
Canagliflozin Ameliorates Oxidative Stress and Autistic-like Features in Valproic-Acid-Induced Autism in Rats: Comparison with Aripiprazole Action


 and
and
Abstract
:1. Introduction
2. Results
2.1. Effects of Canagliflozin on Rats ASD-like Anxiety and Exploratory Behaviors
2.1.1. Open Field Test (OFT)
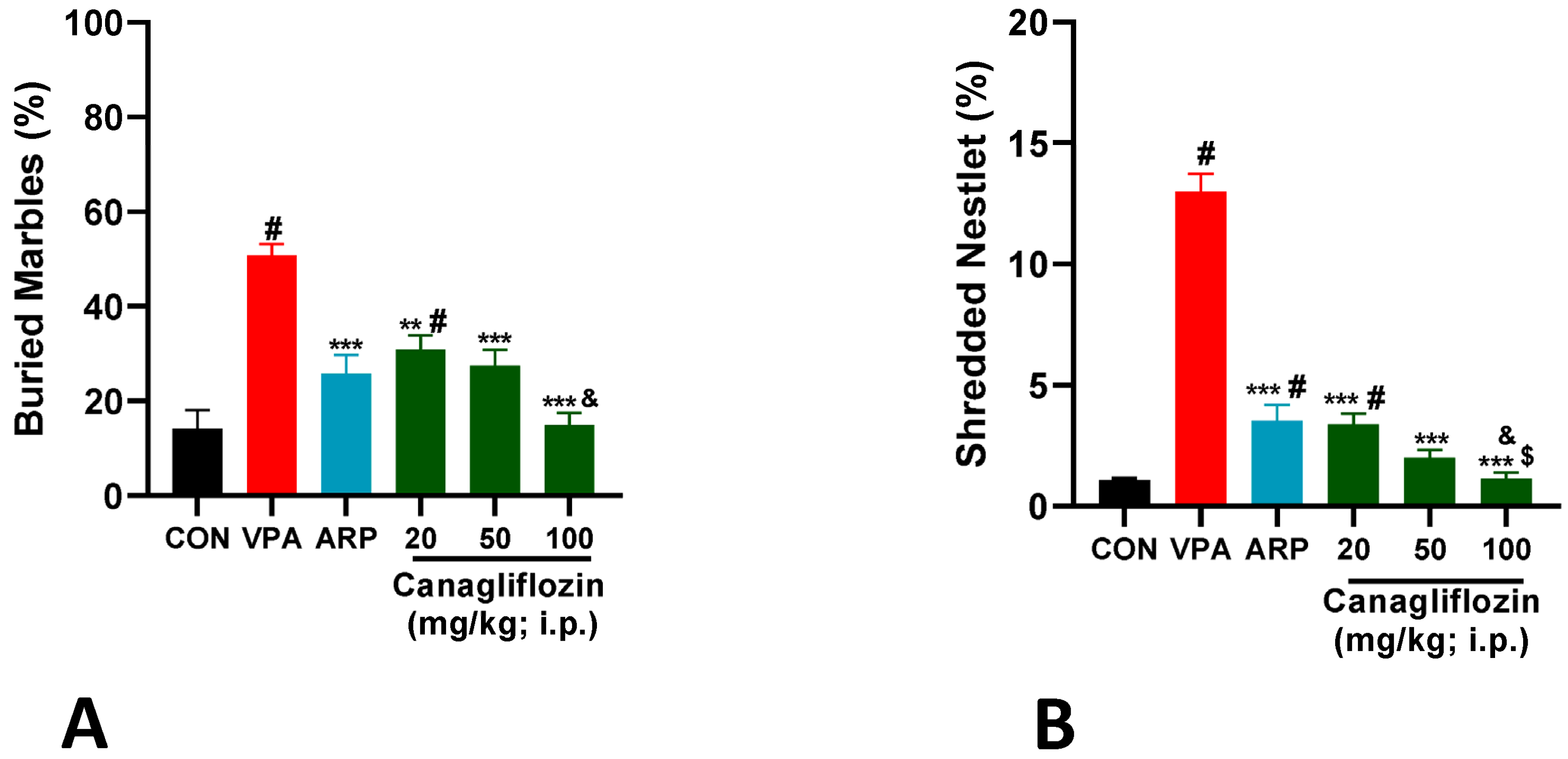
2.1.2. Marble-Burying Test (MBT)
2.1.3. Nestlet-Shredding Test (NST)
2.2. Effects of Canagliflozin on ASD Biomarkers
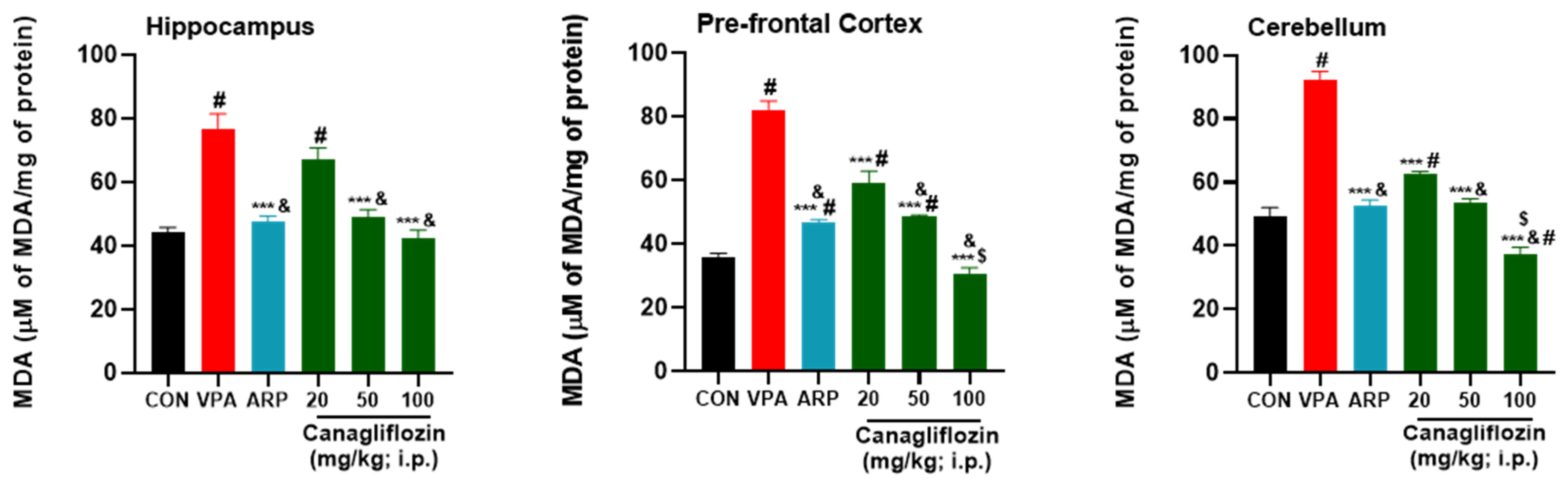
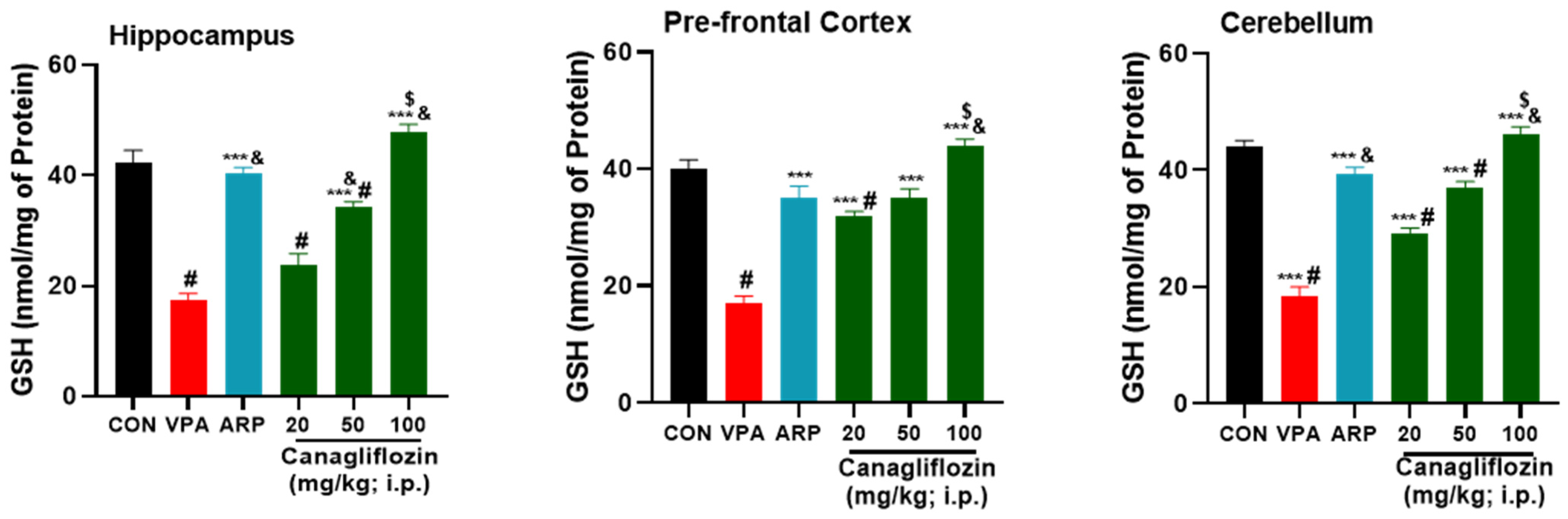
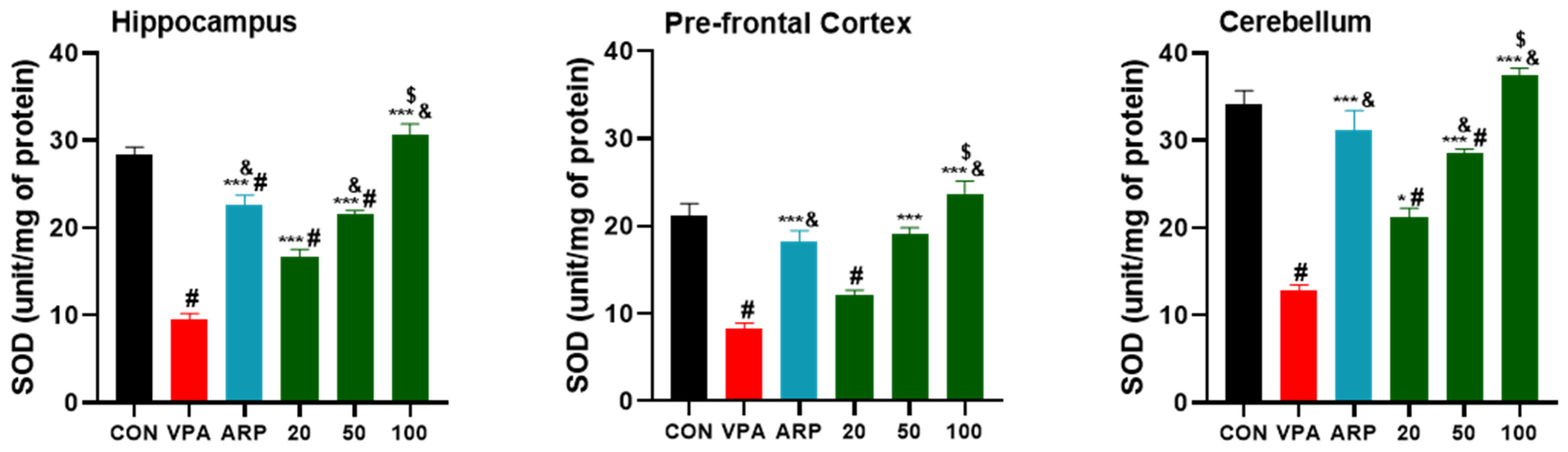
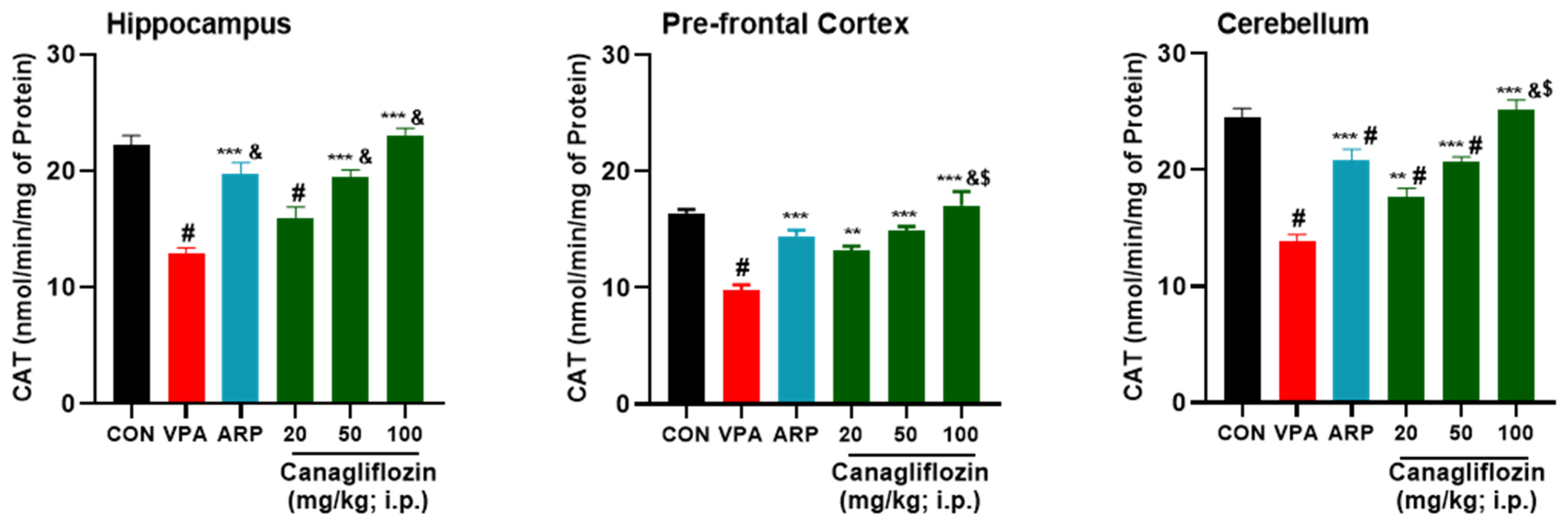
2.2.1. Oxidative Stress Biomarkers
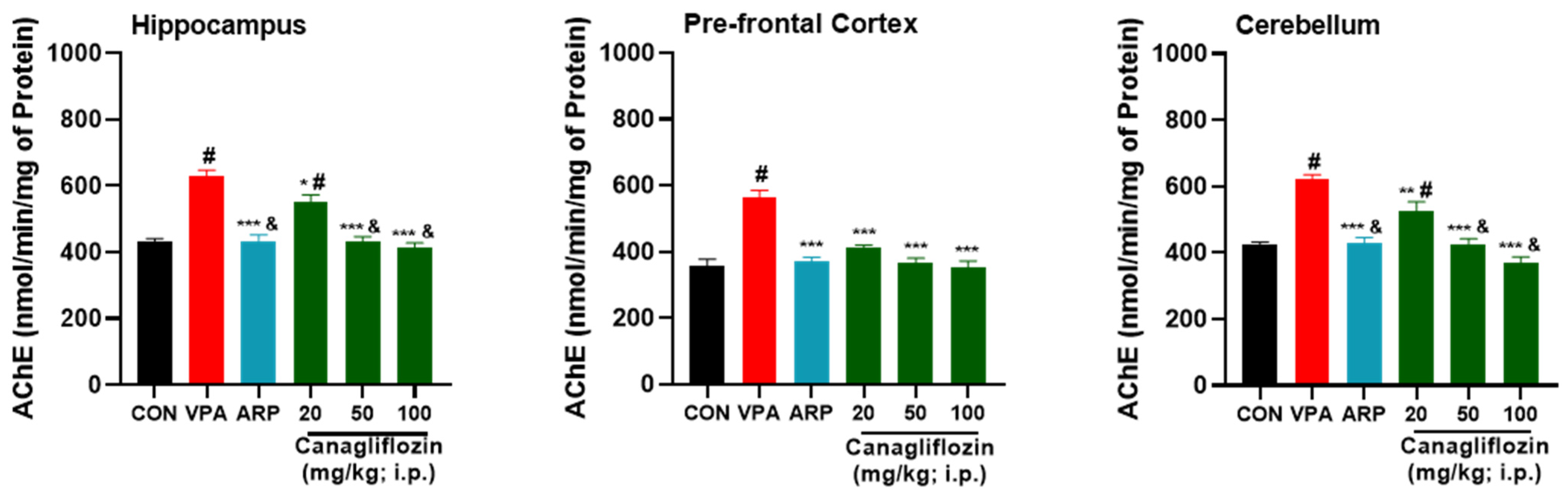
2.2.2. Acetylcholinesterase (AChE) Activity
2.3. Behavioral and ASD Biomarkers’ Response to Treatment Based on Gender
3. Discussion
4. Materials and Methods
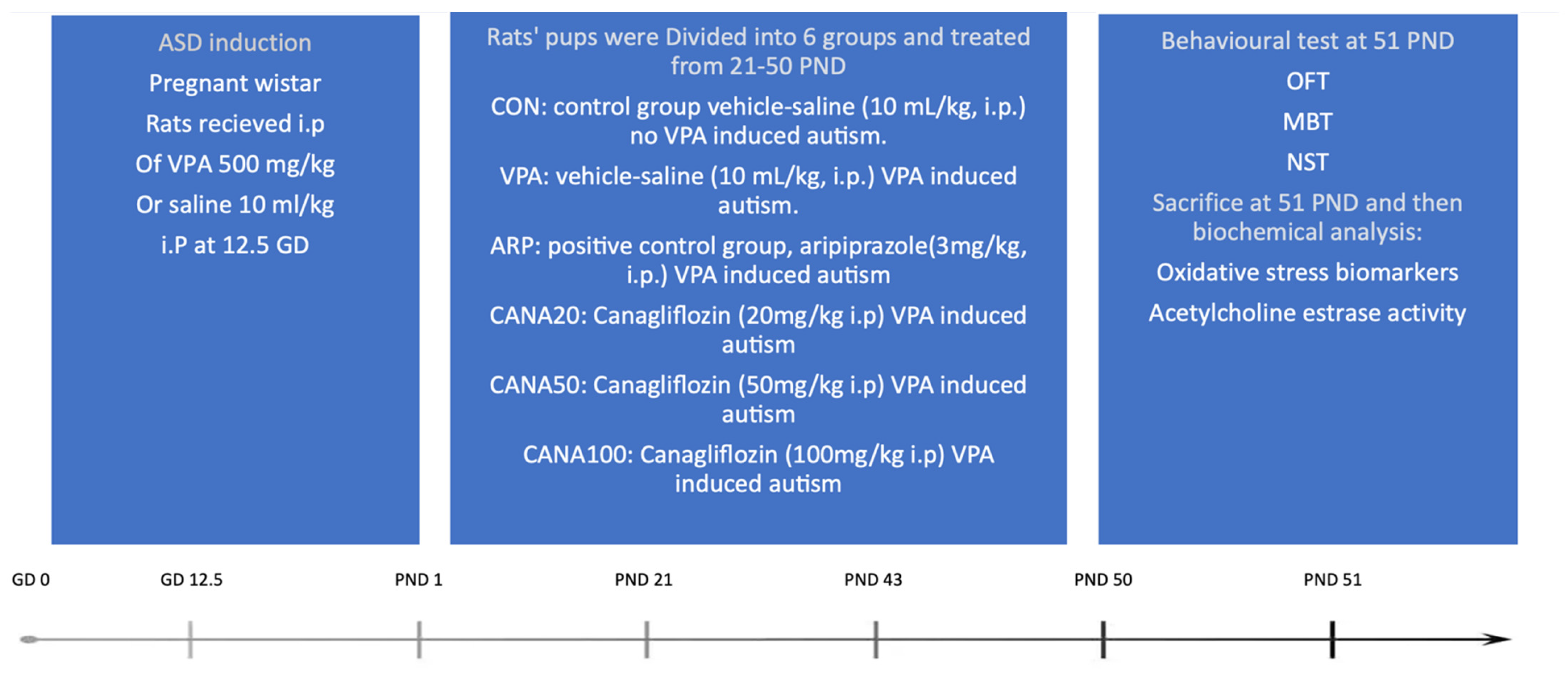
4.1. Animals
4.1.1. Research Design
4.1.2. Choice of Treatment and Dosages
4.1.3. Prenatal Treatment
4.1.4. Postnatal Treatment
4.2. Behavioral Tests
4.2.1. Open Field Test (OFT)
4.2.2. Marble-Burying Test (MBT)
4.2.3. Nestlet-Shredding Test (NST)
4.3. Biochemical Analyses
4.3.1. Brain Extraction and Conditioning for Biochemical Assessments
4.3.2. ASD Biomarkers Assessment Reagents and Experimental Kits
4.4. Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AChE | Acetylcholinesterase |
| ADA | American Diabetes Association |
| AD | Alzheimer’s disease |
| ARP | Aripiprazole |
| ASD | Autism spectrum disorder |
| CAT | Catalase |
| CNS | Central nervous system |
| ESC | European Society of Cardiology |
| FDA | Food and Drug Administration |
| GD | Gestational day |
| GSH | Glutathione |
| HF | Heart failure |
| i.p | Intraperitoneally |
| MDA | Malondialdehyde |
| PD | Parkinson’s disease |
| PND | Postnatal day |
| SGLT 1 | Sodium-glucose cotransporter 1 |
| SGLT 2 | Sodium-glucose cotransporter 2 |
| SOD | Superoxide dismutase |
| T2DM | Type 2 diabetes mellitus |
| VPA | Valproic acid |
References
- Jones, E.J.H.; Gliga, T.; Bedford, R.; Charman, T.; Johnson, M.H. Developmental pathways to autism: A review of prospective studies of infants at risk. Neurosci. Biobehav. Rev. 2014, 39, 1–33. [Google Scholar] [CrossRef]
- Nicholas, J.S.; Charles, J.M.; Carpenter, L.A.; King, L.B.; Jenner, W.; Spratt, E.G. Prevalence and characteristics of children with autism-spectrum disorders. Ann. Epidemiol. 2008, 18, 130–136. [Google Scholar] [CrossRef]
- Medavarapu, S.; Marella, L.L.; Sangem, A.; Kairam, R. Where is the evidence? A narrative literature review of the treatment modalities for autism spectrum disorders. Cureus 2019, 11, e3901. [Google Scholar] [CrossRef]
- Varamini, B.; Sikalidis, A.K.; Bradford, K.L. Resveratrol increases cerebral glycogen synthase kinase phosphorylation as well as protein levels of drebrin and transthyretin in mice: An exploratory study. Int. J. Food Sci. Nutr. 2014, 65, 89–96. [Google Scholar] [CrossRef]
- Indika, N.L.; Frye, R.E.; Rossignol, D.A.; Owens, S.C.; Senarathne, U.D.; Grabrucker, A.M.; Perera, R.; Engelen, M.P.; Deutz, N.E. The Rationale for Vitamin, Mineral, and Cofactor Treatment in the Precision Medical Care of Autism Spectrum Disorder. J. Pers. Med. 2023, 13, 252. [Google Scholar] [CrossRef]
- Elsabbagh, M.; Divan, G.; Koh, Y.J.; Kim, Y.S.; Kauchali, S.; Marcín, C.; Montiel-Nava, C.; Patel, V.; Paula, C.S.; Wang, C.; et al. Global prevalence of autism and other pervasive developmental disorders. Autism Res. 2012, 5, 160–179. [Google Scholar] [CrossRef]
- Nicolini, C.; Fahnestock, M. The valproic acid-induced rodent model of autism. Exp. Neurol. 2018, 299, 217–227. [Google Scholar] [CrossRef]
- Zhu, M.M.; Li, H.L.; Shi, L.H.; Chen, X.P.; Luo, J.; Zhang, Z.L. The pharmacogenomics of valproic acid. J. Hum. Genet. 2017, 62, 1009–1014. [Google Scholar] [CrossRef]
- Chaliha, D.; Albrecht, M.; Vaccarezza, M.; Takechi, R.; Lam, V.; Al-Salami, H.; Mamo, J. A systematic review of the valproic-acid-induced rodent model of autism. Dev. Neurosci. 2020, 42, 12–48. [Google Scholar] [CrossRef]
- Bell, R.M.; Yellon, D.M. SGLT2 inhibitors: Hypotheses on the mechanism of cardiovascular protection. Lancet Diabetes Endocrinol. 2018, 6, 435–437. [Google Scholar] [CrossRef]
- Cowie, M.R.; Fisher, M. SGLT2 inhibitors: Mechanisms of cardiovascular benefit beyond glycaemic control. Nat. Rev. Cardiol. 2020, 17, 761–772. [Google Scholar] [CrossRef]
- Davies, M.J.; Aroda, V.R.; Collins, B.S.; Gabbay, R.A.; Green, J.; Maruthur, N.M.; Rosas, S.E.; Del Prato, S.; Mathieu, C.; Mingrone, G.; et al. Management of hyperglycemia in type 2 diabetes, 2022. A consensus report by the American Diabetes Association (ADA) and the European Association for the Study of Diabetes (EASD). Diabetes Care 2022, 45, 2753–2786. [Google Scholar] [CrossRef]
- Kolesnik, E.; Scherr, D.; Rohrer, U.; Benedikt, M.; Manninger, M.; Sourij, H.; von Lewinski, D. SGLT2 inhibitors and their antiarrhythmic properties. Int. J. Mol. Sci. 2022, 23, 1678. [Google Scholar] [CrossRef]
- Alicic, R.Z.; Johnson, E.J.; Tuttle, K.R. SGLT2 inhibition for the prevention and treatment of diabetic kidney disease: A review. Am. J. Kidney Dis. 2018, 72, 267–277. [Google Scholar] [CrossRef]
- Nakhal, M.M.; Aburuz, S.; Sadek, B.; Akour, A. Repurposing SGLT2 Inhibitors for Neurological Disorders: A Focus on the Autism Spectrum Disorder. Molecules 2022, 27, 7174. [Google Scholar] [CrossRef]
- Shakil, S. Molecular interaction of anti-diabetic drugs with acetylcholinesterase and sodium glucose co-transporter 2. J. Cell. Biochem. 2017, 118, 3855–3865. [Google Scholar] [CrossRef]
- Shaikh, S.; Danish Rizvi, S.M.; Suhail, T.; Shakil, S.; MAbuzenadah, A.; Anis, R.; Naaz, D.; Dallol, A.; Haneef, M.; Ahmad, A.; et al. Prediction of anti-diabetic drugs as dual inhibitors against acetylcholinesterase and beta-secretase: A neuroinformatics study. CNS Neurol. Disord.-Drug Targets (Former. Curr. Drug Targets-CNS Neurol. Disord.) 2016, 15, 1216–1221. [Google Scholar]
- Xue, L.; Yuan, X.; Zhang, S.; Zhao, X. Investigating the effects of dapagliflozin on cardiac function, inflammatory response, and cardiovascular outcome in patients with STEMI complicated with T2DM after PCI. Evid.-Based Complement. Altern. Med. 2021, 2021, 9388562. [Google Scholar] [CrossRef]
- Ganbaatar, B.; Fukuda, D.; Shinohara, M.; Yagi, S.; Kusunose, K.; Yamada, H.; Soeki, T.; Hirata, K.I.; Sata, M. Empagliflozin ameliorates endothelial dysfunction and suppresses atherogenesis in diabetic apolipoprotein E-deficient mice. Eur. J. Pharmacol. 2020, 875, 173040. [Google Scholar] [CrossRef]
- Liu, H.; Sridhar, V.S.; Lovblom, L.E.; Lytvyn, Y.; Burger, D.; Burns, K.; Brinc, D.; Lawler, P.R.; Cherney, D.Z. Markers of kidney injury, inflammation, and fibrosis associated with ertugliflozin in patients with CKD and diabetes. Kidney Int. Rep. 2021, 6, 2095–2104. [Google Scholar] [CrossRef]
- Cefalo, C.M.; Cinti, F.; Moffa, S.; Impronta, F.; Sorice, G.P.; Mezza, T.; Pontecorvi, A.; Giaccari, A. Sotagliflozin, the first dual SGLT inhibitor: Current outlook and perspectives. Cardiovasc. Diabetol. 2019, 18, 1–4. [Google Scholar] [CrossRef] [PubMed]
- Hasan, R.; Lasker, S.; Hasan, A.; Zerin, F.; Zamila, M.; Parvez, F.; Rahman, M.M.; Khan, F.; Subhan, N.; Alam, M.A. Canagliflozin ameliorates renal oxidative stress and inflammation by stimulating AMPK–Akt–eNOS pathway in the isoprenaline-induced oxidative stress model. Sci. Rep. 2020, 10, 14659. [Google Scholar] [CrossRef]
- Zaibi, N.; Li, P.; Xu, S.Z. Protective effects of dapagliflozin against oxidative stress-induced cell injury in human proximal tubular cells. PLoS ONE 2021, 16, e0247234. [Google Scholar] [CrossRef] [PubMed]
- Iannantuoni, F.M.; de Marañon, A.; Diaz-Morales, N.; Falcon, R.; Bañuls, C.; Abad-Jimenez, Z.; Victor, V.M.; Hernandez-Mijares, A.; Rovira-Llopis, S. The SGLT2 inhibitor empagliflozin ameliorates the inflammatory profile in type 2 diabetic patients and promotes an antioxidant response in leukocytes. J. Clin. Med. 2019, 8, 1814. [Google Scholar] [CrossRef] [PubMed]
- Croteau, D.; Luptak, I.; Chambers, J.M.; Hobai, I.; Panagia, M.; Pimentel, D.R.; Siwik, D.A.; Qin, F.; Colucci, W.S. Effects of sodium-glucose linked transporter 2 inhibition with ertugliflozin on mitochondrial function, energetics, and metabolic gene expression in the presence and absence of diabetes mellitus in mice. J. Am. Heart Assoc. 2021, 10, e019995. [Google Scholar] [CrossRef] [PubMed]
- Scholtes, R.A.; Mosterd, C.M.; Hesp, A.C.; Smits, M.M.; Heerspink, H.J.; van Raalte, D.H. Mechanisms underlying the blood pressure-lowering effects of empagliflozin, losartan and their combination in people with type 2 diabetes: A secondary analysis of a randomized crossover trial. Diabetes Obes. Metab. 2023, 25, 198–207. [Google Scholar] [CrossRef]
- Zhou, J.; Zhu, J.; Yu, S.J.; Ma, H.L.; Chen, J.; Ding, X.F.; Chen, G.; Liang, Y.; Zhang, Q. Sodium-glucose co-transporter-2 (SGLT-2) inhibition reduces glucose uptake to induce breast cancer cell growth arrest through AMPK/mTOR pathway. Biomed. Pharmacother. 2020, 132, 110821. [Google Scholar] [CrossRef]
- Sun, X.; Han, F.; Lu, Q.; Li, X.; Ren, D.; Zhang, J.; Han, Y.; Xiang, Y.K.; Li, J. Empagliflozin ameliorates obesity-related cardiac dysfunction by regulating Sestrin2-mediated AMPK-mTOR signaling and redox homeostasis in high-fat diet–induced obese mice. Diabetes 2020, 69, 1292–1305. [Google Scholar] [CrossRef]
- Moellmann, J.; Mann, P.; Krueger, K.; Klinkhammer, B.; Boor, P.; Marx, N.; Lehrke, M. The SGLT2 inhibitor ertugliflozin causes a switch of cardiac substrate utilization leading to reduced cardiac mTOR-signaling, unfolded protein response and apoptosis. Eur. Heart J. 2021, 42, ehab724.3289. [Google Scholar] [CrossRef]
- Eissa, N.; Azimullah, S.; Jayaprakash, P.; Jayaraj, R.L.; Reiner, D.; Ojha, S.K.; Beiram, R.; Stark, H.; Łażewska, D.; Kieć-Kononowicz, K.; et al. The dual-active histamine H3 receptor antagonist and acetylcholine esterase inhibitor E100 alleviates autistic-like behaviors and oxidative stress in valproic acid induced autism in mice. Int. J. Mol. Sci. 2020, 21, 3996. [Google Scholar] [CrossRef]
- Plyusnina, I.; Oskina, I. Behavioral and adrenocortical responses to open-field test in rats selected for reduced aggressiveness toward humans. Physiol. Behav. 1997, 61, 381–385. [Google Scholar] [CrossRef] [PubMed]
- Al Sagheer, T.; Haida, O.; Balbous, A.; Francheteau, M.; Matas, E.; Fernagut, P.O.; Jaber, M. Motor impairments correlate with social deficits and restricted neuronal loss in an environmental model of autism. Int. J. Neuropsychopharmacol. 2018, 21, 871–882. [Google Scholar] [CrossRef] [PubMed]
- Angoa-Pérez, M.; Kane, M.J.; Briggs, D.I.; Francescutti, D.M.; Kuhn, D.M. Marble burying and nestlet shredding as tests of repetitive, compulsive-like behaviors in mice. J. Vis. Exp. JoVE 2013. [Google Scholar] [CrossRef] [PubMed]
- Bjørklund, G.; Meguid, N.A.; El-Bana, M.A.; Tinkov, A.A.; Saad, K.; Dadar, M.; Hemimi, M.; Skalny, A.V.; Hosnedlová, B.; Kizek, R.; et al. Oxidative stress in autism spectrum disorder. Mol. Neurobiol. 2020, 57, 2314–2332. [Google Scholar] [CrossRef] [PubMed]
- Jayaprakash, P.; Isaev, D.; Shabbir, W.; Lorke, D.E.; Sadek, B.; Oz, M. Curcumin potentiates α7 nicotinic acetylcholine receptors and alleviates autistic-like social deficits and brain oxidative stress status in mice. Int. J. Mol. Sci. 2021, 22, 7251. [Google Scholar] [CrossRef]
- Eissa, N.; Venkatachalam, K.; Jayaprakash, P.; Falkenstein, M.; Dubiel, M.; Frank, A.; Reiner-Link, D.; Stark, H.; Sadek, B. The multi-targeting ligand ST-2223 with histamine H3 receptor and dopamine D2/D3 receptor antagonist properties mitigates autism-like repetitive behaviors and brain oxidative stress in mice. Int. J. Mol. Sci. 2021, 22, 1947. [Google Scholar] [CrossRef]
- Natesan, S.; Reckless, G.E.; Nobrega, J.N.; Fletcher, P.J.; Kapur, S. Dissociation between in vivo occupancy and functional antagonism of dopamine D2 receptors: Comparing aripiprazole to other antipsychotics in animal models. Neuropsychopharmacology 2006, 31, 1854–1863. [Google Scholar] [CrossRef]
- Silverman, J.L.; Tolu, S.S.; Barkan, C.L.; Crawley, J.N. Repetitive self-grooming behavior in the BTBR mouse model of autism is blocked by the mGluR5 antagonist MPEP. Neuropsychopharmacology 2010, 35, 976–989. [Google Scholar] [CrossRef]
- Sanavi, M.A.; Ghazvini, H.; Zargari, M.; Ghalehnoei, H.; Hosseini-Khah, Z. Effects of clozapine and risperidone antipsychotic drugs on the expression of CACNA1C and behavioral changes in rat ‘Ketamine model of schizophrenia. Neurosci. Lett. 2022, 770, 136354. [Google Scholar] [CrossRef]
- Deng, W.; Ke, H.; Wang, S.; Li, Z.; Li, S.; Lv, P.; Li, F.; Chen, Y. Metformin alleviates autistic-like behaviors elicited by high-fat diet consumption and modulates the crosstalk between serotonin and gut microbiota in mice. Behav. Neurol. 2022, 2022, 6711160. [Google Scholar] [CrossRef]
- Estaphan, S.; Curpăn, A.S.; Khalifa, D.; Rashed, L.; Ciobica, A.; Cantemir, A.; Ciobica, A.; Trus, C.; Ali, M.; ShamsEldeen, A. Combined Low Dose of Ketamine and Social Isolation: A Possible Model of Induced Chronic Schizophrenia-Like Symptoms in Male Albino Rats. Brain Sci. 2021, 11, 917. [Google Scholar] [CrossRef] [PubMed]
- Amin, E.F.; Rifaai, R.A.; Abdel-latif, R.G. Empagliflozin attenuates transient cerebral ischemia/reperfusion injury in hyperglycemic rats via repressing oxidative–inflammatory–apoptotic pathway. Fundam. Clin. Pharmacol. 2020, 34, 548–558. [Google Scholar] [CrossRef] [PubMed]
- Eren, İ.; Nazıroğlu, M.; Demirdaş, A. Protective effects of lamotrigine, aripiprazole and escitalopram on depression-induced oxidative stress in rat brain. Neurochem. Res. 2007, 32, 1188–1195. [Google Scholar] [CrossRef]
- Hassanein, E.H.; Saleh, F.M.; Ali, F.E.; Rashwan, E.K.; Atwa, A.M.; Abd El-Ghafar, O.A. Neuroprotective effect of canagliflozin against cisplatin-induced cerebral cortex injury is mediated by regulation of HO-1/PPAR-γ, SIRT1/FOXO-3, JNK/AP-1, TLR4/iNOS, and Ang II/Ang 1–7 signals. Immunopharmacol. Immunotoxicol. 2022, 10, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Sandhya, T.; Sowjanya, J.; Veeresh, B. Bacopa monniera (L.) Wettst ameliorates behavioral alterations and oxidative markers in sodium valproate induced autism in rats. Neurochem. Res. 2012, 37, 1121–1131. [Google Scholar] [CrossRef]
- Arafa, N.M.; Ali, E.H.; Hassan, M.K. Canagliflozin prevents scopolamine-induced memory impairment in rats: Comparison with galantamine hydrobromide action. Chem. -Biol. Interact. 2017, 277, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Elgamal, M.A.; Khodeer, D.; Abdel-Wahab, B.A.; Aziz, I.A.; Alzahrani, A.R.; Mostafa, Y.M.; Ali, A.A.; El-sayed, N. Canagliflozin Alleviates Valproic Acid-induced Rat Autism: The Role of PTEN/PDK/PPAR-γ signaling pathways. Front. Pharmacol. 2023, 14, 388. [Google Scholar] [CrossRef]
- Mony, T.J.; Lee, J.W.; Kim, S.S.; Chun, W.; Lee, H.J. Early postnatal valproic acid exposure increase the protein level of astrocyte markers in frontal cortex of rat. Clin. Psychopharmacol. Neurosci. 2018, 16, 214. [Google Scholar] [CrossRef]
- Mony, T.J.; Lee, J.W.; Dreyfus, C.; DiCicco-Bloom, E.; Lee, H.J. Valproic acid exposure during early postnatal gliogenesis leads to autistic-like behaviors in rats. Clin. Psychopharmacol. Neurosci. 2016, 14, 338. [Google Scholar] [CrossRef]
- Iwata, K.; Matsuzaki, H.; Takei, N.; Manabe, T.; Mori, N. Animal models of autism: An epigenetic and environmental viewpoint. J. Cent. Nerv. Syst. Dis. 2010, 2, S6188. [Google Scholar] [CrossRef]
- Cariveau, T.; McCracken, C.E.; Bradshaw, J.; Postorino, V.; Shillingsburg, M.A.; McDougle, C.J.; Aman, M.G.; McCracken, J.T.; Tierney, E.; Johnson, C.; et al. Gender Differences in Treatment-Seeking Youth with Autism Spectrum Disorder. J. Child Fam. Stud. 2021, 30, 784–792. [Google Scholar] [CrossRef]
- Charan, J.; Kantharia, N. How to calculate sample size in animal studies? J. Pharmacol. Pharmacother. 2013, 4, 303–306. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Wahab, A.; Mahmoud, W.; Al-Harizy, R. Comparative renal protective effects of canagliflozin and telmisartan in a rat model of diabetic nephropathy. J. Nephrol. Renal. Ther. 2016, 2, 10. [Google Scholar] [CrossRef] [PubMed]
- Solares-Pascasio, J.I.; Ceballos, G.; Calzada, F.; Barbosa, E.; Velazquez, C. Antihyperglycemic and lipid profile effects of Salvia amarissima Ortega on streptozocin-induced type 2 diabetic mice. Molecules 2021, 26, 947. [Google Scholar] [CrossRef]
- Kumar, A.; Negi, A.S.; Chauhan, A.; Semwal, R.; Kumar, R.; Semwal, R.B.; Singh, R.; Joshi, T.; Chandra, S.; Joshi, S.K.; et al. Formulation and evaluation of SGLT2 inhibitory effect of a polyherbal mixture inspired from Ayurvedic system of medicine. J. Tradit. Complement. Med. 2022, 12, 477–487. [Google Scholar] [CrossRef]
- Bäckström, P.; Etelälahti, T.J.; Hyytiä, P. Attenuation of reinforcing and psychomotor stimulant effects of amphetamine by aripiprazole. Addict. Biol. 2011, 16, 55–63. [Google Scholar] [CrossRef]
- Kehr, J.; Yoshitake, T.; Ichinose, F.; Yoshitake, S.; Kiss, B.; Gyertyán, I.; Adham, N. Effects of cariprazine on extracellular levels of glutamate, GABA, dopamine, noradrenaline and serotonin in the medial prefrontal cortex in the rat phencyclidine model of schizophrenia studied by microdialysis and simultaneous recordings of locomotor activity. Psychopharmacology 2018, 235, 1593–1607. [Google Scholar]
- Saft, P.; Toledo-Cardenas, R.; Coria-Avila, G.A.; Perez-Pouchoulen, M.; Brug, B.; Hernandez, M.E.; Manzo, J. Characterization of four types of tail abnormalities in rats treated prenatally with valproic acid. Eneurobiología 2014, 5, 9. [Google Scholar]
- Ghasemi, A.; Jeddi, S.; Kashfi, K. The laboratory rat: Age and body weight matter. EXCLI J. 2021, 20, 1431. [Google Scholar]
- Fuller, E.A.; Kaiser, A.P. The effects of early intervention on social communication outcomes for children with autism spectrum disorder: A meta-analysis. J. Autism Dev. Disord. 2020, 50, 1683–1700. [Google Scholar] [CrossRef]
- Paudel, R.; Raj, K.; Gupta, Y.K.; Singh, S. Oxiracetam and zinc ameliorates autism-like symptoms in propionic acid model of rats. Neurotox. Res. 2020, 37, 815–826. [Google Scholar] [CrossRef]
- Kim, J.W.; Park, K.; Kang, R.J.; Gonzales, E.L.; Kim, D.G.; Oh, H.A.; Seung, H.; Ko, M.J.; Kwon, K.J.; Kim, K.C.; et al. Pharmacological modulation of AMPA receptor rescues social impairments in animal models of autism. Neuropsychopharmacology 2019, 44, 314–323. [Google Scholar] [CrossRef]
- Kavitha, U.; Kayalvizhi, E.; Revathy, K.; Brindha, T.R.; Muthulakshmi, R.; Chandrasekar, M. A systematic review on Valproate induced rat model of autism: Pathophysiology, treatment, developmental and neurobehavioral assessment of rat offsprings. Biomedicine 2021, 41, 9–15. [Google Scholar] [CrossRef]
- Arakawa, H. Implication of the social function of excessive self-grooming behavior in BTBR T+ ltpr3tf/J mice as an idiopathic model of autism. Physiol. Behav. 2021, 237, 113432. [Google Scholar] [CrossRef]
- Lazic, S.E. Analytical strategies for the marble burying test: Avoiding impossible predictions and invalid p-values. BMC Res. Notes 2015, 8, 141. [Google Scholar] [CrossRef]
- Eissa, N.; Azimullah, S.; Jayaprakash, P.; Jayaraj, R.L.; Reiner, D.; Ojha, S.K.; Beiram, R.; Stark, H.; Łażewska, D.; Kieć-Kononowicz, K.; et al. The dual-active histamine H3 receptor antagonist and acetylcholine esterase inhibitor E100 ameliorates stereotyped repetitive behavior and neuroinflammmation in sodium valproate induced autism in mice. Chem.-Biol. Interact. 2019, 312, 108775. [Google Scholar] [CrossRef]
- Gage, G.J.; Kipke, D.R.; Shain, W. Whole animal perfusion fixation for rodents. JoVE (J. Vis. Exp.) 2012, 30, e3564. [Google Scholar]
- Bastaki, S.M.; Abdulrazzaq, Y.M.; Shafiullah, M.; Więcek, M.; Kieć-Kononowicz, K.; Sadek, B. Anticonvulsant and reproductive toxicological studies of the imidazole-based histamine H3R antagonist 2-18 in mice. Drug Des. Dev. Ther. 2018, 12, 179–194. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Control | VPA Group | ARP | Canagliflozin (20 mg) (Group 4) | Canagliflozin (50 mg) (Group 5) | Canagliflozin (100 mg) (Group 6) |
|---|---|---|---|---|---|---|
| Time spent at center (s) | 14.8 ± 3.18 | 3.00 ± 0.89 # | 9.33 ± 4.41 #,* | 7.16 ± 2.31 # | 7.83 ± 1.72 #,* | 13.00 ± 3.09 *,& |
| Time spent at periphery (min) | 9.45 ± 0.31 | 9.57 ± 0.01 # | 9.50 ± 0.04 * | 9.52 ± 0.04 # | 9.52 ± 0.02 # | 9.47 ± 0.03 * |
| Locomotion (s) | 147 ± 42.7 | 303 ± 140 # | 157 ± 41.6 * | 161 ± 34.9 * | 154 ± 44.7 * | 147 ± 33.6 * |
| Distance traveled (m) | 15.8 ± 5.37 | 27.9 ± 10.3 # | 17.3 ± 3.68 * | 21.58 ± 6.06 | 20.0 ± 5.14 | 15.88 ± 1.59 * |
| Grooming (n) | 5.33 ± 1.03 | 9.50 ± 2.88 # | 8.00 ± 3.03 | 9.83 ± 1.72 # | 7.17 ± 1.72 | 5.50 ± 0.83 *,& |
| Defecation (n) | 3.33 ± 0.81 | 6.00 ± 1.78 # | 4.00 ± 1.41 | 5.00 ± 1.09 | 4.17 ± 1.72 | 3.83 ± 0.75 |
| Parameter/Group (Mean ± SD) | Control | p-Value | VPA | p-Value | ARP | p-Value | CANA20 | p-Value | CANA50 | p-Value | CANA100 | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M | F | M | F | M | F | M | F | M | F | M | F | |||||||
| Time Spent at Center (s) | 16.0 ± 4.3 | 13.6 ± 1.5 | 0.3 | 2.3 ± 0.5 | 3.6 ± 0.5 | 0.5 | 10.6 ± 2.5 | 8 ± 6.1 | 0.2 | 6.00 ± 1.00 | 8.3 ± 2.8 | 0.3 | 9 ± 1.7 | 6.6 ± 0.5 | 0.3 | 12.6 ± 3.7 | 13.0 ± 3 | 0.7 |
| Time Spent at Periphery (min) | 9.4 ± 0.04 | 9.4 ± 0.01 | 0.3 | 9.5 ± 0.01 | 9.5 ± 0.01 | 0.6 | 9.4 ± 0.02 | 9.5 ± 0.06 | 0.2 | 9 ± 0.01 | 9.5 ± 0.04 | 0.1 | 9.5 ± 0.01 | 9.5 ± 0.01 | 0.3 | 9.4 ± 0.03 | 9.4 ± 0.03 | 0.7 |
| Locomotion (s) | 160 ± 59.9 | 133.3± 21 | 0.6 | 337 ± 173 | 268 ± 123 | 0.2 | 158.3 ± 61.6 | 156 ± 23 | 0.9 | 137 ± 28.5 | 184.3 ± 22 | 0.4 | 143 ± 53.3 | 165 ± 42.2 | 0.7 | 151.6 ± 50.5 | 141.6 ± 13.5 | 0.8 |
| Distance Traveled (m) | 17.7 ± 7.5 | 13.9 ± 2.1 | 0.4 | 32.1 ± 13.0 | 23.6 ± 6.5 | 0.1 | 16.9 ± 5 | 17.7 ± 2.8 | 0.8 | 26.3 ± 3.50 | 16.8 ± 3.4 | 0.05 | 21.1 ± 6.7 | 18.8 ± 4.1 | 0.6 | 16.1 ± 1.6 | 15.7 ± 1.8 | 0.9 |
| Grooming (mean ± SD) | 4.6 ± 0.5 | 6 ± 1 | 0.4 | 9.00 ± 4.30 | 10 ± 1 | 0.5 | 8.6 ± 4 | 7.3 ± 2.3 | 0.4 | 10.3 ± 2.08 | 9.3 ± 1.5 | 0.5 | 7.3 ± 0.6 | 7 ± 2.6 | 0.8 | 5.3 ± 1.1 | 5.6 ± 0.5 | 0.8 |
| Marble Buried (%) | 18.3 ± 12.6 | 10 ± 5 | 0.2 | 55 ± 5 | 46.6 ± 2.8 | 0.2 | 25 ± 5 | 26.6 ± 14.4 | 0.7 | 31.6 ± 10.4 | 30 ± 5 | 0.7 | 21.6 ± 7.6 | 33.3 ± 2.8 | 0.08 | 15 ± 5 | 15 ± 8.6 | 1 |
| Nestlet Shredded (%) | 1.1± 0.2 | 1.1± 0.1 | 0.9 | 14.1 * ± 1.9 | 11.9 ± 1.03 | 0.011 * | 4.83 ** ± 0.80 | 2.2 ± 1.02 | 0.002 ** | 3.7 ± 1.30 | 3 ± 0.7 | 0.3 | 2.5 ± 0.8 | 1.4 ± 0.2 | 0.1 | 1.5 ± 0.5 | 0.7 ± 0.5 | 0.2 |
| MDA (μm MDA/mg of protein) | ||||||||||||||||||
| Hippocampus | 46.6 ± 3.2 | 42.1 ± 1.6 | 0.4 | 81.0 ± 14.7 | 72.7 ± 6.7 | 0.1 | 50.9 ± 1.9 | 44 ± 3.7 | 0.2 | 67.6 ± 1.30 | 66.6 ± 14.1 | 0.86 | 53.4 ± 4.1 | 44.8 ± 1.4 | 0.1 | 45.9 ± 6.2 | 38.9 ± 4.8 | 0.2 |
| Prefrontal Cortex | 34.7 ± 1.9 | 36.8 ± 4.2 | 0.5 | 84.6 ± 4.10 | 79.8 ± 8.6 | 0.2 | 46 ± 2.8 | 47.4 ± 1.5 | 0.7 | 53.40 * ± 10.40 | 64.9 ± 1.10 | 0.006 ** | 47.7 ± 0.5 | 49.3 ± 1.8 | 0.6 | 33.3 ± 2.6 | 28.2 ± 4.6 | 0.1 |
| Cerebellum | 54.3 ± 5.30 | 44.5 ± 2.50 | 0.011 * | 92.6 ± 7.70 | 92.3 ± 5.20 | 0.9 | 53.3 ± 4.6 | 51.4 ± 6 | 0.6 | 63.30 ± 2.4 | 61.6 ± 1.80 | 0.65 | 52.6 ± 3 | 54.1 ± 4.4 | 0.6 | 42.5 ** ± 2.30 | 31.7 ± 1.1 | 0.006 ** |
| GSH (nmol/mg of protein) | ||||||||||||||||||
| Hippocampus | 41.7 ± 2.8 | 43.2 ± 7.5 | 0.6 | 18 ± 4.3 | 16.8 ± 1.3 | 0.7 | 40.5 ± 3.4 | 40.6 ± 0.6 | 0.9 | 25.4 ± 6.9 | 22.2 ± 2.6 | 0.3 | 33.7 ± 1.2 | 34.9 ± 3.4 | 0.7 | 45 ± 1 | 50.7 ± 2.2 | 0.07 |
| Prefrontal Cortex | 38.8 ± 3.9 | 41.2 ± 4.0 | 0.4 | 16.8 ± 2.1 | 17.2 ± 4.2 | 0.8 | 31.4 ± 3.6 | 38.6 ± 3.4 | 0.019 * | 31.6 ± 1.2 | 32.2 ± 2.8 | 0.8 | 34.3 ± 4.5 | 35.7 ± 4.1 | 0.6 | 44.3 ± 3.1 | 43.6 ± 3.2 | 0.8 |
| Cerebellum | 43.6 ± 1.8 | 44.6 ± 3.1 | 0.6 | 17.4 ± 1.3 | 19.3 ± 5.8 | 0.4 | 40.7 ± 2.1 | 38.1 ± 2.8 | 0.3 | 28 ± 2.1 | 30.4 ± 1.5 | 0.3 | 37.8 ± 3.7 | 35.9 ± 1.8 | 0.4 | 46.3 ± 3.2 | 45.8 ± 4 | 0.8 |
| GSH (nmol/mg of protein) | ||||||||||||||||||
| Hippocampus | 41.7 ± 2.8 | 43.2 ± 7.5 | 0.6 | 18 ± 4.3 | 16.8 ± 1.3 | 0.7 | 40.5 ± 3.4 | 40.6 ± 0.6 | 0.9 | 25.4 ± 6.9 | 22.2 ± 2.6 | 0.3 | 33.7 ± 1.2 | 34.9 ± 3.4 | 0.7 | 45 ± 1 | 50.7 ± 2.2 | 0.07 |
| Prefrontal Cortex | 38.8 ± 3.9 | 41.2 ± 4 | 0.4 | 16.8 ± 2.1 | 17.2 ± 4.2 | 0.8 | 31.4 ± 3.6 | 38.6 ± 3.4 | 0.01 * | 31.6 ± 1.2 | 32.2 ± 2.8 | 0.8 | 34.3 ± 4.5 | 35.7 ± 4.1 | 0.6 | 44 ± 3 | 43.6 ± 3.2 | 0.8 |
| Cerebellum | 43.6 ± 1.8 | 44.6 ± 3.1 | 0.6 | 17.4 ± 1.3 | 19.3 ± 5.8 | 0.4 | 40.7 ± 2.1 | 38.1 ± 2.8 | 0.3 | 28 ± 2.1 | 30.4 ± 1.5 | 0.3 | 37.8 ± 3.7 | 35.9 ± 1.8 | 0.4 | 46.3 ± 3.2 | 45.8 ± 4 | 0.8 |
| SOD (unit/mg of protein) | ||||||||||||||||||
| Hippocampus | 28.2 ± 3.6 | 28.3 ± 0.5 | 0.9 | 9 ± 2.1 | 10.1 ± 0.4 | 0.546 | 21.1 ± 2.8 | 24.2 ± 1.8 | 0.1 | 15.8 ± 1.8 | 17.4 ± 2.8 | 0.4 | 20.7 ± 1 | 22.2 ± 0.8 | 0.4 | 30.5 ± 2.1 | 30.7 ± 4.3 | 0.9 |
| Prefrontal Cortex | 23.6 ± 3.2 | 18.7 ± 0.8 | 0.03 * | 8.6 ± 1.1 | 8 ± 1.8 | 0.772 | 17.9 ± 1.4 | 18.6 ± 4.4 | 0.7 | 12.1 ± 1.8 | 11.9 ± 1.4 | 0.9 | 18.4 ± 2.6 | 19.6 ± 1.4 | 0.5 | 23.5 ± 1.2 | 23.8 ± 5.4 | 0.9 |
| Cerebellum | 35.8 ± 4 | 32.4 ± 3.2 | 0.2 | 11.8 ± 2 | 13.6 ± 1.3 | 0.473 | 28.4 ± 6.9 | 33.8 ± 3.1 | 0.04 * | 20.4 ± 1.9 | 21.9 ± 3.3 | 0.5 | 27.9 ± 0.9 | 29 ± 1.5 | 0.6 | 37.5 ± 1.4 | 37.4 ± 2.5 | 0.9 |
| CAT (nmol/min/mg of protein) | ||||||||||||||||||
| Hippocampus | 20.9 ± 1.9 | 23.4 ± 1.1 | 0.121 | 12.4 ± 1.3 | 13.3 ± 1.2 | 0.5 | 20.6 ± 3.2 | 18.8 ± 1.3 | 0.2 | 15.5 ± 2.9 | 16.3 ± 2.1 | 0.6 | 19.5 ± 1.9 | 19.4 ± 1.1 | 0.9 | 22.4 ± 0.6 | 23.6 ± 1.8 | 0.4 |
| Prefrontal Cortex | 16 ± 0.8 | 16.7 ± 0.5 | 0.586 | 9.8 ± 0.6 | 9.7 ± 1.5 | 0.9 | 13.4 ± 0.4 | 15.3 ± 1.2 | 0.1 | 13.1 ± 0.9 | 13.3 ± 0.9 | 0.7 | 14.8 ± 1.1 | 14.9 ± 0.4 | 0.9 | 15.6 ± 3 | 18.4 ± 2.5 | 0.02 * |
| Cerebellum | 23.3 ± 1.6 | 25.5 ± 1.9 | 0.129 | 13.6 ± 1.7 | 14 ± 1.8 | 0.7 | 20.7 ± 1.3 | 21 ± 3.3 | 0.8 | 16.9 ± 1.5 | 18.5 ± 0.6 | 0.2 | 19.9 ± 0.4 | 21.4 ± 1.6 | 0.2 | 23.5 ± 0.9 | 26.8 ± 1.3 | 0.02 * |
| AChE (nmol/min/mg of protein) | ||||||||||||||||||
| Hippocampus | 428.6 ± 36.1 | 431.6 ± 20 | 0.933 | 638.3 ± 48 | 622.6 ± 44.4 | 0.664 | 450 ± 41.1 | 419 ± 46.5 | 0.392 | 544.6 ± 48.6 | 557 ± 67.9 | 0.7 | 433.3 ± 47.3 | 428 ± 43.4 | 0.8 | 427.3 ± 36.1 | 404.3 ± 22.5 | 0.5 |
| Prefrontal Cortex | 382 ± 9.1 | 334.3 ± 59.7 | 0.125 | 545.3 ± 68.8 | 584.3 ± 17.5 | 0.205 | 387.3 ± 37.1 | 351 ± 20.8 | 0.237 | 425.3 ± 5.8 | 399.3 ± 22.3 | 0.3 | 345.3 ± 22.5 | 387.3 ± 34.7 | 0.1 | 379.6 ± 13.4 | 326 ± 56.6 | 0.08 |
| Cerebellum | 425.6 ± 25.8 | 423 ± 1 | 0.944 | 630 ± 45.5 | 612.6 ± 26.5 | 0.647 | 452.3 ± 27.7 | 408.3 ± 40.5 | 0.251 | 520.3 ± 65.2 | 530.6 ± 86.6 | 0.7 | 439.6 ± 10.6 | 413.6 ± 54.5 | 0.4 | 341.6 ± 38.1 | 393.6 ± 52.6 | 0.1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakhal, M.M.; Jayaprakash, P.; Aburuz, S.; Sadek, B.; Akour, A. Canagliflozin Ameliorates Oxidative Stress and Autistic-like Features in Valproic-Acid-Induced Autism in Rats: Comparison with Aripiprazole Action. Pharmaceuticals 2023, 16, 769. https://doi.org/10.3390/ph16050769
Nakhal MM, Jayaprakash P, Aburuz S, Sadek B, Akour A. Canagliflozin Ameliorates Oxidative Stress and Autistic-like Features in Valproic-Acid-Induced Autism in Rats: Comparison with Aripiprazole Action. Pharmaceuticals. 2023; 16(5):769. https://doi.org/10.3390/ph16050769
Chicago/Turabian StyleNakhal, Mohammed Moutaz, Petrilla Jayaprakash, Salahdein Aburuz, Bassem Sadek, and Amal Akour. 2023. "Canagliflozin Ameliorates Oxidative Stress and Autistic-like Features in Valproic-Acid-Induced Autism in Rats: Comparison with Aripiprazole Action" Pharmaceuticals 16, no. 5: 769. https://doi.org/10.3390/ph16050769
APA StyleNakhal, M. M., Jayaprakash, P., Aburuz, S., Sadek, B., & Akour, A. (2023). Canagliflozin Ameliorates Oxidative Stress and Autistic-like Features in Valproic-Acid-Induced Autism in Rats: Comparison with Aripiprazole Action. Pharmaceuticals, 16(5), 769. https://doi.org/10.3390/ph16050769