

Taq-Polymerase Stop Assay to Determine Target Selectivity of G4 Ligands in Native Promoter Sequences of MYC, TERT, and KIT Oncogenes

Abstract
:
1. Introduction
2. Results and Discussion
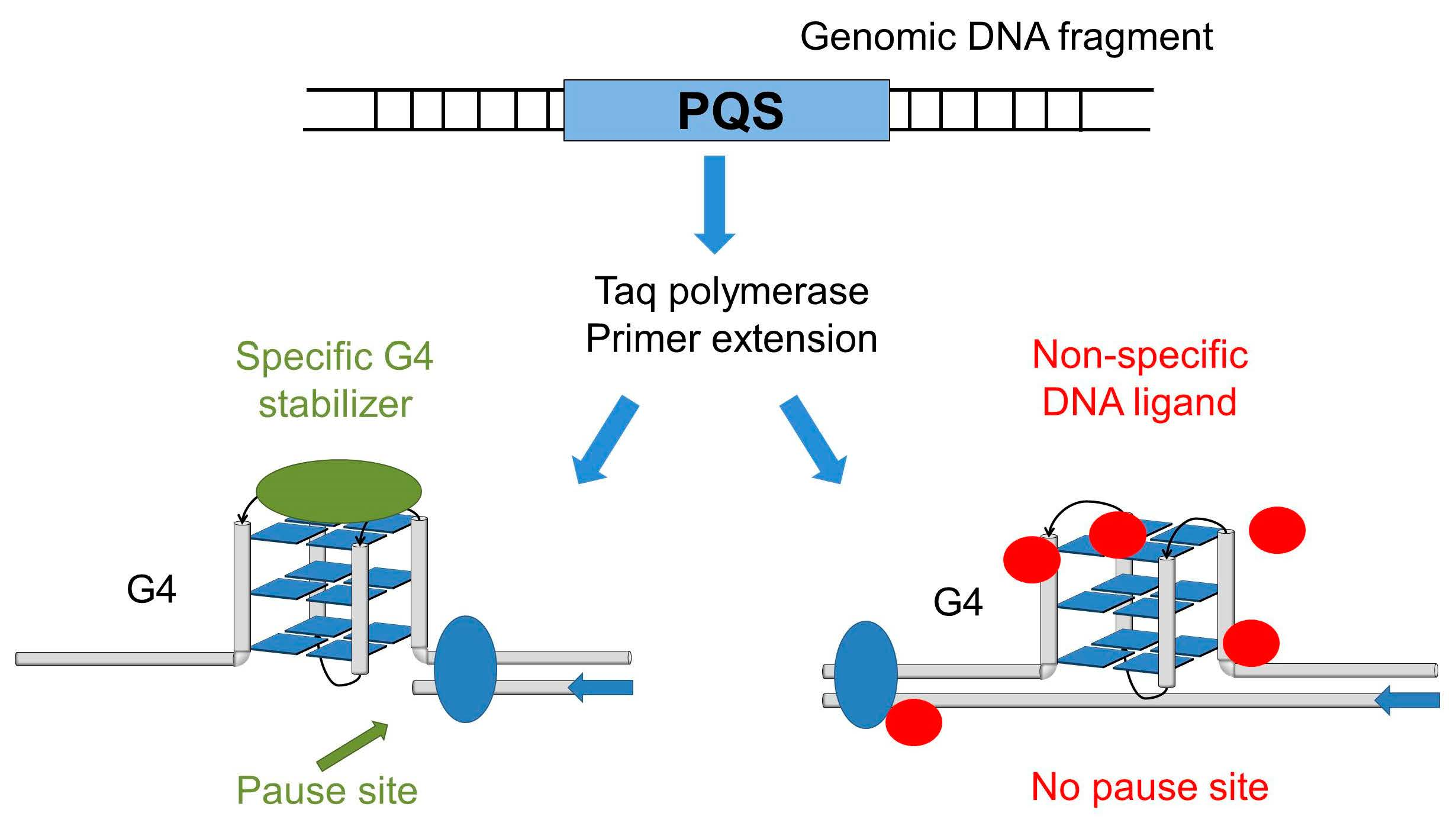
2.1. Pause Sites of the Thermostable DNA Polymerase Are Caused by the Stabilization of G4 Structures by DNA-binding Ligands
2.2. Polymerase Pause Sites Correspond to G4 Sequences
2.3. FRET Melting Assay of DNA Sequences Corresponded to Polymerase Pause Sites
3. Materials and Methods
3.1. Reagents
3.2. Oligonucleotides
3.3. Amplification of Promoter Regions
3.4. Primer Extension in the Presence of Ligands
3.5. FRET Melting Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rhodes, D.; Lipps, H.J. G-quadruplexes and their regulatory roles in biology. Nucleic Acids Res. 2015, 43, 8627–8637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, V.S.; Marsico, G.; Boutell, J.M.; Di Antonio, M.; Smith, G.P.; Balasubramanian, S. High-throughput sequencing of DNA G-quadruplex structures in the human genome. Nat. Biotechnol. 2015, 33, 877–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kosiol, N.; Juranek, S.; Brossart, P.; Heine, A.; Paeschke, K. G-quadruplexes: A promising target for cancer therapy. Mol. Cancer 2021, 20, 40. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, S.; Hurley, L.H.; Neidle, S. Targeting G-quadruplexes in gene promoters: A novel anticancer strategy? Nat. Rev. Drug Discov. 2011, 10, 261–275. [Google Scholar] [CrossRef] [Green Version]
- Asamitsu, S.; Obata, S.; Yu, Z.; Bando, T.; Sugiyama, H. Recent Progress of Targeted G-Quadruplex-Preferred Ligands Toward Cancer Therapy. Molecules. 2019, 24, 429. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Dickerhoff, J.; Sakai, S.; Yang, D. DNA G-Quadruplex in Human Telomeres and Oncogene Promoters: Structures, Functions, and Small Molecule Targeting. Acc. Chem. Res. 2022, 55, 2628–2646. [Google Scholar] [CrossRef]
- Sengupta, P.; Bhattacharya, A.; Sa, G.; Das, T.; Chatterjee, S. Truncated G-Quadruplex Isomers Cross-Talk with the Transcription Factors To Maintain Homeostatic Equilibria in c-MYC Transcription. Biochemistry 2019, 58, 1975–1991. [Google Scholar] [CrossRef]
- Grün, J.T.; Schwalbe, H. Folding dynamics of polymorphic G-quadruplex structures. Biopolymers 2022, 113, e23477. [Google Scholar] [CrossRef]
- Monsen, R.C.; DeLeeuw, L.W.; Dean, W.L.; Gray, R.D.; Chakravarthy, S.; Hopkins, J.B.; Chaires, J.B.; Trent, J.O. Long promoter sequences form higher-order G-quadruplexes: An integrative structural biology study of c-Myc, k-Ras and c-Kit promoter sequences. Nucleic Acids Res. 2022, 50, 4127–4147. [Google Scholar] [CrossRef]
- Monsen, R.C.; Trent, J.O.; Chaires, J.B. G-quadruplex DNA: A Longer Story. Acc. Chem. Res. 2022, 55, 3242–3252. [Google Scholar] [CrossRef]
- Vianney, Y.M.; Weisz, K. High-affinity binding at quadruplex–duplex junctions: Rather the rule than the exception. Nucleic Acids Res. 2022, 50, 11948–11964. [Google Scholar] [CrossRef]
- Bahls, B.; Aljnadi, I.M.; Emídio, R.; Mendes, E.; Paulo, A. G-Quadruplexes in c-MYC Promoter as Targets for Cancer Therapy. Biomedicines 2023, 11, 969. [Google Scholar] [CrossRef]
- Nasiri, H.R.; Bell, N.M.; McLuckie, K.I.; Husby, J.; Abell, C.; Neidle, S.; Balasubramanian, S. Targeting a c-MYC G-quadruplex DNA with a fragment library. Chem. Commun. 2014, 50, 1704–1707. [Google Scholar] [CrossRef]
- Calabrese, D.R.; Chen, X.; Leon, E.C.; Gaikwad, S.M.; Phyo, Z.; Hewitt, W.M.; Alden, S.; Hilimire, T.A.; He, F.; Michalowski, A.M.; et al. Chemical and structural studies provide a mechanistic basis for recognition of the MYC G-quadruplex. Nat. Commun. 2018, 9, 4229. [Google Scholar] [CrossRef] [Green Version]
- Mathad, R.I.; Hatzakis, E.; Dai, J.; Yang, D. c-MYC promoter G-quadruplex formed at the 5’-end of NHE III1 element: Insights into biological relevance and parallel-stranded G-quadruplex stability. Nucleic Acids Res. 2011, 39, 9023–9033. [Google Scholar] [CrossRef]
- Sun, D.; Hurley, L.H. The importance of negative superhelicity in inducing the formation of G-quadruplex and i-motif structures in the c-Myc promoter: Implications for drug targeting and control of gene expression. J. Med. Chem. 2009, 52, 2863–2874. [Google Scholar] [CrossRef] [Green Version]
- Quazi, S. Telomerase gene therapy: A remission toward cancer. Med. Oncol. 2022, 39, 105. [Google Scholar] [CrossRef]
- Palumbo, S.L.; Ebbinghaus, S.W.; Hurley, L.H. Formation of a unique end-to-end stacked pair of G-quadruplexes in the hTERT core promoter with implications for inhibition of telomerase by G-quadruplex-interactive ligands. J. Am. Chem. Soc. 2009, 131, 10878–10891. [Google Scholar] [CrossRef] [Green Version]
- Micheli, E.; Martufi, M.; Cacchione, S.; De Santis, P.; Savino, M. Self-organization of G-quadruplex structures in the hTERT core promoter stabilized by polyaminic side chain perylene derivatives. Biophys. Chem. 2010, 153, 43–53. [Google Scholar] [CrossRef]
- Monsen, R.C.; DeLeeuw, L.; Dean, W.L.; Gray, R.D.; Sabo, T.M.; Chakravarthy, S.; Chaires, J.B.; Trent, J.O. The hTERT core promoter forms three parallel G-quadruplexes. Nucleic Acids Res. 2020, 48, 5720–5734. [Google Scholar] [CrossRef]
- Dreos, R.; Ambrosini, G.; Cavin Périer, R.; Bucher, P. EPD and EPDnew, high-quality promoter resources in the next-generation sequencing era. Nucleic Acids Res. 2013, 41, D157–D164. [Google Scholar] [CrossRef] [PubMed]
- Pathania, S.; Pentikäinen, O.T.; Singh, P.K. A holistic view on c-Kit in cancer: Structure, signaling, pathophysiology and its inhibitors. Biochim. Biophys. Acta. Rev. Cancer 2021, 1876, 188631. [Google Scholar] [CrossRef] [PubMed]
- Ducani, C.; Bernardinelli, G.; Högberg, B.; Keppler, B.K.; Terenzi, A. Interplay of Three G-Quadruplex Units in the KIT Promoter. J. Am. Chem. Soc. 2019, 141, 10205–10213. [Google Scholar] [CrossRef] [PubMed]
- Zorzan, E.; Da Ros, S.; Musetti, C.; Shahidian, L.Z.; Coelho, N.F.; Bonsembiante, F.; Létard, S.; Gelain, M.E.; Palumbo, M.; Dubreuil, P.; et al. Screening of candidate G-quadruplex ligands for the human c-KIT promotorial region and their effects in multiple in-vitro models. Oncotarget 2016, 7, 21658–21675. [Google Scholar] [CrossRef] [Green Version]
- Wei, D.; Husby, J.; Neidle, S. Flexibility and structural conservation in a c-KIT G-quadruplex. Nucleic Acids Res. 2015, 43, 629–644. [Google Scholar] [CrossRef] [Green Version]
- Martino, L.; Pagano, B.; Fotticchia, I.; Neidle, S.; Giancola, C. Shedding light on the interaction between TMPyP4 and human telomeric quadruplexes. J. Phys. Chem. B 2009, 113, 14779–14786. [Google Scholar] [CrossRef]
- Rodriguez, R.; Miller, K.M.; Forment, J.V.; Bradshaw, C.R.; Nikan, M.; Britton, S.; Oelschlaegel, T.; Xhemalce, B.; Balasubramanian, S.; Jackson, S.P. Small-molecule–induced DNA damage identifies alternative DNA structures in human genes. Nat. Chem. Biol. 2012, 8, 301–310. [Google Scholar] [CrossRef] [Green Version]
- De Cian, A.; DeLemos, E.; Mergny, J.-L.; Teulade-Fichou, M.-P.; Monchaud, D. Highly efficient G-quadruplex recognition by bisquinolinium compounds. J. Am. Chem. Soc. 2007, 129, 1856–1857. [Google Scholar] [CrossRef]
- Burger, A.M.; Dai, F.; Schultes, C.M.; Reszka, A.P.; Moore, M.J.; Double, J.A.; Neidle, S. The G-quadruplex-interactive molecule BRACO-19 inhibits tumor growth, consistent with telomere targeting and interference with telomerase function. Cancer Res. 2005, 65, 1489–1496. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Han, H. A DNA Polymerase Stop Assay for Characterization of G-Quadruplex Formation and Identification of G-Quadruplex-Interactive Compounds. In G-Quadruplex Nucleic Acids: Methods and Protocols; Yang, D., Lin, C., Eds.; Springer: New York, NY, USA, 2019; pp. 223–231. [Google Scholar] [CrossRef]
- Han, H.; Hurley, L.H.; Salazar, M. A DNA polymerase stop assay for G-quadruplex-interactive compounds. Nucleic Acids Res 1999, 27, 537–542. [Google Scholar] [CrossRef]
- Chashchina, G.V.; Beniaminov, A.D.; Kaluzhny, D.N. Stable G-Quadruplex Structures of Oncogene Promoters Induce Potassium-Dependent Stops of Thermostable DNA Polymerase. Biochemistry 2019, 84, 562–569. [Google Scholar] [CrossRef]
- Niknezhad, Z.; Hassani, L.; Norouzi, D. Investigating actinomycin D binding to G-quadruplex, i-motif and double-stranded DNA in 27-nt segment of c-MYC gene promoter. Mater. Sci. Eng. C 2016, 58, 1188–1193. [Google Scholar] [CrossRef]
- Bilgen, E.; Persil Çetinkol, Ö. Doxorubicin exhibits strong and selective association with VEGF Pu22 G-quadruplex. Biochim. Biophys. Acta BBA Gen. Subj. 2020, 1864, 129720. [Google Scholar] [CrossRef]
- De Rache, A.; Mergny, J.L. Assessment of selectivity of G-quadruplex ligands via an optimised FRET melting assay. Biochimie 2015, 115, 194–202. [Google Scholar] [CrossRef]
- Beniaminov, A.D.; Chashchina, G.V.; Livshits, M.A.; Kechko, O.I.; Mitkevich, V.A. Discrimination between G/C Binding Sites by Olivomycin A Is Determined by Kinetics of the Drug-DNA Interaction. Int. J. Mol. Sci. 2020, 21, 5299. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oligonucleotide Name | KCl | PDS | TMPyP4 | PhenDC3 | BRACO-19 |
|---|---|---|---|---|---|
| QMYC-1 | 66.4 ± 1.4 | 83.8 ± 1.1 | >95 | >95 | 85.4 ± 0.3 |
| QMYC-2 | 66.4 ± 1.1 | 81.1 ± 3.8 | >95 | 91.3 ± 1.6 | 82.1 ± 0.4 |
| QMYC-3 | 44 ± 3.1 | 80.3 ± 1.8 | >95 | 86.2 ± 0.3 | 78.3 ± 0.7 |
| QTERT-1 | 55.7 ± 1.8 | 74.3 ± 1.3 | ND | 86.4 ± 0 | 81 ± 0.6 |
| QTERT-2 | 58.6 ± 0.8 | 79.6 ± 0.3 | ND | 85.4 ± 0 | 82.2 ± 0.6 |
| QKIT-1 | 33.6 ± 0.8 | 75 ± 2.8 | >95 | 74.1 ± 3.3 | 77.9 ± 0.1 |
| QKIT-n1 | 56.7 ± 0.4 | 68.2 ± 2 | >95 | 87.4 ± 1.4 | 79.1 ± 0.1 |
| QKIT-sp | 56.3 ± 1 | 68.7 ± 1.6 | >95 | 81.4 ± 0.3 | 75.6 ± 6.5 |
| QKIT-n2 | 41.1 ± 0.4 | 79.8 ± 0.8 | >95 | 89 ± 0 | 77 ± 0.8 |
| QKIT-2 | 48.3 ± 1.3 | 78.3 ± 1.8 | >95 | 83.9 ± 0.7 | 79.7 ± 0.1 |
| Primers for PCR | Fluorescently Labeled Primers for Localizing DNA Polymerase Pause Sites | |
|---|---|---|
| MYC | For: GAGGAGCAGCAGAGAAAGGG Rev: TCCCTCCGTTCTTTTTCCCG | FAM-TCCTAGAGCTAGAGTGCTCGG |
| KIT | For: GTGGAGAGAGAAAGGGGCTC Rev: AAGCAGTAGGAGCAGAACGC | FAM-GCGGCAAAGCCGAGCC |
| TERT | For: GGCCGATTCGACCTCTCT Rev: CAGCGCTGCCTGAAACTC | FAM-CTTCCAGCTCCGCCTCCTCC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chashchina, G.V.; Tevonyan, L.L.; Beniaminov, A.D.; Kaluzhny, D.N. Taq-Polymerase Stop Assay to Determine Target Selectivity of G4 Ligands in Native Promoter Sequences of MYC, TERT, and KIT Oncogenes. Pharmaceuticals 2023, 16, 544. https://doi.org/10.3390/ph16040544
Chashchina GV, Tevonyan LL, Beniaminov AD, Kaluzhny DN. Taq-Polymerase Stop Assay to Determine Target Selectivity of G4 Ligands in Native Promoter Sequences of MYC, TERT, and KIT Oncogenes. Pharmaceuticals. 2023; 16(4):544. https://doi.org/10.3390/ph16040544
Chicago/Turabian StyleChashchina, Galina V., Liana L. Tevonyan, Artemy D. Beniaminov, and Dmitry N. Kaluzhny. 2023. "Taq-Polymerase Stop Assay to Determine Target Selectivity of G4 Ligands in Native Promoter Sequences of MYC, TERT, and KIT Oncogenes" Pharmaceuticals 16, no. 4: 544. https://doi.org/10.3390/ph16040544
APA StyleChashchina, G. V., Tevonyan, L. L., Beniaminov, A. D., & Kaluzhny, D. N. (2023). Taq-Polymerase Stop Assay to Determine Target Selectivity of G4 Ligands in Native Promoter Sequences of MYC, TERT, and KIT Oncogenes. Pharmaceuticals, 16(4), 544. https://doi.org/10.3390/ph16040544