

rAAV2-Mediated Restoration of GALC in Neural Stem Cells from Krabbe Patient-Derived iPSCs


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
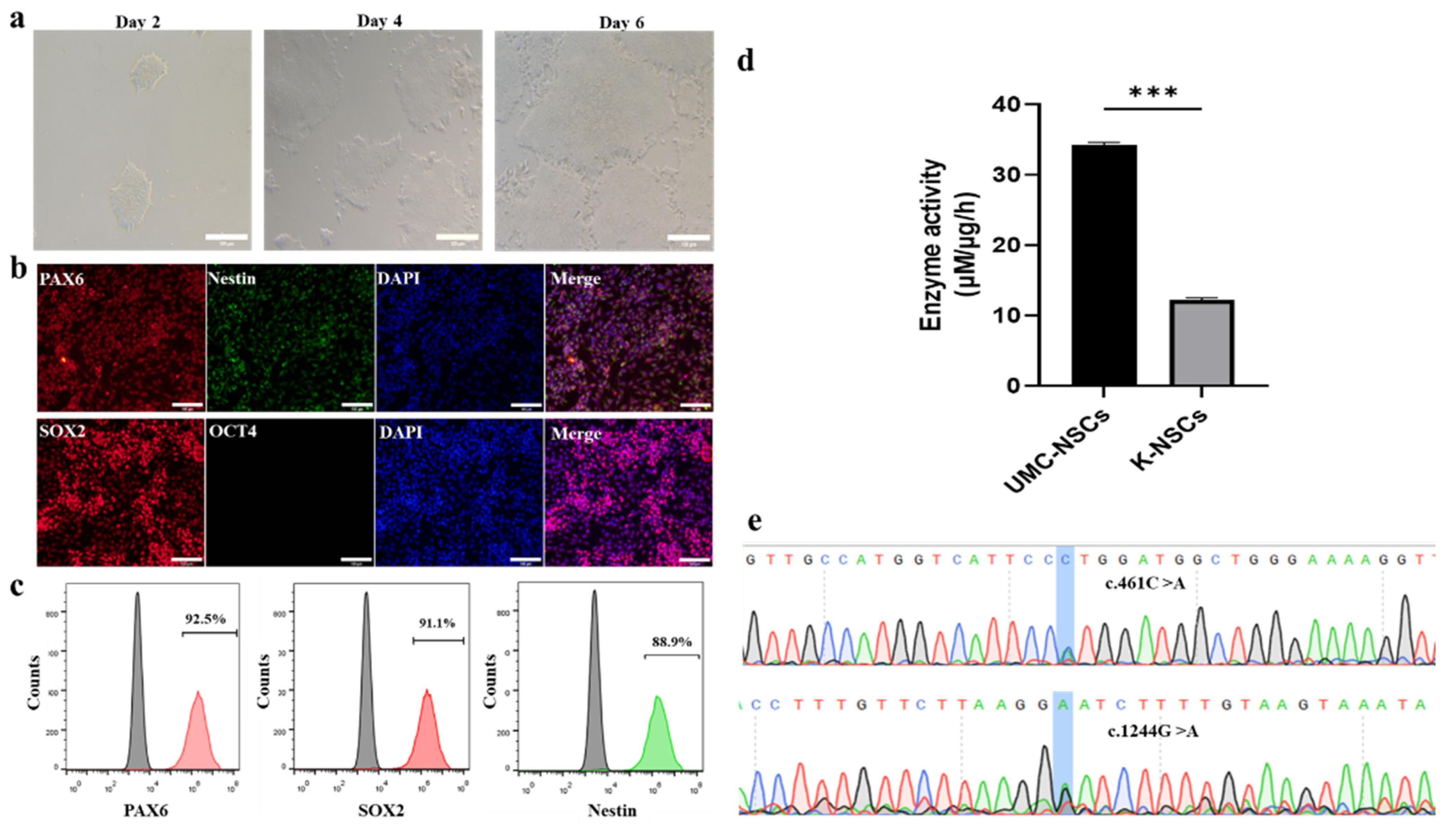
2.1. K-NSCs Are Induced Successfully from K-iPSCs
2.2. The rAAV2 Vector Efficiently Infects K-NSCs
2.3. rAAV2-GALC Restores GALC Enzymatic Activity in K-NSCs
3. Discussion
4. Materials and Methods
4.1. Antibodies
4.2. NSC Induction and Culture
4.3. Immunofluorescence Staining
4.4. Sanger Sequencing
4.5. Flow Cytometry Assays
4.6. rAAV Packaging and Purification
4.7. rAAV Titer Measurement
4.8. Western Blotting
4.9. NSC Infection
4.10. GALC Enzymatic Assay
4.11. Determination of the Psychosine Concentration
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wenger, D.A.; Rafi, M.A.; Luzi, P. Molecular genetics of Krabbe disease (globoid cell leukodystrophy): Diagnostic and clinical implications. Hum. Mutat. 1997, 10, 268–279. [Google Scholar] [CrossRef]
- Wenger, D.A.; Rafi, M.A.; Luzi, P.; Datto, J.; Costantino-Ceccarini, E. Krabbe disease: Genetic aspects and progress toward therapy. Mol. Genet. Metab. 2000, 70, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Feltri, M.L.; Weinstock, N.I.; Favret, J.; Dhimal, N.; Wrabetz, L.; Shin, D. Mechanisms of demyelination and neurodegeneration in globoid cell leukodystrophy. Glia 2021, 69, 2309–2331. [Google Scholar] [CrossRef] [PubMed]
- Potter, G.B.; Petryniak, M.A. Neuroimmune mechanisms in Krabbe’s disease. J. Neurosci. Res. 2016, 94, 1341–1348. [Google Scholar] [CrossRef] [PubMed]
- Voccoli, V.; Tonazzini, I.; Signore, G.; Caleo, M.; Cecchini, M. Role of extracellular calcium and mitochondrial oxygen species in psychosine-induced oligodendrocyte cell death. Cell Death Dis. 2014, 5, e1529. [Google Scholar] [CrossRef]
- Nasir, G.; Chopra, R.; Elwood, F.; Ahmed, S.S. Krabbe Disease: Prospects of Finding a Cure Using AAV Gene Therapy. Front. Med.-Lausanne 2021, 8, 760236. [Google Scholar] [CrossRef]
- Orsini, J.J.; Escolar, M.L.; Wasserstein, M.P.; Caggana, M. Krabbe Disease. In GeneReviews((R)); Adam, M.P., Mirzaa, G.M., Pagon, R.A., Wallace, S.E., Bean, L.J.H., Gripp, K.W., Amemiya, A., Eds.; University of Washington: Seattle, WA, USA, 1993. [Google Scholar]
- Duffner, P.K.; Barczykowski, A.; Jalal, K.; Yan, L.; Kay, D.M.; Carter, R.L. Early infantile Krabbe disease: Results of the World-Wide Krabbe Registry. Pediatr. Neurol. 2011, 45, 141–148. [Google Scholar] [CrossRef]
- Beltran-Quintero, M.L.; Bascou, N.A.; Poe, M.D.; Wenger, D.A.; Saavedra-Matiz, C.A.; Nichols, M.J.; Escolar, M.L. Early progression of Krabbe disease in patients with symptom onset between 0 and 5 months. Orphanet. J. Rare Dis. 2019, 14, 46. [Google Scholar] [CrossRef]
- Escolar, M.L.; Poe, M.D.; Provenzale, J.M.; Richards, K.C.; Allison, J.; Wood, S.; Wenger, D.A.; Pietryga, D.; Wall, D.; Champagne, M.; et al. Transplantation of umbilical-cord blood in babies with infantile Krabbe’s disease. N. Engl. J. Med. 2005, 352, 2069–2081. [Google Scholar] [CrossRef]
- Wright, M.D.; Poe, M.D.; DeRenzo, A.; Haldal, S.; Escolar, M.L. Developmental outcomes of cord blood transplantation for Krabbe disease A 15-year study. Neurology 2017, 89, 1365–1372. [Google Scholar] [CrossRef]
- Wenger, D.A. Murine, canine and non-human primate models of Krabbe disease. Mol. Med. Today 2000, 6, 449–451. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.P.; Chuang, C.Y.; Kuo, H.C. Induced pluripotent stem cell technology for disease modeling and drug screening with emphasis on lysosomal storage diseases. Stem Cell Res. Ther. 2012, 3, 34. [Google Scholar] [CrossRef] [PubMed]
- Bradbury, A.M.; Bongarzone, E.R.; Sands, M.S. Krabbe disease: New hope for an old disease. Neurosci. Lett. 2021, 752, 135841. [Google Scholar] [CrossRef] [PubMed]
- Mangiameli, E.; Cecchele, A.; Morena, F.; Sanvito, F.; Matafora, V.; Cattaneo, A.; Della Volpe, L.; Gnani, D.; Paulis, M.; Susani, L.; et al. Human iPSC-based neurodevelopmental models of globoid cell leukodystrophy uncover patient- and cell type-specific disease phenotypes. Stem Cell Rep. 2021, 16, 1478–1495. [Google Scholar] [CrossRef]
- Wang, W.; Lv, Y.F.; Zhang, Y.J.; Dong, W.J.; Zhang, Y. Generation of a human induced pluripotent stem cell line PUMCi001-A from a patient with Krabbe disease. Stem Cell Res. 2020, 48, 101937. [Google Scholar] [CrossRef]
- De Filippis, L.; Zalfa, C.; Ferrari, D. Neural Stem Cells and Human Induced Pluripotent Stem Cells to Model Rare CNS Diseases. CNS Neurol. Disord. Drug Targets 2017, 16, 915–926. [Google Scholar] [CrossRef]
- Szlachcic, W.J.; Wiatr, K.; Trzeciak, M.; Figlerowicz, M.; Figiel, M. The Generation of Mouse and Human Huntington Disease iPS Cells Suitable for In vitro Studies on Huntingtin Function. Front. Mol. Neurosci. 2017, 10, 253. [Google Scholar] [CrossRef]
- Brennand, K.J.; Simone, A.; Jou, J.; Gelboin-Burkhart, C.; Tran, N.; Sangar, S.; Li, Y.; Mu, Y.L.; Chen, G.; Yu, D.; et al. Modelling schizophrenia using human induced pluripotent stem cells. Nature 2011, 473, 221–225. [Google Scholar] [CrossRef]
- Choi, H.W.; Kim, J.S.; Choi, S.; Hong, Y.J.; Kim, M.J.; Seo, H.G.; Do, J.T. Neural stem cells differentiated from iPS cells spontaneously regain pluripotency. Stem Cells 2014, 32, 2596–2604. [Google Scholar] [CrossRef]
- Fuentealba, L.C.; Rompani, S.B.; Parraguez, J.I.; Obernier, K.; Romero, R.; Cepko, C.L.; Alvarez-Buylla, A. Embryonic Origin of Postnatal Neural Stem Cells. Cell 2015, 161, 1644–1655. [Google Scholar] [CrossRef]
- Mendell, J.R.; Al-Zaidy, S.A.; Rodino-Klapac, L.R.; Goodspeed, K.; Gray, S.J.; Kay, C.N.; Boye, S.L.; Boye, S.E.; George, L.A.; Salabarria, S.; et al. Current Clinical Applications of In Vivo Gene Therapy with AAVs. Mol. Ther. 2021, 29, 464–488. [Google Scholar] [CrossRef] [PubMed]
- Kang, L.; Jin, S.; Wang, J.; Lv, Z.; Xin, C.; Tan, C.; Zhao, M.; Wang, L.; Liu, J. AAV vectors applied to the treatment of CNS disorders: Clinical status and challenges. J. Control. Release 2023, 355, 458–473. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Zhong, L.; Danos, O. Exploiting natural diversity of AAV for the design of vectors with novel properties. Methods Mol. Biol. 2011, 807, 93–118. [Google Scholar] [CrossRef]
- Zhao, Z.M.; Anselmo, A.C.; Mitragotri, S. Viral vector-based gene therapies in the clinic. Bioeng. Transl. Med. 2022, 7, e10258. [Google Scholar] [CrossRef]
- Hirsch, M.L.; Green, L.; Porteus, M.H.; Samulski, R.J. Self-complementary AAV mediates gene targeting and enhances endonuclease delivery for double-strand break repair. Gene Ther. 2010, 17, 1175–1180. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Ma, H.I.; Li, J.; Sun, L.; Zhang, J.; Xiao, X. Rapid and highly efficient transduction by double-stranded adeno-associated virus vectors in vitro and in vivo. Gene Ther. 2003, 10, 2105–2111. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Irvine, E.E.; Eleftheriadou, I.; Naranjo, C.J.; Hearn-Yeates, F.; Bosch, L.; Glegola, J.A.; Murdoch, L.; Czerniak, A.; Meloni, I.; et al. Gene replacement ameliorates deficits in mouse and human models of cyclin-dependent kinase-like 5 disorder. Brain 2020, 143, 811–832. [Google Scholar] [CrossRef]
- Grimm, D.; Lee, J.S.; Wang, L.; Desai, T.; Akache, B.; Storm, T.A.; Kay, M.A. In vitro and in vivo gene therapy vector evolution via multispecies interbreeding and retargeting of adeno-associated viruses. J. Virol. 2008, 82, 5887–5911. [Google Scholar] [CrossRef]
- Chan, K.Y.; Jang, M.J.; Yoo, B.B.; Greenbaum, A.; Ravi, N.; Wu, W.L.; Sanchez-Guardado, L.; Lois, C.; Mazmanian, S.K.; Deverman, B.E.; et al. Engineered AAVs for efficient noninvasive gene delivery to the central and peripheral nervous systems. Nat. Neurosci. 2017, 20, 1172–1179. [Google Scholar] [CrossRef]
- Yoon, I.C.; Bascou, N.A.; Poe, M.D.; Szabolcs, P.; Escolar, M.L. Long-term neurodevelopmental outcomes of hematopoietic stem cell transplantation for late-infantile Krabbe disease. Blood 2021, 137, 1719–1730. [Google Scholar] [CrossRef]
- Weinstock, N.I.; Shin, D.; Dhimal, N.; Hong, X.; Irons, E.E.; Silvestri, N.J.; Reed, C.B.; Nguyen, D.; Sampson, O.; Cheng, Y.C.; et al. Macrophages Expressing GALC Improve Peripheral Krabbe Disease by a Mechanism Independent of Cross-Correction. Neuron 2020, 107, 65–81.e69. [Google Scholar] [CrossRef]
- Berardi, A.S.; Pannuzzo, G.; Graziano, A.; Costantino-Ceccarini, E.; Piomboni, P.; Luddi, A. Pharmacological chaperones increase residual beta-galactocerebrosidase activity in fibroblasts from Krabbe patients. Mol. Genet. Metab. 2014, 112, 294–301. [Google Scholar] [CrossRef]
- Martino, S.; Tiribuzi, R.; Tortori, A.; Conti, D.; Visigalli, I.; Lattanzi, A.; Biffi, A.; Gritti, A.; Orlacchio, A. Specific determination of beta-galactocerebrosidase activity via competitive inhibition of beta-galactosidase. Clin. Chem. 2009, 55, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Kreher, C.; Favret, J.; Weinstock, N.I.; Maulik, M.; Hong, X.; Gelb, M.H.; Wrabetz, L.; Feltri, M.L.; Shin, D. Neuron-specific ablation of the Krabbe disease gene galactosylceramidase in mice results in neurodegeneration. PLoS Biol. 2022, 20, e3001661. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Liu, Y.; Huang, J.; Zhang, X.; Wei, C. Current Approaches and Molecular Mechanisms for Directly Reprogramming Fibroblasts Into Neurons and Dopamine Neurons. Front. Aging Neurosci. 2021, 13, 738529. [Google Scholar] [CrossRef] [PubMed]
- Rowe, R.G.; Daley, G.Q. Induced pluripotent stem cells in disease modelling and drug discovery. Nat. Rev. Genet. 2019, 20, 377–388. [Google Scholar] [CrossRef]
- Lee, H.K.; Morin, P.; Xia, W.M. Peripheral blood mononuclear cell-converted induced pluripotent stem cells (iPSCs) from an early onset Alzheimer’s patient. Stem Cell Res. 2016, 16, 213–215. [Google Scholar] [CrossRef] [PubMed]
- Meneghini, V.; Frati, G.; Sala, D.; De Cicco, S.; Luciani, M.; Cavazzin, C.; Paulis, M.; Mentzen, W.; Morena, F.; Giannelli, S.; et al. Generation of Human Induced Pluripotent Stem Cell-Derived Bona Fide Neural Stem Cells for Ex Vivo Gene Therapy of Metachromatic Leukodystrophy. Stem Cell Transl. Med. 2017, 6, 352–368. [Google Scholar] [CrossRef]
- Tubsuwan, A.; Pires, C.; Rasmussen, M.A.; Schmid, B.; Nielsen, J.E.; Hjermind, L.E.; Hall, V.; Nielsen, T.T.; Waldemar, G.; Hyttel, P.; et al. Generation of induced pluripotent stem cells (iPSCs) from an Alzheimer’s disease patient carrying a L150P mutation in PSEN-1. Stem Cell Res. 2016, 16, 110–112. [Google Scholar] [CrossRef]
- Nieuwenhuis, B.; Haenzi, B.; Hilton, S.; Carnicer-Lombarte, A.; Hobo, B.; Verhaagen, J.; Fawcett, J.W. Optimization of adeno-associated viral vector-mediated transduction of the corticospinal tract: Comparison of four promoters. Gene Ther. 2021, 28, 56–74. [Google Scholar] [CrossRef]
- Smith-Arica, J.R.; Thomson, A.J.; Ansell, R.; Chiorini, J.; Davidson, B.; McWhir, J. Infection efficiency of human and mouse embryonic stem cells using adenoviral and adeno-associated viral vectors. Cloning Stem Cells 2003, 5, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.H.; Koerber, J.T.; Kim, J.S.; Asuri, P.; Vazin, T.; Bartel, M.; Keung, A.; Kwon, I.; Park, K.I.; Schaffer, D.V. An evolved adeno-associated viral variant enhances gene delivery and gene targeting in neural stem cells. Mol. Ther. 2011, 19, 667–675. [Google Scholar] [CrossRef] [PubMed]
- Hughes, S.M.; Moussavi-Harami, F.; Sauter, S.L.; Davidson, B.L. Viral-mediated gene transfer to mouse primary neural progenitor cells. Mol. Ther. 2002, 5, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Lu, H.X.; Hao, Z.M.; Jiao, Q.; Xie, W.L.; Zhang, J.F.; Lu, Y.F.; Cai, M.; Wang, Y.Y.; Yang, Z.Q.; Parker, T.; et al. Neurotrophin-3 gene transduction of mouse neural stem cells promotes proliferation and neuronal differentiation in organotypic hippocampal slice cultures. Med. Sci. Monit. 2011, 17, BR305–BR311. [Google Scholar] [CrossRef]
- Qing, K.; Mah, C.; Hansen, J.; Zhou, S.Z.; Dwarki, V.; Srivastava, A. Human fibroblast growth factor receptor 1 is a co-receptor for infection by adeno-associated virus 2. Nat. Med. 1999, 5, 71–77. [Google Scholar] [CrossRef]
- Strazza, M.; Luddi, A.; Carbone, M.; Rafi, M.A.; Costantino-Ceccarini, E.; Wenger, D.A. Significant correction of pathology in brains of twitcher mice following injection of genetically modified mouse neural progenitor cells. Mol. Genet. Metab. 2009, 97, 27–34. [Google Scholar] [CrossRef]
- Lancaster, M.A.; Knoblich, J.A. Generation of cerebral organoids from human pluripotent stem cells. Nat. Protoc. 2014, 9, 2329–2340. [Google Scholar] [CrossRef]
- Rabeling, A.; Goolam, M. Cerebral organoids as an in vitro model to study autism spectrum disorders. Gene Ther. 2022, 1–11. [Google Scholar] [CrossRef]
- Camp, J.G.; Badsha, F.; Florio, M.; Kanton, S.; Gerber, T.; Wilsch-Brauninger, M.; Lewitus, E.; Sykes, A.; Hevers, W.; Lancaster, M.; et al. Human cerebral organoids recapitulate gene expression programs of fetal neocortex development. Proc. Natl. Acad. Sci. USA 2015, 112, 15672–15677. [Google Scholar] [CrossRef]
- Broekman, M.L.D.; Comer, L.A.; Hyman, B.T.; Siena-Esteves, M. Adeno-associated virus vectors serotyped with AAV8 capsid are more efficient than AAV-1 or-2 serotypes for widespread gene delivery to the neonatal mouse brain. Neuroscience 2006, 138, 501–510. [Google Scholar] [CrossRef]
- Song, R.; Pekrun, K.; Khan, T.A.; Zhang, F.J.; Pasca, S.P.; Kay, M.A. Selection of rAAV vectors that cross the human blood-brain barrier and target the central nervous system using a transwell model. Mol. Ther.-Meth. Clin. D 2022, 27, 73–88. [Google Scholar] [CrossRef] [PubMed]
- Ricca, A.; Cascino, F.; Morena, F.; Martino, S.; Gritti, A. In vitro Validation of Chimeric beta-Galactosylceramidase Enzymes With Improved Enzymatic Activity and Increased Secretion. Front. Mol. Biosci. 2020, 7, 167. [Google Scholar] [CrossRef] [PubMed]
- Matthes, F.; Andersson, C.; Stein, A.; Eistrup, C.; Fogh, J.; Gieselmann, V.; Wenger, D.A.; Matzner, U. Enzyme replacement therapy of a novel humanized mouse model of globoid cell leukodystrophy. Exp. Neurol. 2015, 271, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Kofoed, R.H.; Noseworthy, K.; Wu, K.; Sivadas, S.; Stanek, L.; Elmer, B.; Hynynen, K.; Shihabuddin, L.S.; Aubert, I. The engineered AAV2-HBKO promotes non-invasive gene delivery to large brain regions beyond ultrasound targeted sites. Mol. Ther. Methods Clin. Dev. 2022, 27, 167–184. [Google Scholar] [CrossRef]
- Weber-Adrian, D.; Kofoed, R.H.; Silburt, J.; Noroozian, Z.; Shah, K.; Burgess, A.; Rideout, S.; Kugler, S.; Hynynen, K.; Aubert, I. Systemic AAV6-synapsin-GFP administration results in lower liver biodistribution, compared to AAV1&2 and AAV9, with neuronal expression following ultrasound-mediated brain delivery. Sci. Rep. 2021, 11, 1934. [Google Scholar] [CrossRef] [PubMed]
- Salegio, E.A.; Samaranch, L.; Kells, A.P.; Forsayeth, J.; Bankiewicz, K. Guided delivery of adeno-associated viral vectors into the primate brain. Adv. Drug Deliver. Rev. 2012, 64, 598–604. [Google Scholar] [CrossRef]
- Ohno, K.; Samaranch, L.; Hadaczek, P.; Bringas, J.R.; Allen, P.C.; Sudhakar, V.; Stockinger, D.E.; Snieckus, C.; Campagna, M.V.; San Sebastian, W.; et al. Kinetics and MR-Based Monitoring of AAV9 Vector Delivery into Cerebrospinal Fluid of Nonhuman Primates. Mol. Ther.-Meth. Clin. D 2019, 13, 47–54. [Google Scholar] [CrossRef]
- Hinderer, C.; Bell, P.; Katz, N.; Vite, C.H.; Louboutin, J.P.; Bote, E.; Yu, H.; Zhu, Y.; Casal, M.L.; Bagel, J.; et al. Evaluation of Intrathecal Routes of Administration for Adeno-Associated Viral Vectors in Large Animals. Hum. Gene Ther. 2018, 29, 15–24. [Google Scholar] [CrossRef]
- Kotterman, M.A.; Vazin, T.; Schaffer, D.V. Enhanced selective gene delivery to neural stem cells in vivo by an adeno-associated viral variant. Development 2015, 142, 1885–1892. [Google Scholar] [CrossRef]
- Rafi, M.A.; Rao, H.Z.; Luzi, P.; Curtis, M.T.; Wenger, D.A. Extended normal life after AAVrh10-mediated gene therapy in the mouse model of Krabbe disease. Mol. Ther. 2012, 20, 2031–2042. [Google Scholar] [CrossRef]
- Duan, F.; Huang, R.; Zhang, F.; Zhu, Y.; Wang, L.; Chen, X.; Bai, L.; Guo, W.; Chang, S.C.; Hu, X.; et al. Biphasic modulation of insulin signaling enables highly efficient hematopoietic differentiation from human pluripotent stem cells. Stem Cell Res. Ther. 2018, 9, 205. [Google Scholar] [CrossRef]
- Frati, G.; Luciani, M.; Meneghini, V.; De Cicco, S.; Stahlman, M.; Blomqvist, M.; Grossi, S.; Filocamo, M.; Morena, F.; Menegon, A.; et al. Human iPSC-based models highlight defective glial and neuronal differentiation from neural progenitor cells in metachromatic leukodystrophy. Cell Death Dis. 2018, 9, 698. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.K.; Gomes, K.N.; Parreira, R.C.; Scalzo, S.; Pinto, M.C.X.; Santiago, H.C.; Birbrair, A.; Sack, U.; Ulrich, H.; Resende, R.R. Mouse Neural Stem Cell Differentiation and Human Adipose Mesenchymal Stem Cell Transdifferentiation Into Neuron-and Oligodendrocyte-like Cells With Myelination Potential. Stem Cell Rev. Rep. 2022, 18, 732–751. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Z.Y.; Yang, G.H.; Wang, H.Y.; Liu, S.Y.; Zhang, Y.J.; Cai, Y.; Chen, F.; Dai, H.; Xiao, Y.; Cheng, M.B.; et al. Klf4 methylated by Prmt1 restrains the commitment of primitive endoderm. Nucleic Acids Res. 2022, 50, 2005–2018. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.M.; Sun, W.Z.; Fan, X.Z.; Xu, Y.L.; Cheng, M.B.; Zhang, Y. Methylation of C/EBPa by PRMT1 Inhibits Its Tumor-Suppressive Function in Breast Cancer. Cancer Res. 2019, 79, 2865–2877. [Google Scholar] [CrossRef] [PubMed]
- Lai, K.; Kaspar, B.K.; Gage, F.H.; Schaffer, D.V. Sonic hedgehog regulates adult neural progenitor proliferation in vitro and in vivo. Nat. Neurosci. 2003, 6, 21–27. [Google Scholar] [CrossRef]
- Kaspar, B.K.; Vissel, B.; Bengoechea, T.; Crone, S.; Randolph-Moore, L.; Muller, R.; Brandon, E.P.; Schaffer, D.; Verma, I.M.; Lee, K.F.; et al. Adeno-associated virus effectively mediates conditional gene modification in the brain. Proc. Natl. Acad. Sci. USA 2002, 99, 2320–2325. [Google Scholar] [CrossRef]
- Excoffon, K.J.; Koerber, J.T.; Dickey, D.D.; Murtha, M.; Keshavjee, S.; Kaspar, B.K.; Zabner, J.; Schaffer, D.V. Directed evolution of adeno-associated virus to an infectious respiratory virus. Proc. Natl. Acad. Sci. USA 2009, 106, 3865–3870. [Google Scholar] [CrossRef]
- Van Vliet, K.; Mohiuddin, Y.; McClung, S.; Blouin, V.; Rolling, F.; Moullier, P.; Agbandje-McKenna, M.; Snyder, R.O. Adeno-associated virus capsid serotype identification: Analytical methods development and application. J. Virol. Methods 2009, 159, 167–177. [Google Scholar] [CrossRef]
- Senis, E.; Mosteiro, L.; Wilkening, S.; Wiedtke, E.; Nowrouzi, A.; Afzal, S.; Fronza, R.; Landerer, H.; Abad, M.; Niopek, D.; et al. AAVvector-mediated in vivo reprogramming into pluripotency. Nat. Commun. 2018, 9, 2651. [Google Scholar] [CrossRef]
- Kremer, L.P.M.; Cerrizuela, S.; Dehler, S.; Stiehl, T.; Weinmann, J.; Abendroth, H.; Kleber, S.; Laure, A.; El Andari, J.; Anders, S.; et al. High throughput screening of novel AAV capsids identifies variants for transduction of adult NSCs within the subventricular zone. Mol. Ther. Methods Clin. Dev. 2021, 23, 33–50. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.B.; Zhang, Y.; Cao, C.Y.; Zhang, W.L.; Zhang, Y.; Shen, Y.F. Specific phosphorylation of histone demethylase KDM3A determines target gene expression in response to heat shock. PLoS Biol. 2014, 12, e1002026. [Google Scholar] [CrossRef] [PubMed]
- Zanfini, A.; Dreassi, E.; Berardi, A.; Governini, L.; Corbini, G.; Costantino-Ceccarini, E.; Balestri, P.; Luddi, A. Quantification of psychosine in the serum of twitcher mouse by LC-ESI-tandem-MS analysis. J. Pharm. Biomed. Anal. 2013, 80, 44–49. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, G.; Cao, C.; Li, S.; Wang, W.; Zhang, Y.; Lv, Y. rAAV2-Mediated Restoration of GALC in Neural Stem Cells from Krabbe Patient-Derived iPSCs. Pharmaceuticals 2023, 16, 624. https://doi.org/10.3390/ph16040624
Tian G, Cao C, Li S, Wang W, Zhang Y, Lv Y. rAAV2-Mediated Restoration of GALC in Neural Stem Cells from Krabbe Patient-Derived iPSCs. Pharmaceuticals. 2023; 16(4):624. https://doi.org/10.3390/ph16040624
Chicago/Turabian StyleTian, Guoshuai, Chunyu Cao, Shuyue Li, Wei Wang, Ye Zhang, and Yafeng Lv. 2023. "rAAV2-Mediated Restoration of GALC in Neural Stem Cells from Krabbe Patient-Derived iPSCs" Pharmaceuticals 16, no. 4: 624. https://doi.org/10.3390/ph16040624
APA StyleTian, G., Cao, C., Li, S., Wang, W., Zhang, Y., & Lv, Y. (2023). rAAV2-Mediated Restoration of GALC in Neural Stem Cells from Krabbe Patient-Derived iPSCs. Pharmaceuticals, 16(4), 624. https://doi.org/10.3390/ph16040624