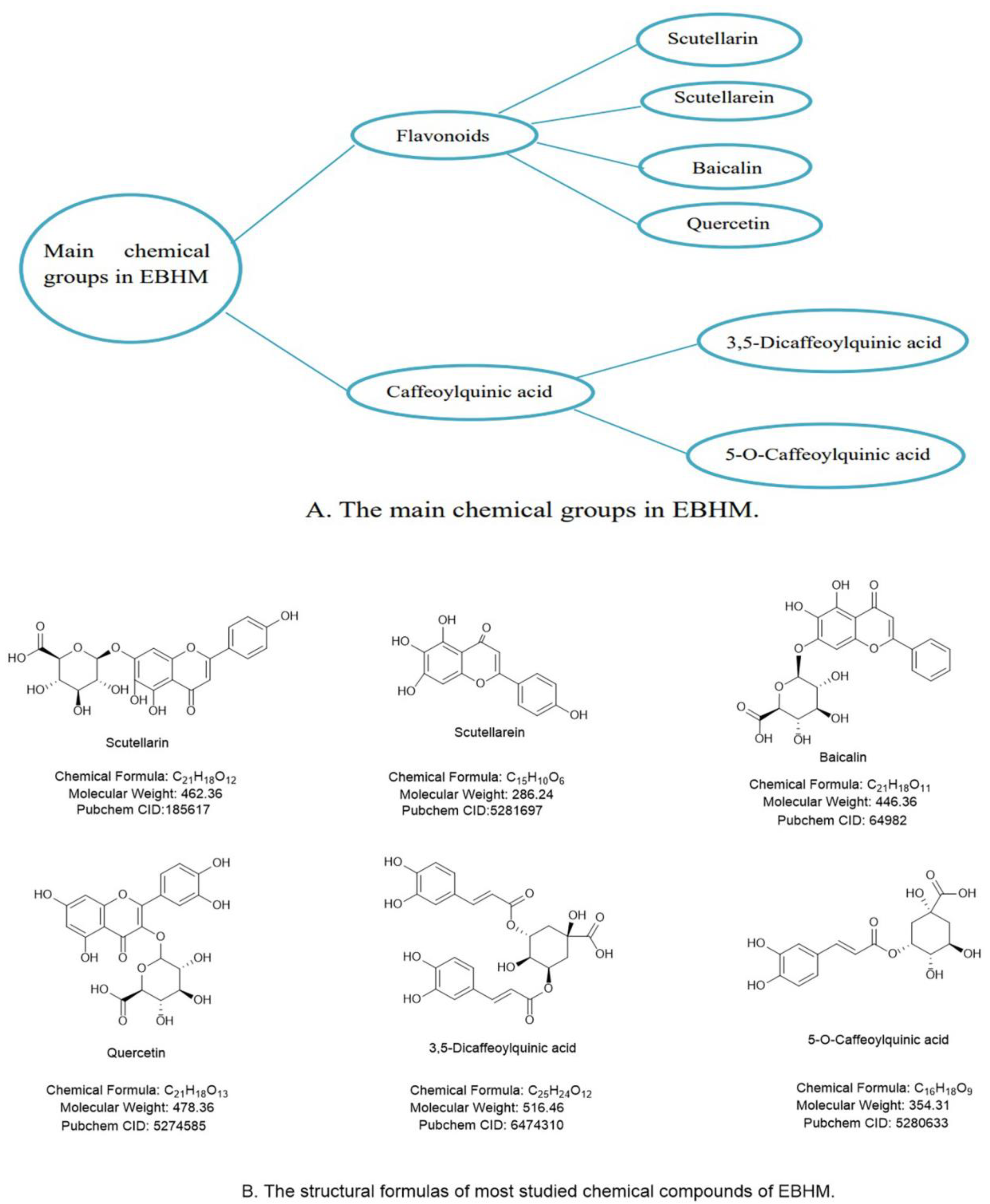{kind=link}
{kind=link}
| Active Ingredients | Model | Administration | Pharmacological Actions | Test Index | Possible Mechanism | References |
|---|---|---|---|---|---|---|
| Scutellarin | APP/PS1 mice | 20/100 mg/kg, p.o. | Regulates intestinal microbiota, decreases Aβ oligomer and downregulates IL-1β by reversing cAMP-PKA-CREB-HDAC3 in microglia. | OFT, NORT | Targeting the composition of intestinal microbiota, Aβ, neuroinflammation | [27] |
| Scutellarin | MCAO rats | 0.5 mg/kg, i.v. | Inhibits MMP-9 transcription | — | Targeting BBB dysfunction | [28] |
| Scutellarein | BV2 cells | 20/50 μM | Reduces TNF-α expression and maintains the integrity of TJs | — | Targeting BBB dysfunction, neuroinflammation | [29] |
| Scutellarin | C57BL/6 mice | 50 mg/kg, p.o. | Activates AMPKα, promotes glucose uptake and increases SOD levels | FBG, INS, HOMA-IR, HOMA-IS, HOMA-β, QUICKI, IPGTT, IPITT | Targeting insulin-resistance | [30] |
| Scutellarin | APP/PS1 mice | 50 mg/kg, i.v. | Lowers cortical levels of soluble human Aβ42 and Aβ40 | EPM, MWM | Targeting Aβ | [31] |
| Scutellarin | db/db mice | 25/50/100 mg/kg, i.g. | Increases SOD activity inhibits MDA production and reduces IL-1β expression | OGTT | Targeting oxidative stress, neuroinflammation | [32] |
| Scutellarin | STZ (35 mg/kg)-induced diabetic rats | 50 mg/kg, i.g. | Increases SOD levels, reduces Bcl-2 levels | MWM, OFT, FBG, INS, HOMA-IR, ISI | Targeting anti-apoptosis, oxidative stress, neuroinflammation | [33] |
| Scutellarin | pBCAO rats | 30 mg/kg, p.o. | Reduces the production of Aβ by preventing the expression of APP and BACE-1 and prevents glial cell activation | MWM | Targeting Aβ, neuroinflammation | [34] |
| Scutellarin | Balb/c male mice(D-gal, AlCl3) | 20 mg/kg, p.o. | Increases acetylcholine and SOD levels while decreasing p-tau and Aβ42 levels | MWM | Targeting t126au protein, Aβ, oxidative stress | [35] |
| Scutellarin | APP/PS1 mice | 50 mg/kg, p.o. | Lowers the expression of pro-inflammatory cytokines and decreases Aβ expression in the brain and plasma | MWM | Targeting Aβ, neuroinflammation | [36] |
| Scutellarin | LPS (500 μg/kg)-induced cognitive impairment mice | 5/25/50 mg/kg, i.g. | Increases SOD activity decreases MDA activity and decreases the level of proinflammatory cytokines | Y-maze, novel object recognition, passive avoidance test | Targeting oxidative stress, neuroinflammation | [37] |
| Baicalin | SH-SY5Y cells | 5/10/20 µM | Improves SH-SY5Y cells activity | — | Targeting anti-apoptosis | [38] |
| Baicalin | Neuron–Astrocyte Cocultures | 34.38 μg/ml | Increases SOD activity inhibits reactive astrocytes production and reduces IL-1β expression | — | Targeting anti-apoptosis, oxidative stress, neuroinflammation | [39] |
| Baicalin | db/db mice | 400 mg/kg, i.g. | Increases SOD activity decreases MDA activity and decreases the level of proinflammatory cytokines | FBG, OGTT, ITT, HOMA-IR | Targeting oxidative stress, neuroinflammation | [40] |
| Baicalin | Male C57BL/6 mice | 200/400 mg/kg, i.g. | Upregulates Insulin receptor expression, and reduces TNF-α expression by modulating macrophage differentiation in the adipose tissue and liver. | OGTT, OFTT, HOMA-IR | Targeting insulin resistance, neuroinflammation | [41] |
| Baicalin | APP/PS1 mice | 100 mg/kg, i.p. | Reduces the quantity of pro-inflammatory cytokines and the amount of activated microglia | MWM, probe test | Targeting neuroinflammation | [42] |
| Baicalin | Male ICR mice (Aβ-injected hippocampus) | 100 mg/kg, i.g. | Reduces inflammatory factor (IL-6, TNF-α) expression and glial cell activation | MWM, probe test | Targeting neuroinflammation | [43] |
| Baicalin | STZ (45 mg/kg)-induced diabetic rats | 80 mg/kg, i.p. | Activates AMPKα, promotes glucose uptake, and inhibits MDA production | — | Targeting insulin-resistance | [44] |
| Quercetin | db/db mice | 70 mg/kg, i.g. | Upregulates SIRT1 protein expression and inhibits the expression of ER signaling pathway-related proteins | OGTT, ITT, MWM | Targeting ER stress | [45] |
| Quercetin | STZ(60 mg/kg)-induced diabetic rats | 50 mg/kg, i.g. | Increases SOD activity decreases MDA activity and decreases the level of proinflammatory cytokines | FBG | Targeting oxidative stress, neuroinflammation | [46] |
| Quercetin | db/db mice | 70 mg/kg, i.g. | Decreases NLRP3 inflammation-related proteins and inflammatory factor (IL-1β and IL-18) expressions | MWM, NORT, OGTT, IRT | Targeting oxidative stress, neuroinflammation | [47] |
| Quercetin | glutamate-exposed HT22 cells | 5/9 µM | Decreases p-tau | — | Targeting tau protein | [48] |
| Quercetin | okadaic acid-exposed HT22 cells | 1/3/5 µM | Decreases p-tau | — | Targeting tau protein | [49] |
| Quercetin | STZ (60 mg/kg)-induced diabetic rats | 90 mg/kg, i.g. | Increases AGEs levels by inhibiting Glo-1 expression, reduces IL-β, and TNF-α expression and increases SOD levels | FBG | Targeting oxidative stress, neuroinflammation | [50] |
| 5-CQA | APP/PS2 transgenic mice | 0.8% (w/w) | Modulates Aβ and neuronal loss | Y-maze, novel object recognition | Targeting Aβ | [24] |
| 3,5-di-CQA | STZ (40 mg/kg)-induced diabetic rats | 100/200 mg/kg, i.p. | Inhibits α -glucosidase activity | OGTT | Targeting insulin resistance | [23] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, S.; Zhou, Z.; Zhang, S.; Cai, Y. Advances in Anti-Diabetic Cognitive Dysfunction Effect of Erigeron Breviscapus (Vaniot) Hand-Mazz. Pharmaceuticals 2023, 16, 50. https://doi.org/10.3390/ph16010050
Gu S, Zhou Z, Zhang S, Cai Y. Advances in Anti-Diabetic Cognitive Dysfunction Effect of Erigeron Breviscapus (Vaniot) Hand-Mazz. Pharmaceuticals. 2023; 16(1):50. https://doi.org/10.3390/ph16010050
Chicago/Turabian StyleGu, Shanye, Ziyi Zhou, Shijie Zhang, and Yefeng Cai. 2023. "Advances in Anti-Diabetic Cognitive Dysfunction Effect of Erigeron Breviscapus (Vaniot) Hand-Mazz" Pharmaceuticals 16, no. 1: 50. https://doi.org/10.3390/ph16010050
APA StyleGu, S., Zhou, Z., Zhang, S., & Cai, Y. (2023). Advances in Anti-Diabetic Cognitive Dysfunction Effect of Erigeron Breviscapus (Vaniot) Hand-Mazz. Pharmaceuticals, 16(1), 50. https://doi.org/10.3390/ph16010050