
Synergistic Effects of Caffeine in Combination with Conventional Drugs: Perspectives of a Drug That Never Ages

 ,
,  ,
,  and
and 
Abstract
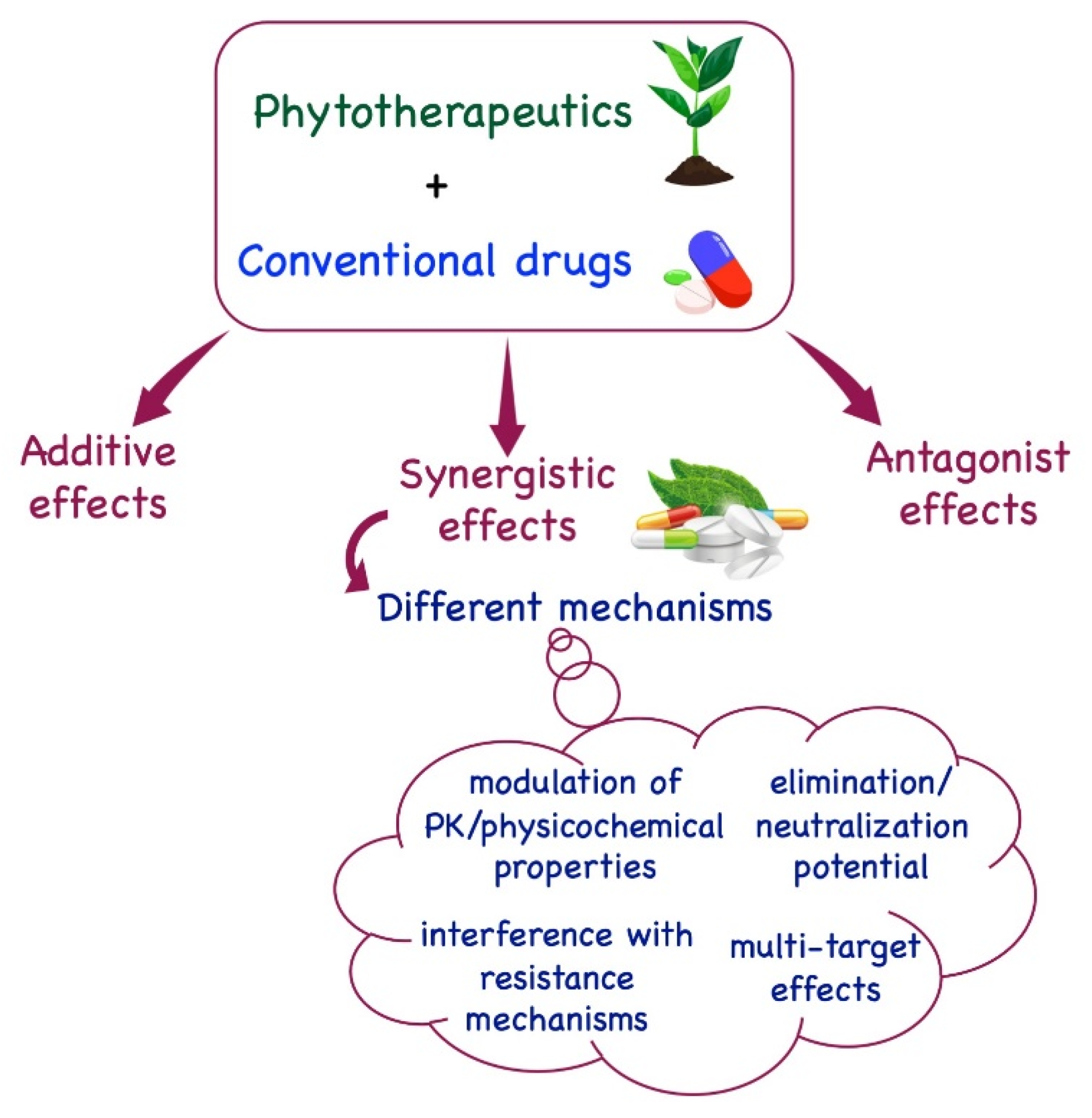
:1. Introduction
2. Biological Activities of Caffeine
2.1. Molecular Activities
2.2. Pharmacological Effects
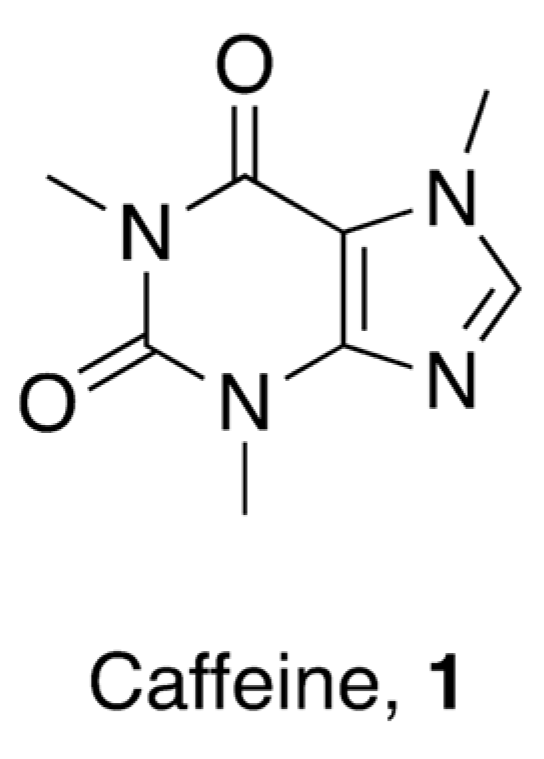
3. Caffeine and Antimicrobial Drugs
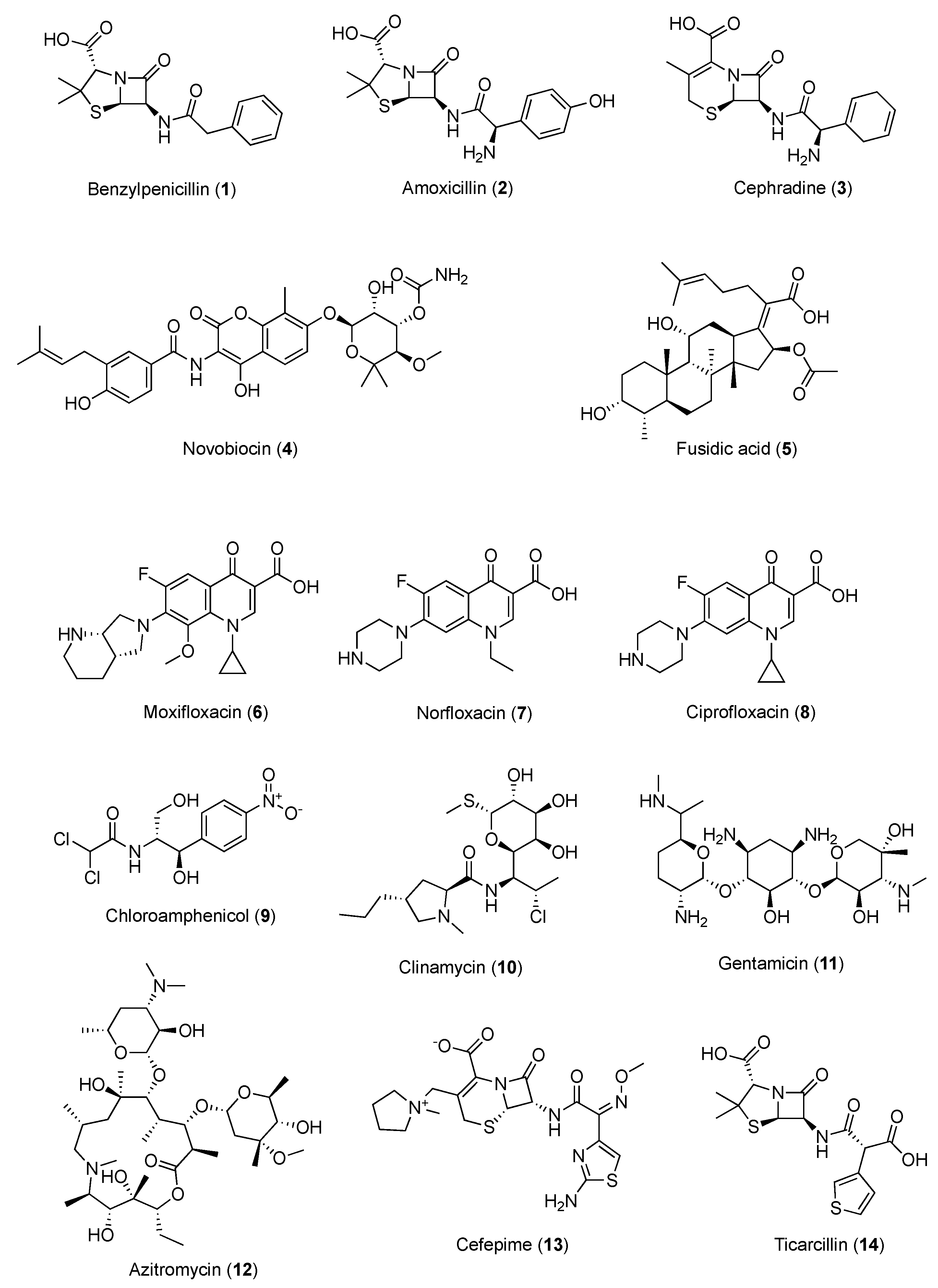
3.1. Caffeine and Antibacterial Agents
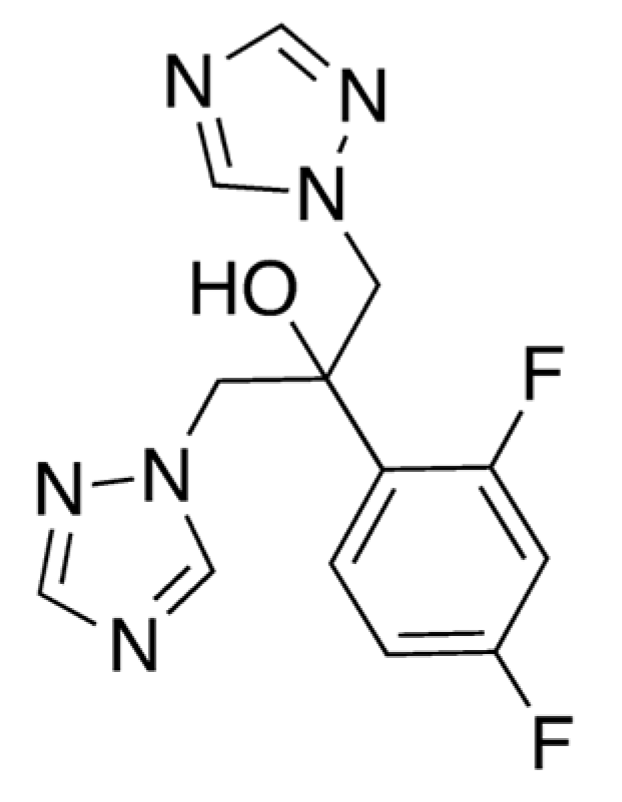
3.2. Caffeine and Fluconazole
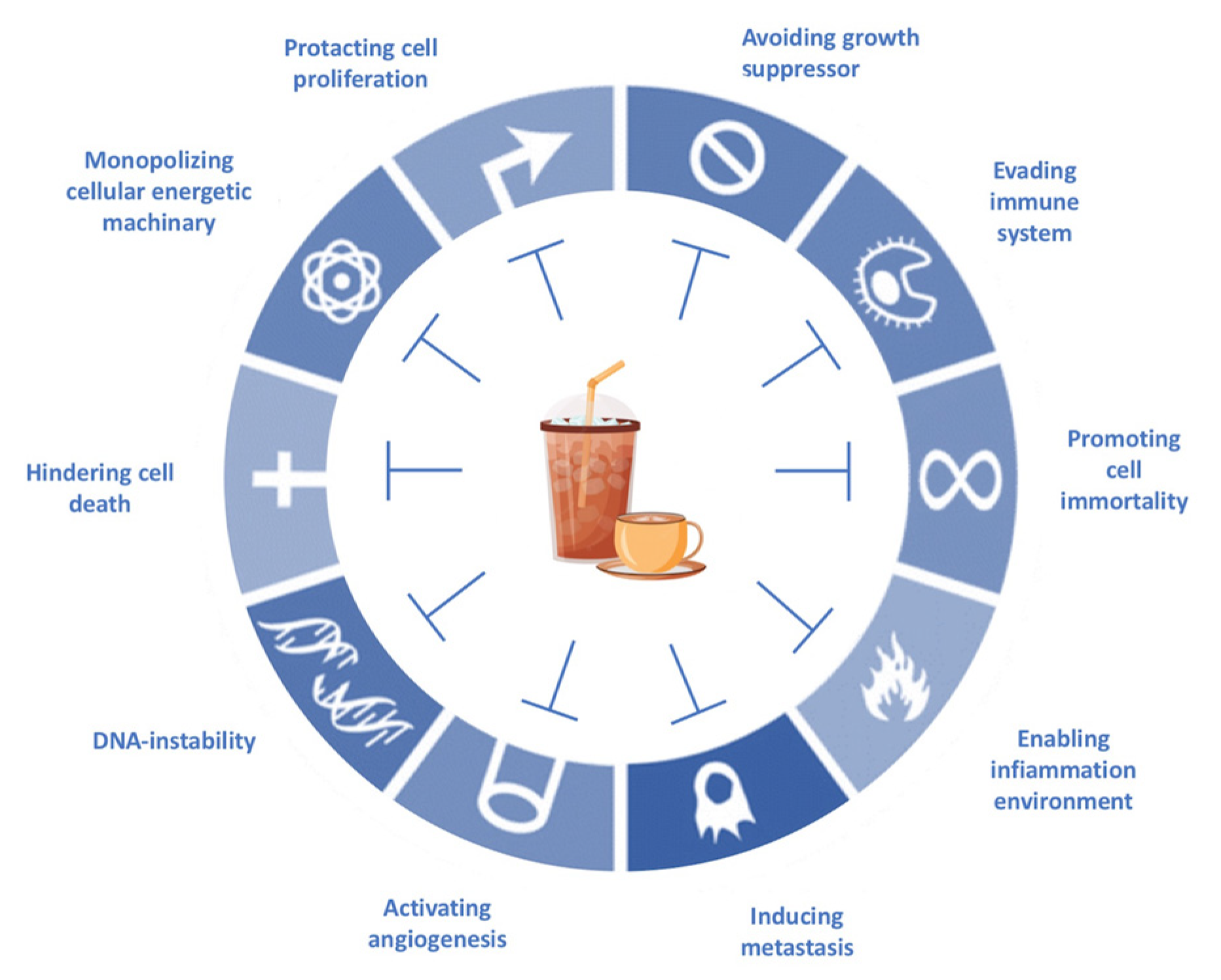
4. Caffeine and Anticancer Agents
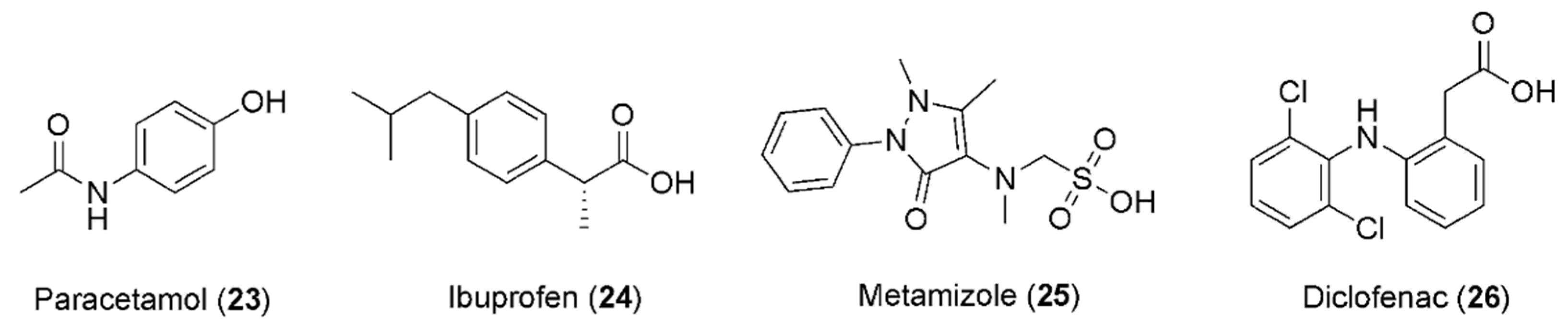
5. Caffeine and Anti-Inflammatory Drugs
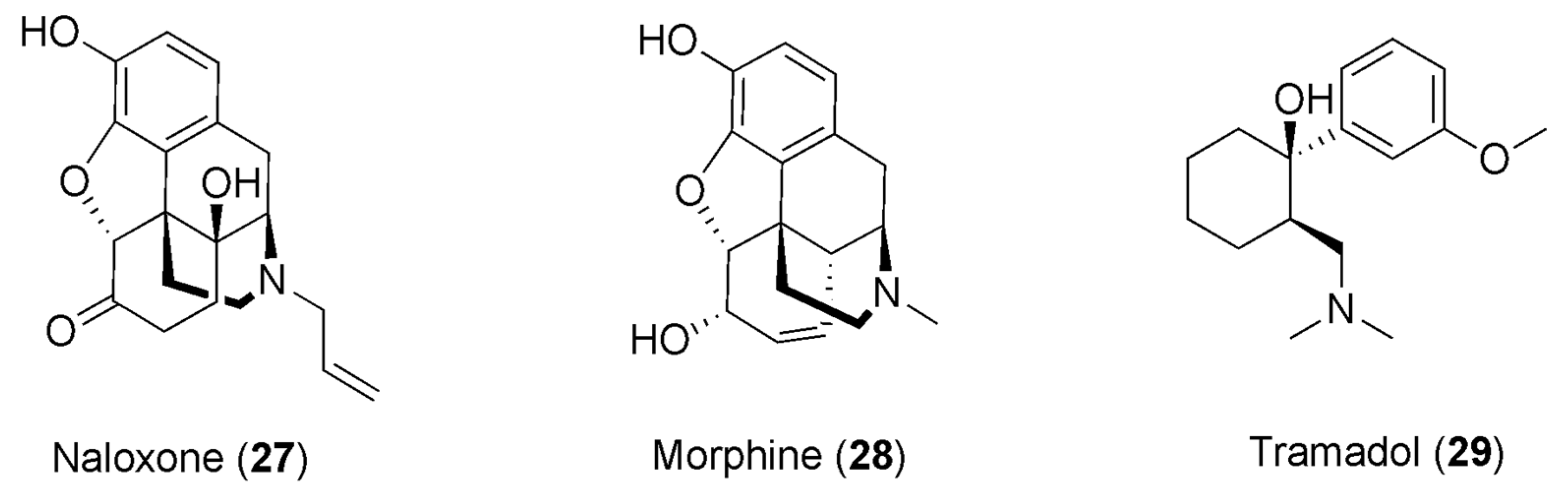
6. Caffeine and Opioids
7. Synergistic Effects of Caffeine in the Treatment of Some Diseases
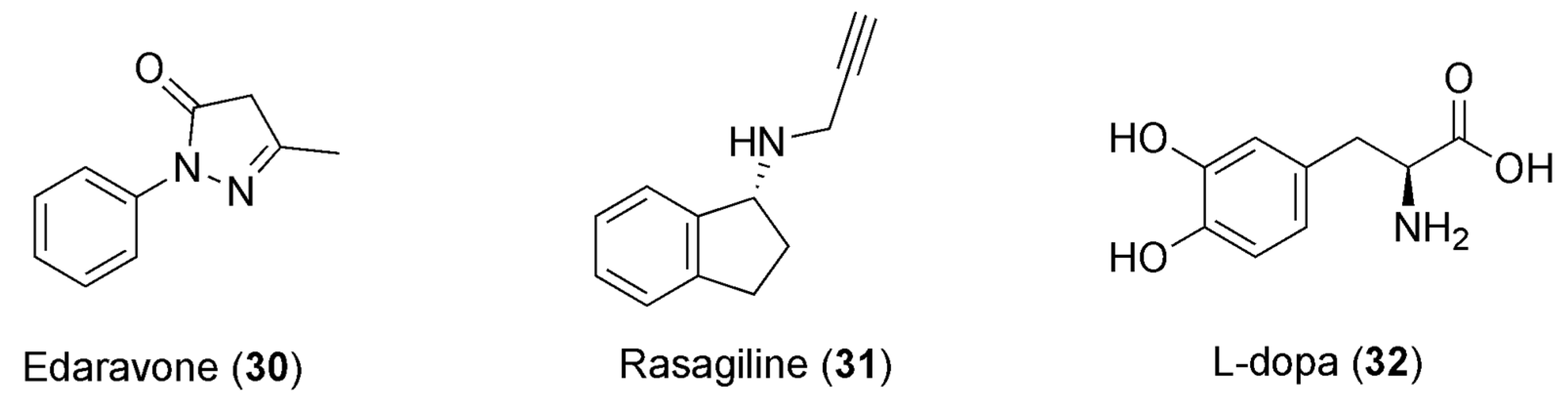
7.1. Caffeine and Parkinson’s Disease
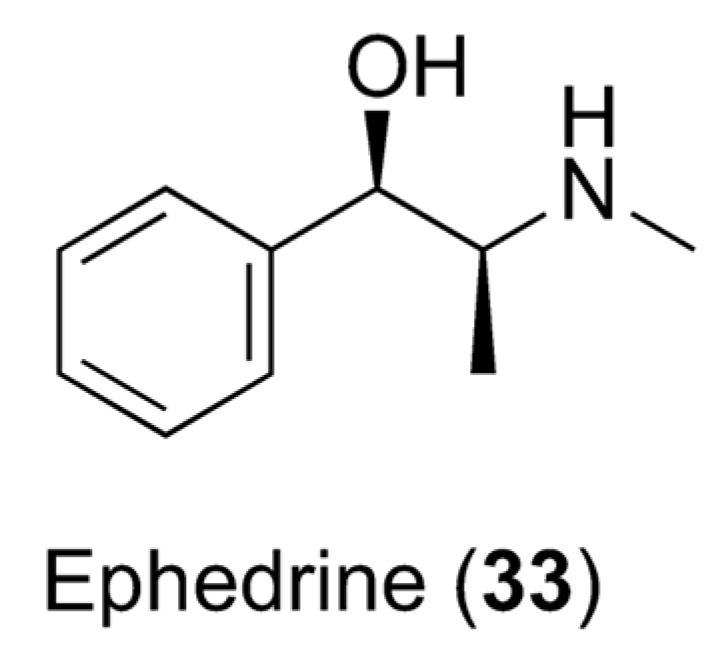
7.2. Caffeine and Obesity
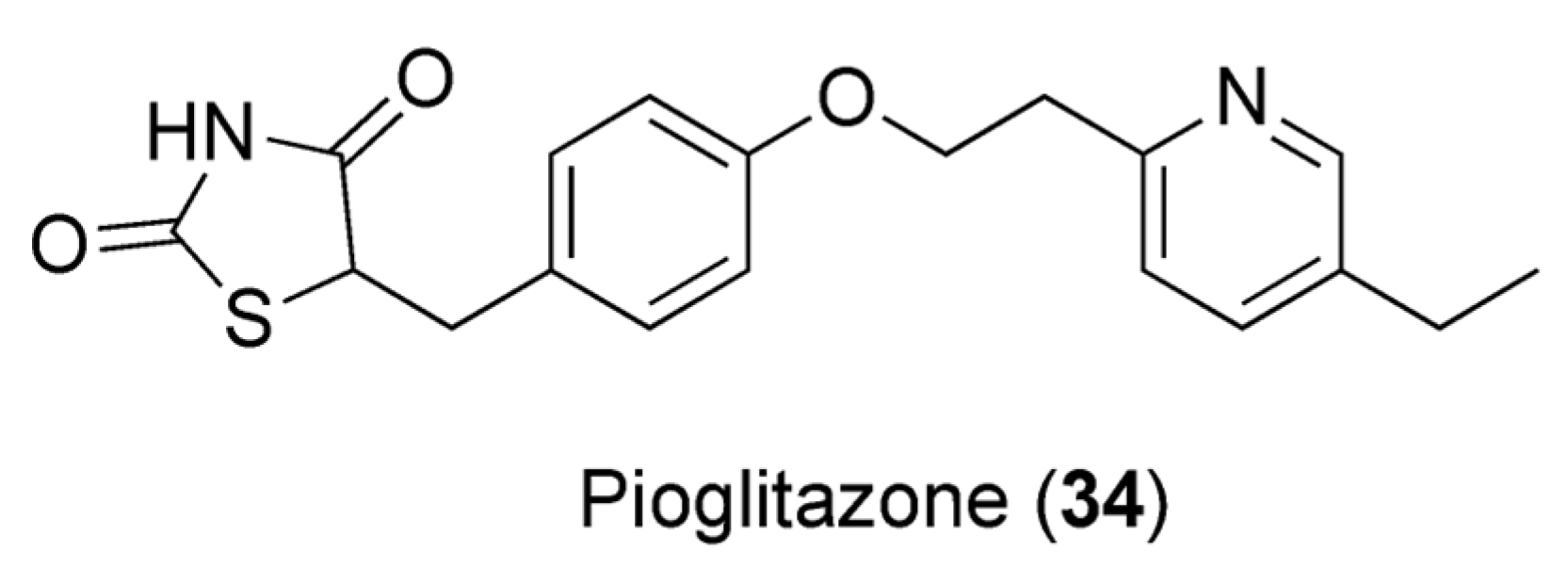
7.3. Caffeine and Pioglitazone
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Doos, L.; Roberts, E.O.; Corp, N.; Kadam, U.T. Multi-drug therapy in chronic condition multimorbidity: A systematic review. Fam. Pract. 2014, 31, 654–663. [Google Scholar] [CrossRef] [PubMed]
- Pezzani, R.; Salehi, B.; Vitalini, S.; Iriti, M.; Zuñiga, F.A.; Sharifi-Rad, J.; Martorell, M.; Martins, N. Synergistic effects of plant derivatives and conventional chemotherapeutic agents: An update on the cancer perspective. Medicina 2019, 55, 110. [Google Scholar] [CrossRef] [PubMed]
- Foucquier, J.; Guedj, M. Analysis of drug combinations: Current methodological landscape. Pharmacol. Res. Perspect. 2015, 3, e00149, Erratum in Pharmacol. Res. Perspect. 2019, 7, e00549. [Google Scholar] [CrossRef] [PubMed]
- Obodozie-Ofoegbu, O. Pharmacokinetics and Drug Interactions of Herbal Medicines: A Missing Critical Step in the Phytomedicine/Drug Development Process. In Readings in Advanced Pharmacokinetics—Theory, Methods and Applications; IntechOpen: London, UK, 2012. [Google Scholar]
- Dzobo, K. The Role of natural products as sources of therapeutic agents for innovative drug discovery. Compr. Pharmacol. 2022, 2, 408–422. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the 30 years from 1981 to 2010. J. Nat. Prod. 2012, 75, 311–335. [Google Scholar] [CrossRef] [PubMed]
- Cragg, G.M.; Grothaus, P.G.; Newman, D.J. Impact of natural products on developing new anti-cancer agents. Chem. Rev. 2009, 109, 3012–3043. [Google Scholar] [CrossRef]
- Berenbaum, M.C. What is synergy? Pharmacol. Rev. 1989, 41, 93–141. [Google Scholar]
- Silva, D.M.; Costa, P.A.D.; Ribon, A.O.B.; Purgato, G.A.; Gaspar, D.M.; Diaz, M.A.N. Plant extracts display synergism with different classes of antibiotics. An. Acad. Bras. Cienc. 2019, 91, e20180117. [Google Scholar] [CrossRef]
- Pöch, G. Synergism/potentiation and antagonism—phenomena and mechanisms. In Combined Effects of Drugs and Toxic Agents; Springer: Vienna, Austria, 1993. [Google Scholar]
- Veldstra, H. Synergism and potentiation with special reference to the combination of structural analogues. Pharmacol Rev. 1956, 8, 339–387. [Google Scholar]
- Imming, P.; Sinning, C.; Meyer, A. Drugs, their targets and the nature and number of drug targets. Nat. Rev. Drug Discov. 2006, 5, 821–834. [Google Scholar] [CrossRef]
- Butterweck, V.; Jurgenliemk, G.; Nahrstedt, A.; Winterho, H. Flavonoids from Hypericum perforatum show antidepressant activity in the forced swimming test. Planta Med. 2000, 66, 3–6. [Google Scholar] [CrossRef]
- Hemaiswarya, S.; Kruthiventi, A.K.; Doble, M. Synergism between natural products and antibiotics against infectious diseases. Phytomedicine 2008, 15, 639–652. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.H.; Cheng, Y.C. Old formula, new rx: The journey of phy906 as cancer adjuvant therapy. J. Ethnopharmacol. 2012, 140, 614–623. [Google Scholar] [CrossRef] [PubMed]
- Rasool, M.; Iqbal, J.; Malik, A.; Ramzan, H.S.; Qureshi, M.S.; Asif, M.; Qazi, M.H.; Kamal, M.A.; Chaudhary, A.G.; Al-Qahtani, M.H.; et al. Hepatoprotective effects of Silybum marianum (silymarin) and Glycyrrhiza glabra (glycyrrhizin) in combination: A possible synergy. Evid. Based Complement. Alternat. Med. 2014, 2014, 641597. [Google Scholar] [CrossRef]
- Betoni, J.E.; Mantovani, R.P.; Barbosa, L.N.; Di Stasi, L.C.; Fernandes Junior, A. Synergism between plant extract and antimicrobial drugs used on Staphylococcus aureus diseases. Mem. Ist. Oswaldo Cruz 2006, 101, 387–390. [Google Scholar] [CrossRef] [PubMed]
- Darwish, R.M.; Aburjai, T.; Al-Khalil, S.; Mahafzah, A. Screening of antibiotic resistant inhibitors from local plant materials against two different strains of Staphylococcus aureus. J. Ethnopharmacol. 2002, 79, 359–364. [Google Scholar] [CrossRef]
- Palaniappan, K.; Holley, R.A. Use of natural antimicrobials to increase antibiotic susceptibility of drug resistant bacteria. Int. J. Food Microbiol. 2010, 140, 164–168. [Google Scholar] [CrossRef]
- Nascimento, G.G.; Locatelli, J.; Freitas, P.C.; Silva, G.L. Antibacterial activity of plant extracts and phytochemicals on antibiotic-resistant bacteria. Braz. J. Microbiol. 2000, 31, 247–256. [Google Scholar] [CrossRef]
- Wink, M. Evolutionary advantage and molecular modes of action of multi-component mixtures used in phytomedicine. Curr. Drug Metab. 2008, 9, 996–1009. [Google Scholar] [CrossRef]
- Wink, M. Modes of action of herbal medicines and plant secondary metabolites. Medicines 2015, 2, 251–286. [Google Scholar] [CrossRef]
- Izzo, A.A. Interactions between herbs and conventional drugs: Overview of the clinical data. Med. Princip. Pract. 2012, 21, 404–428. [Google Scholar] [CrossRef]
- Gupta, D.; Lis, C.G.; Birdsall, T.C.; Grutsch, J.F. The use of dietary supplements in a community hospital comprehensive cancer center: Implications for conventional cancer care. Support. Care Cancer 2005, 13, 912–919. [Google Scholar] [CrossRef]
- Diener, H.C.; Pfaffenrath, V.; Pageler, L.; Peil, H.; Aicher, B. The fixed combination of acetylsalicylic acid, paracetamol and caffeine is more effective than single substances and dual combination for the treatment of headache: A multicentre, randomized, double-blind, single-dose, placebo-controlled parallel group study. Cephalalgia 2005, 25, 776–787. [Google Scholar] [CrossRef]
- Faudone, G.; Arifi, S.; Merk, D. The medicinal chemistry of caffeine. J. Med. Chem. 2021, 64, 7156–7178. [Google Scholar] [CrossRef] [PubMed]
- Nehlig, A. Interindividual differences in caffeine metabolism and factors driving caffeine consumption. Pharmacol. Rev. 2018, 70, 384–411. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 1997, 23, 3–25. [Google Scholar] [CrossRef]
- Wilkinson, S.C.; Maas, W.J.M.; Nielsen, J.B.; Greaves, L.C.; van de Sandt, J.J.M.; Williams, F.M. Interactions of skin thickness and physicochemical properties of test compounds in percutaneous penetration studies. Int. Arch. Occup. Environ. Health 2006, 79, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, K.A.; Gao, Z.G.; Matricon, P.; Eddy, M.T.; Carlsson, J. Adenosine A2A receptor antagonists: From caffeine to selective non-xanthines. Br. J. Pharmacol. 2022, 79, 3496–3511. [Google Scholar] [CrossRef] [PubMed]
- Francis, S.H.; Sekhar, K.R.; Ke, H.; Corbin, J.D. Inhibition of cyclic nucleotide phosphodiesterases by methylxanthines and related compounds. Handb. Exp. Pharmacol. 2011, 200, 93–133. [Google Scholar] [CrossRef]
- Shi, D.; Padgett, W.L.; Daly, J.W. Caffeine analogs: Effects on ryanodine-sensitive calcium-release channels and GABA-A receptors. Cell. Mol. Neurobiol. 2003, 23, 331–347. [Google Scholar] [CrossRef]
- Fabiani, C.; Murray, A.P.; Corradi, J.; Antollini, S.S. A novel pharmacological activity of caffeine in the cholinergic system. Neuropharmacology 2018, 135, 464–473. [Google Scholar] [CrossRef]
- McPhersonx, P.S.; Kim, Y.K.; Valdivia, H.; Knudson, C.M.; Takekura, H.; Franzini-Armstrong, C.; Coronadot, R.; Campbell, K.P. The brain ryanodine receptor: A caffeine-sensitive calcium release channel. Neuron 1991, 7, 17–25. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, R.S. Caffeine. In Reproductive and Developmental Toxicology; Gupta, R.C., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2011; pp. 355–364. [Google Scholar]
- Alfaro, T.M.; Monteiro, R.A.; Cunha, R.A.; Cordeiro, C.R. Chronic Coffee consumption and respiratory disease: A systematic review. Clin. Respir. J. 2018, 12, 1283–1294. [Google Scholar] [CrossRef] [PubMed]
- Henderson-Smart, D.J.; De Paoli, A.G.; Haughton, D. Methylxanthine treatment for apnoea in preterm infants. Cochrane Database Syst. Rev. 2010, CD000140. [Google Scholar] [CrossRef] [PubMed]
- Eichenwald, E.C. Apnea of Prematurity. Pediatrics 2016, 137, e20153757. [Google Scholar] [CrossRef] [PubMed]
- Lipton, R.B.; Diener, H.C.; Robbins, M.S.; Garas, S.Y.; Patel, K. Caffeine in the management of patients with headache. J. Headache Pain 2017, 18, 107. [Google Scholar] [CrossRef]
- Sawynok, J. Caffeine and Pain. Pain 2011, 152, 726–729. [Google Scholar] [CrossRef]
- Doepker, C.; Lieberman, H.R.; Smith, A.P.; Peck, J.D.; El-Sohemy, A.; Welsh, B.T. Caffeine: Friend or Foe? Annu. Rev. Food Sci. Technol. 2016, 7, 117–137. [Google Scholar] [CrossRef]
- Weiser, T.; Weigmann, H. Effect of caffeine on the bioavailability and pharmacokinetics of an acetylsalicylic acid-paracetamol combination: Results of a phase I study. Adv. Ther. 2019, 36, 597–607. [Google Scholar] [CrossRef]
- Silva, A.C.; Lemos, C.; Gonçalves, F.Q.; Pliássova, A.V.; Machado, N.J.; Silva, H.B.; Canas, P.M.; Cunha, R.A.; Lopes, J.P.; Agostinho, P. Blockade of adenosine A2A receptors recovers early deficits of memory and plasticity in the triple transgenic mouse model of Alzheimer’s disease. Neurobiol. Dis. 2018, 117, 72–81. [Google Scholar] [CrossRef]
- Shapiro, R.E. Caffeine and headaches. Curr. Pain Headache Rep. 2008, 12, 311–315. [Google Scholar] [CrossRef]
- Van Gelder, B.M.; Buijsse, B.; Tijhuis, M.; Kalmijn, S.; Giampaoli, S.; Nissinen, A.; Kromhout, D. Coffee consumption is inversely associated with cognitive decline in elderly european men: The FINE study. Eur. J. Clin. Nutr. 2007, 61, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Arendash, G.W.; Schleif, W.; Rezai-Zadeh, K.; Jackson, E.K.; Zacharia, L.C.; Cracchiolo, J.R.; Shippy, D.; Tan, J. Caffeine protects Alzheimer’s mice against cognitive impairment and reduces brain β-amyloid production. Neuroscience 2006, 142, 941–952. [Google Scholar] [CrossRef] [PubMed]
- Arendash, G.W.; Mori, T.; Cao, C.; Mamcarz, M.; Runfeldt, M.; Dickson, A.; Rezai-Zadeh, K.; Tan, J.; Citron, B.A.; Lin, X.; et al. Caffeine reverses cognitive impairment and decreases brain amyloid-β levels in aged Alzheimer’s disease mice. J. Alzheimers Dis. 2009, 17, 661–680. [Google Scholar] [CrossRef] [PubMed]
- Dall’Igna, O.P.; Fett, P.; Gomes, M.W.; Souza, D.O.; Cunha, R.A.; Lara, D.R. Caffeine and adenosine A2a receptor antagonists prevent β-amyloid (25−35)-induced cognitive deficits in mice. Exp. Neurol. 2007, 203, 241–245. [Google Scholar] [CrossRef]
- Wostyn, P.; Van Dam, D.; Audenaert, K.; De Deyn, P.P. increased cerebrospinal fluid production as a possible mechanism underlying caffeine’s protective effect against Alzheimer’s disease. Int. J. Alzheimer’s Dis. 2011, 2011, 617420. [Google Scholar] [CrossRef]
- Laurent, C.; Burnouf, S.; Ferry, B.; Batalha, V.L.; Coelho, J.E.; Baqi, Y.; Malik, E.; Mariciniak, E.; Parrot, S.; Van Der Jeugd, A.; et al. A2A adenosine receptor deletion is protective in a mouse model of tauopathy. Mol. Psychiatry 2016, 21, 97–107. [Google Scholar] [CrossRef]
- Ferri, M.; Ranucci, E.; Romagnoli, P.; Giaccone, V. Antimicrobial resistance: A global emerging threat to public health systems. Crit. Rev. Food Sci. Nutr. 2017, 57, 2857–2876. [Google Scholar] [CrossRef]
- Robinson, T.P.; Wertheim, H.F.L.; Kakkar, M.; Kariuki, S.; Bu, D.; Price, L.B. Animal production and antimicrobial resistance in the clinic. Lancet 2016, 387, e1–e3. [Google Scholar] [CrossRef]
- Durão, P.; Balbontín, R.; Gordo, I. Evolutionary mechanisms shaping the maintenance of antibiotic resistance. Trends Microbiol. 2018, 26, 677–691. [Google Scholar] [CrossRef]
- Esimone, C.O.; Okoye, F.B.C.; Nworu, C.S.; Agubata, C.O. In vitro interaction between caffeine and some penicillin antibiotics against Staphylococcus aureus. Trop. J. Pharm. Res. 2008, 7, 969–974. [Google Scholar] [CrossRef]
- Roberts, J.J. Mechanism of Potentiation by Caffeine of Genotoxic Damage Induced by Physical and Chemical Agents: Possible Relevance to Carcinogenesis. In Caffeine; Dews, P.B., Ed.; Springer: Berlin/Heidelberg, Germany, 1984. [Google Scholar] [CrossRef]
- Woziwodzka, A.; Krychowiak-Ma’snicka, M.; Gołu´nski, G.; Łosiewska, A.; Borowik, A.; Wyrzykowski, D.; Piosik, J. New life of an old drug: Caffeine as a modulator of antibacterial activity of commonly used antibiotics. Pharmaceuticals 2022, 15, 872. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, S.A.; Salameh, M.M.; Phetsomphou, S.; Yang, H.; Seo, C.W. Application of caffeine, 1,3,7-trimethylxanthine, to control Escherichia coli O157:H7. Food Chem. 2006, 99, 645–650. [Google Scholar] [CrossRef]
- Kang, T.M.; Yuan, J.; Nguyen, A.; Becket, E.; Yang, H.; Miller, J.H. The aminoglycoside antibiotic kanamycin damages DNA bases in Escherichia coli: Caffeine potentiates the DNA-damaging effects of kanamycin while suppressing cell killing by ciprofloxacin in Escherichia coli and Bacillus anthracis. Antimicrob. Agents Chemother. 2012, 56, 3216–3223. [Google Scholar] [CrossRef]
- Woziwodzka, A.; Gołu´nski, G.; Wyrzykowski, D.; Ka´zmierkiewicz, R.; Piosik, J. Caffeine and other methylxanthines as interceptors of food-borne aromatic mutagens: Inhibition of Trp-P-1 and Trp-P-2 mutagenic activity. Chem. Res. Toxicol. 2013, 26, 1660–1673. [Google Scholar] [CrossRef] [PubMed]
- Gołu´nski, G.; Woziwodzka, A.; Iermak, I.; Rychłowski, M.; Piosik, J. Modulation of acridine mutagen ICR191 intercalation to DNA by methylxanthines-analysis with mathematical models. Bioorg. Med. Chem. 2013, 21, 3280–3289. [Google Scholar] [CrossRef]
- Ulanowska, K.; Piosik, J.; Gwizdek-Wi´sniewska, A.; Wegrzyn, G. Formation of stacking complexes between caffeine (1,2,3-trimethylxanthine) and 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine may attenuate biological effects of this neurotoxin. Bioorg. Chem. 2005, 33, 402–413. [Google Scholar] [CrossRef]
- Anttila, V.J.; Ruutu, P.; Bondestam, S.; Jansson, S.E.; Nordling, S.; Farkkila, M.; Sivonen, A.; Castren, M.; Ruutu, T. Hepatosplenic yeast infection in patients with acute leukemia: A diagnostic problem. Clin. Infect. Dis. 1994, 18, 979–981. [Google Scholar] [CrossRef]
- Bossche, H.V. Biochemical targets for antifungal azole derivatives in: Current topics in medical mycology: Hypothesis on the mode of action. Springer 1985, 1, 313–351. [Google Scholar] [CrossRef]
- Okubo, S.; Toda, M.; Hara, Y.; Shimamura, T. Antifungal and fungicidal activities of tea extract and catechin against Trichophyton. Nihon Saikingaku Zasshi 1991, 46, 509–514. [Google Scholar] [CrossRef]
- Reinke, A.; Chen, J.C.; Aronova, S.; Powers, T. Caffeine targets TOR complex I and provides evidence for a regulatory link between the FRB and kinase domains of Tor1p. J. Biol. Chem. 2006, 281, 31616–31626. [Google Scholar] [CrossRef] [PubMed]
- Gaascht, F.; Dicato, M.; Diederich, M. Coffee provides a natural multitarget pharmacopeia against the hallmarks of cancer. Genes Nutr. 2015, 6, 51. [Google Scholar] [CrossRef] [PubMed]
- Kang, N.J.; Lee, K.W.; Kim, B.H.; Bode, A.M.; Lee, H.J.; Heo, Y.S.; Boardman, L.; Limburg, P.; Lee, H.J.; Dong, Z. Coffee phenolic phytochemicals suppress colon cancer metastasis by targeting MEK and TOPK. Carcinogenesis 2011, 6, 921–928. [Google Scholar] [CrossRef] [PubMed]
- Nakata, H.; Miyazaki, T.; Iwasaki, T.; Nakamura, A.; Kidani, T.; Sakayama, K.; Masumoto, J.; Miura, H. The development of tumor-specific caffeine-potentiated chemotherapy using a novel drug delivery system with Span 80 nano-vesicles. Oncology Rep. 2015, 4, 1593–1598. [Google Scholar] [CrossRef]
- Chung, F.L.; Wang, M.; Rivenson, A.; Iatropoulos, M.J.; Reinhardt, J.C.; Pittman, B.; Ho, C.T.; Amin, S.G. Inhibition of lung carcinogenesis by black tea in fischer rats treated with a tobacco-specific carcinogen: Caffeine as an important constituent. Cancer Res. 1998, 58, 4096–4101. [Google Scholar]
- Motegi, T.; Katayama, M.; Uzuka, Y.; Okamura, Y. Evaluation of anticancer effects and enhanced doxorubicin cytotoxicity of xanthine derivatives using canine hemangiosarcoma cell lines. Res.Vet. Sci. 2013, 2, 600–605. [Google Scholar] [CrossRef]
- Mohammadrezaei, F.M.; Movaghar, A.F.; Gharghabi, M. The Effect of caffeine and chk2 inhibitor on doxorubicin-induced cellular senescence in MCF-7 cells. Drug Res. 2016, 9, 450–454. [Google Scholar] [CrossRef]
- Tsuchiya, H.; Tomita, K.; Yamamoto, N.; Mori, Y.; Asada, N. Caffeine-potentiated chemotherapy and conservative surgery for high-grade soft-tissue sarcoma. Anticancer Res. 1998, 5, 3651–3656. [Google Scholar]
- Tomita, K.; Tsuchiya, H. Enhancement of cytocidal and antitumor effect of cisplatin by caffeine in human osteosarcoma. Clin. Ther. 1989, 1, 43–52. [Google Scholar]
- Karita, M.; Tsuchiya, H.; Kawahara, M.; Kasaoka, S.; Tomita, K. The antitumor effect of liposome-encapsulated cisplatin on rat osteosarcoma and its enhancement by caffeine. Anticancer Res. 2008, 28, 1449–1457. [Google Scholar]
- Tsabar, M.; Mason, J.M.; Chan, Y.L.; Bishop, D.K.; Haber, J.E. Caffeine inhibits gene conversion by displacing Rad51 from ssDNA. Nucleic Acids Res. 2015, 43, 6902–6918. [Google Scholar] [CrossRef] [PubMed]
- Oda, Y.; Hidaka, M.; Suzuki, A. Caffeine has a synergistic anticancer effect with cisplatin via inhibiting fanconi anemia group D2 protein monoubiquitination in hepatocellular carcinoma cells. Biol. Pharm. Bull. 2017, 40, 2005–2009. [Google Scholar] [CrossRef]
- Hashimoto, T.; He, Z.; Ma, W.Y.; Schmid, P.C.; Bode, A.M.; Yang, C.S.; Dong, Z. Caffeine inhibits cell proliferation by G0/G1 phase arrest in JB6 cells. Cancer Res. 2004, 64, 3344–3349. [Google Scholar] [CrossRef] [PubMed]
- Abe, K.; Yamamoto, N.; Hayashi, K.; Takeuchi, A.; Tsuchiya, H. Caffeine citrate enhanced cisplatin antitumor effects in osteosarcoma and fibrosarcoma in vitro and in vivo. BMC Cancer 2019, 19, 689. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, K.; Yamamoto, N.; Hayashi, K.; Takeuchi, A.; Kimura, H.; Miwa, S.; Hoffman, R.M.; Tsuchiya, H. Non-toxic efficacy of the combination of caffeine and valproic acid on human osteosarcoma cells in vitro and in orthotopic nude-mouse models. Anticancer Res. 2016, 36, 4477–4482. [Google Scholar] [CrossRef]
- Gururajanna, B.; Al-Katib, A.A.; Li, Y.W.; Aranha, O.; Vaitkevicius, V.K.; Sarkar, F.H. Molecular effects of Taxol and caffeine on pancreatic cancer cells. Int. J. Mol. Med. 1999, 5, 501–507. [Google Scholar] [CrossRef]
- Popović, D.J.; Lalošević, D.; Miljković, D.; Popović, K.J.; Čapo, I.; Popović, J.K. Caffeine induces metformin anticancer effect on fibrosarcoma in hamsters. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 2461–2467. [Google Scholar] [CrossRef]
- Bruno, S.; Ledda, B.; Tenca, C.; Ravera, S.; Orengo, A.M.; Mazzarello, A.N.; Pesenti, E.; Casciaro, S.; Racchi, O.; Ghiotto, F.; et al. Metformin inhibits cell cycle progression of B-cell chronic lymphocytic leukemia cells. Oncotarget 2015, 6, 22624–22640. [Google Scholar] [CrossRef]
- Hwang, Y.P.; Jeong, H.G. Metformin blocks migration and invasion of tumour cells by inhibition of matrix metalloproteinase-9 activation through a calcium and protein kinase Cα-dependent pathway: Phorbol-12-myristate-13-acetate-induced/extracellular signal-regulated kinase/activator protein-1. Br. J. Pharmacol. 2010, 160, 1195–1211. [Google Scholar] [CrossRef]
- Wang, Z.; Gu, C.; Wang, X.; Lang, Y.; Wu, Y.; Wu, X.; Zhu, X.; Wang, K.; Yang, H. Caffeine enhances the anti-tumor effect of 5-fluorouracil via increasing the production of reactive oxygen species in hepatocellular carcinoma. Med. Oncol. 2019, 36, 97. [Google Scholar] [CrossRef]
- Granados-Soto, V.; Castañeda-Hernández, G. A review of the pharmacokinetic and pharmacodynamic factors in the potentiation of the antinociceptive effect of nonsteroidal anti-infammatory drugs by caffeine. J. Pharm. Toxicol. 1999, 42, 67–72. [Google Scholar] [CrossRef]
- Granados-Soto, V.; López-Muñoz, F.J.; Castañeda-Hernández, G.; Salazar, L.A.; Villarreal, J.E.; Flores-Murrieta, F.J. Characterization of the analgesic effect of paracetamol and caffeine combinations in the pain-induced functional impairment model in the rat. J. Pharm. Pharmacol. 1993, 45, 627–631. [Google Scholar] [CrossRef]
- Sawynok, J. Adenosine receptor activation and nociception. Eur. J. Pharmacol. 1998, 317, 1–11. [Google Scholar] [CrossRef]
- Polski, A.; Kasperek, R.; Sobotka-Polska, K.; Poleszak, E. Review on analgesic effect of co-administrated ibuprofen and caffeine. Curr. Issues Pharm. Med. Sci. 2014, 27, 10–13. [Google Scholar] [CrossRef]
- Diaz-Reval, M.I.; Galvan-Orozco, R.; Lopez-Munoz, F.J.; Carrillo-Munguia, N. Sinergismo de la cafeína sobre los efectos antinociceptivos del metamizol. Cir. Ciruj. 2008, 76, 241–246. [Google Scholar]
- Sawynok, J.; Reid, A. Caffeine antinociception: Role of formalin concentration and adenosine A1 and A2 receptors. Eur. J. Pharmacol. 1996, 298, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Flores-Ramos, J.M.; Diaz-Reval, M.I. Opioid mechanism involvement in the synergism produced by the combination of diclofenac and caffeine in the formalin model. ISRN Pain 2013, 2013, 196429. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.; Dalziel, S. Antenatal corticosteroids for accelerating fetal lung maturation for women at risk of preterm birth. Cochrane Database Syst. Rev. 2017, 3, CD004454. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, B.; Roberts, R.; Millar, D.; Kirpalani, H. Evidence-based neonatal drug therapy for prevention of bronchopulmonary dysplasia in very-low-birth- weight infants. Neonatology 2008, 93, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Fehrholz, M.; Bersani, I.; Kramer, B.W.; Speer, C.P.; Kunzmann, S. Synergistic Effect of caffeine and glucocorticoids on expression of surfactant protein B (SP-B) mRNA. PLoS ONE 2012, 7, e51575. [Google Scholar] [CrossRef] [PubMed]
- Misra, A.L.; Pontani, R.B.; Vadlamani, N.L. Potentiation of morphine analgesia by caffeine. Br. J. Pharmacol. 1985, 84, 789–791. [Google Scholar] [CrossRef]
- Ryan, S.J.; Hassett, A.L.; Brummett, C.M.; Harris, R.E.; Clauw, D.J.; Harte, S.E. Caffeine as an opioid analgesic adjuvant in fibromyalgia. J. Pain Res. 2017, 10, 1801–1809. [Google Scholar] [CrossRef]
- Mercadante, S.; Serretta, R.; Casuccio, A. Effects of caffeine as an adjuvant to morphine in advanced cancer patients. A randomized, double-blind, placebo-controlled, crossover study. J. Pain Symptom Manag. 2001, 21, 369–372. [Google Scholar] [CrossRef] [PubMed]
- Suh, S.Y.; Choi, Y.S.; Oh, S.C.; Kim, Y.S.; Cho, K.; Bae, W.K.; Lee, J.H.; Seo, A.R.; Ahn, H.Y. Caffeine as an adjuvant therapy to opioids in cancer pain: A randomized, double-blind, placebo-controlled trial. J. Pain Symptom Manag. 2013, 46, 474–482. [Google Scholar] [CrossRef] [PubMed]
- Engelhardt, G.; Mauz, A.B.; Pairet, M. Role of caffeine in combined analgesic drugs from the point of view of experimental pharmacology. Arzneimittelforschung 1997, 47, 917–927. [Google Scholar] [PubMed]
- Gayawali, K.; Pandhi, P.; Sharma, P.L. Determination of the optimal analgesia-potentiating dose of caffeine and a study of its effect on the pharmacokinetics of aspirin in mice. Meth. Find. Exp. Clin. Pharmacol. 1991, 13, 529–533. [Google Scholar]
- Castañeda-Hernández, G.; Castillo-Méndez, M.S.; López-Muñoz, F.J.; Granados-Soto, V.; Flores-Murrieta, F.J. Potentiation by caffeine of the analgesic effect of aspirin in the pain-induced functional impairment model in the rat. Can. J. Physiol. Pharmacol. 1994, 72, 1127–1131. [Google Scholar] [CrossRef]
- Arnold, M.A.; Carr, D.B.; Togasaki, D.M.; Pian, M.C.; Martin, J.B. Caffeine stimulates beta-endorphin release in blood but not in cerebrospinal fluid. Life Sci. 1982, 31, 1017–1024. [Google Scholar] [CrossRef]
- Berkowitz, B.A.; Tarver, J.H.; Spector, S. Release of norepinephrine in the central nervous system by theophylline and caffeine. Eur. J. Pharmacol. 1970, 10, 64–71. [Google Scholar] [CrossRef]
- Corrodi, H.; Fuxe, K.; Jonsson, G. Effects of caffeine on central monoamine neurons. J. Pharm. Pharmacol. 1972, 24, 155–158. [Google Scholar] [CrossRef]
- Ortiz, M.I.; Castañeda-Hernández, G. Examination of the interaction between peripheral lumiracoxib and opioids on the 1% formalin test in rats. Eur. J. Pain 2008, 12, 233–240. [Google Scholar] [CrossRef]
- Sawynok, J.; Yaksh, T.L. Caffeine as an analgesic adjuvant: A review of pharmacology and mechanisms of action. Pharmacol. Rev. 1993, 45, 43–84. [Google Scholar] [PubMed]
- Díaz-Reval, M.I.; Carrillo-Munguía, N.; Martínez-Casas, M.; González-Trujano, M.E. Tramadol and caffeine produce synergistic interactions on antinociception measured in a formalin model. Pharmacol. Biochem. Behav. 2010, 97, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Bastia, E.; Schwarzschild, M. Therapeutic potential of adenosine A(2A) receptor antagonists in Parkinson’s disease. Pharmacol. Ther. 2005, 105, 267–310. [Google Scholar] [CrossRef]
- Simola, N.; Pinna, A.; Frau, L.; Morelli, M. Protective agents in Parkinson’s disease: Caffeine and adenosine A2A receptor antagonists. In Handbook of Neurotoxicity, 1st ed.; Kostrzewa, R.M., Ed.; Springer: New York, NY, USA, 2014; pp. 2281–2298. [Google Scholar]
- Bandookwala, M.; Sahu, A.K.; Thakkar, D.; Sharma, M.; Khairnar, A.; Sengupta, P. Edaravone-caffeine combination for the effective management of rotenone induced Parkinson’s disease in rats: An evidence based affirmative from a comparative analysis of behavior and biomarker expression. Neurosci. Lett. 2019, 711, 134438. [Google Scholar] [CrossRef]
- Ikram, M.; Park, T.J.; Ali, T.; Kim, M.O. Antioxidant and neuroprotective effects of caffeine against Alzheimer’s and Parkinson’s disease: Insight into the role of Nrf-2 and A2AR signaling. Antioxidants 2020, 9, 902. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.T.; Chan, L.; Bai, C.H. The effect of caffeine on the risk and progression of Parkinson’s Disease: A meta-analysis. Nutrients 2020, 12, 1860. [Google Scholar] [CrossRef] [PubMed]
- Aboutabl, M.E.; Salman, A.M.; Gamal el Din, A.A.; Yousreya, A.M. Simultaneous administration of coffee and rasagiline/L-dopa protects against paraquat-induced neurochemical and motor behavior impairments in vivo. Bull. Natl. Res. Cent. 2021, 45, 219. [Google Scholar] [CrossRef]
- Greenway, F.L. The safety and efficacy of pharmaceutical and herbal caffeine and ephedrine use as a weight loss agent. Obes. Rev. 2001, 2, 199–211. [Google Scholar] [CrossRef]
- Astrup, A.; Toubro, S.; Cannon, S.; Hein, P.; Madsen, J. Thermogenic synergism between ephedrine and caffeine in healthy volunteers: A double-blind, placebo-controlled study. Metabolism 1991, 40, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.G.; Arceneaux III, K.P.; Chu, J.T.; Jacob, G., Jr.; Schreiber, A.L.; Tipton, R.C.; Yu, Y.; Johnson, W.D.; Greenway, F.L.; Primeaux, S.D. The effect of caffeine and albuterol on body composition and metabolic rate. Obesity 2015, 23, 1830–1835. [Google Scholar] [CrossRef] [PubMed]
- Al-Majed, A.; Bakheit, A.H.; Abdel Aziz, H.A.; Alharbi, H.; Al-Jenoobi, F.I. Pioglitazone. In Profiles of Drug Substances, Excipients and Related Methodology, 1st ed.; Brittain, H.G., Ed.; Academic Press: Cambridge, MA, USA, 2016; Volume 41, pp. 379–438. [Google Scholar]
- Alshabi, A.M.; Alkahtani, S.A.; Shaikh, I.A.; Habeeb, M.S. Caffeine modulates pharmacokinetic and pharmacodynamic profiles of pioglitazone in diabetic rats: Impact on therapeutics. Saudi Med. J. 2021, 42, 151–160. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Drug | Structure | Activity | Caffeine Synergistic Effects |
|---|---|---|---|
| Ifosfamide (16) |  | DNA crosslinks by alkylation at guanine N-7 positions. | Interference with DNA damage repair processes and apoptosis induction. |
| Doxorubicin (17) |  | Intercalation and inhibition of DNA biosynthesis. | Inhibition of doxorubicin efflux from cancer cells and induction of apoptosis. |
| Cisplatin (18) |  | Binding to and interference with DNA transcription and replication. | Increase in CDDP toxicity and chromosome breakage. |
| Valproic acid (19) |  | Blockage of voltage-gated ion channels and inhibition of histone deacetylase. | Apoptosis induction at lower concentrations with respect to monotherapy. |
| Taxol (20) |  | Stabilizing and preventing microtubules’ depolymerization, causing cell death and cell-cycle arrest at the G2/M phase. | Inhibition of cell growth and arrest, leading to the death of pancreatic adenocarcinoma cells. |
| Metformin (21) |  | Alteration of the energy metabolism of the cell. | Inhibition of cell growth, an expansion of necrosis, and a reduction in tumor neo-vasculature. |
| 5-Fluorouracil (22) |  | DNA damage via fraudulent incorporation in nucleic acid synthesis. | Inhibition of cell proliferation and induction of ROS-dependent apoptosis. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ialongo, D.; Tudino, V.; Arpacioglu, M.; Messore, A.; Patacchini, E.; Costi, R.; Di Santo, R.; Madia, V.N. Synergistic Effects of Caffeine in Combination with Conventional Drugs: Perspectives of a Drug That Never Ages. Pharmaceuticals 2023, 16, 730. https://doi.org/10.3390/ph16050730
Ialongo D, Tudino V, Arpacioglu M, Messore A, Patacchini E, Costi R, Di Santo R, Madia VN. Synergistic Effects of Caffeine in Combination with Conventional Drugs: Perspectives of a Drug That Never Ages. Pharmaceuticals. 2023; 16(5):730. https://doi.org/10.3390/ph16050730
Chicago/Turabian StyleIalongo, Davide, Valeria Tudino, Merve Arpacioglu, Antonella Messore, Elisa Patacchini, Roberta Costi, Roberto Di Santo, and Valentina Noemi Madia. 2023. "Synergistic Effects of Caffeine in Combination with Conventional Drugs: Perspectives of a Drug That Never Ages" Pharmaceuticals 16, no. 5: 730. https://doi.org/10.3390/ph16050730
APA StyleIalongo, D., Tudino, V., Arpacioglu, M., Messore, A., Patacchini, E., Costi, R., Di Santo, R., & Madia, V. N. (2023). Synergistic Effects of Caffeine in Combination with Conventional Drugs: Perspectives of a Drug That Never Ages. Pharmaceuticals, 16(5), 730. https://doi.org/10.3390/ph16050730