
Exploring Cannabinoids with Enhanced Binding Affinity for Targeting the Expanded Endocannabinoid System: A Promising Therapeutic Strategy for Alzheimer’s Disease Treatment

 ,
,  , ,
, ,  , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
2.1. General Health Parameters
2.2. The Activation of the Expanded Endocannabinoid System Using Cannabinoid Receptor Ligands Reduced Anxiety-like Behavior and Partially Reversed the Cognitive Impairment in AβPP/PS1 Mice
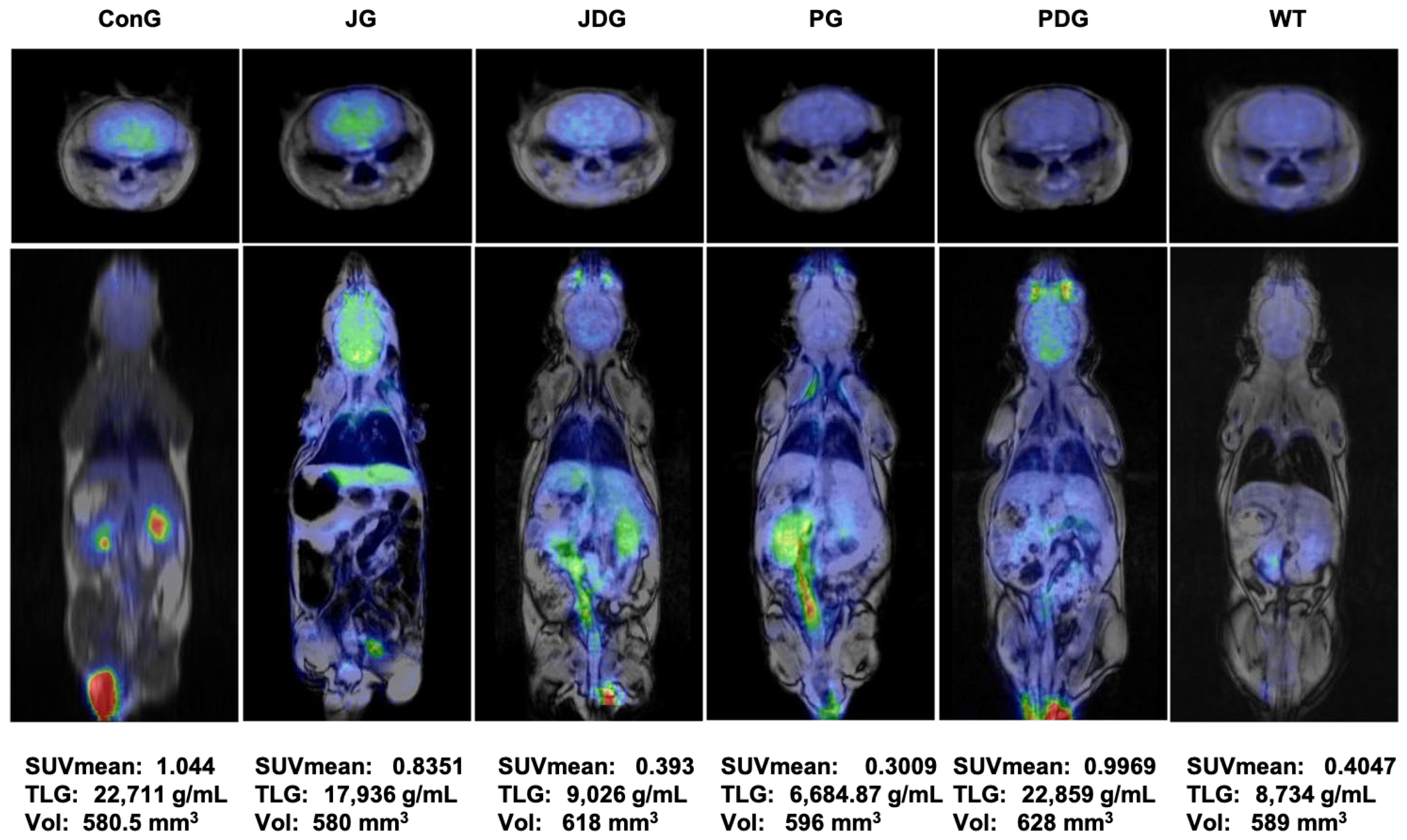
2.3. Prolonged Therapy with Selective CB2 Agonist JWH-133 or Cannabixir® Medium Flos Reduces Cerebral Glucose Metabolism in APP/PS1 Mice
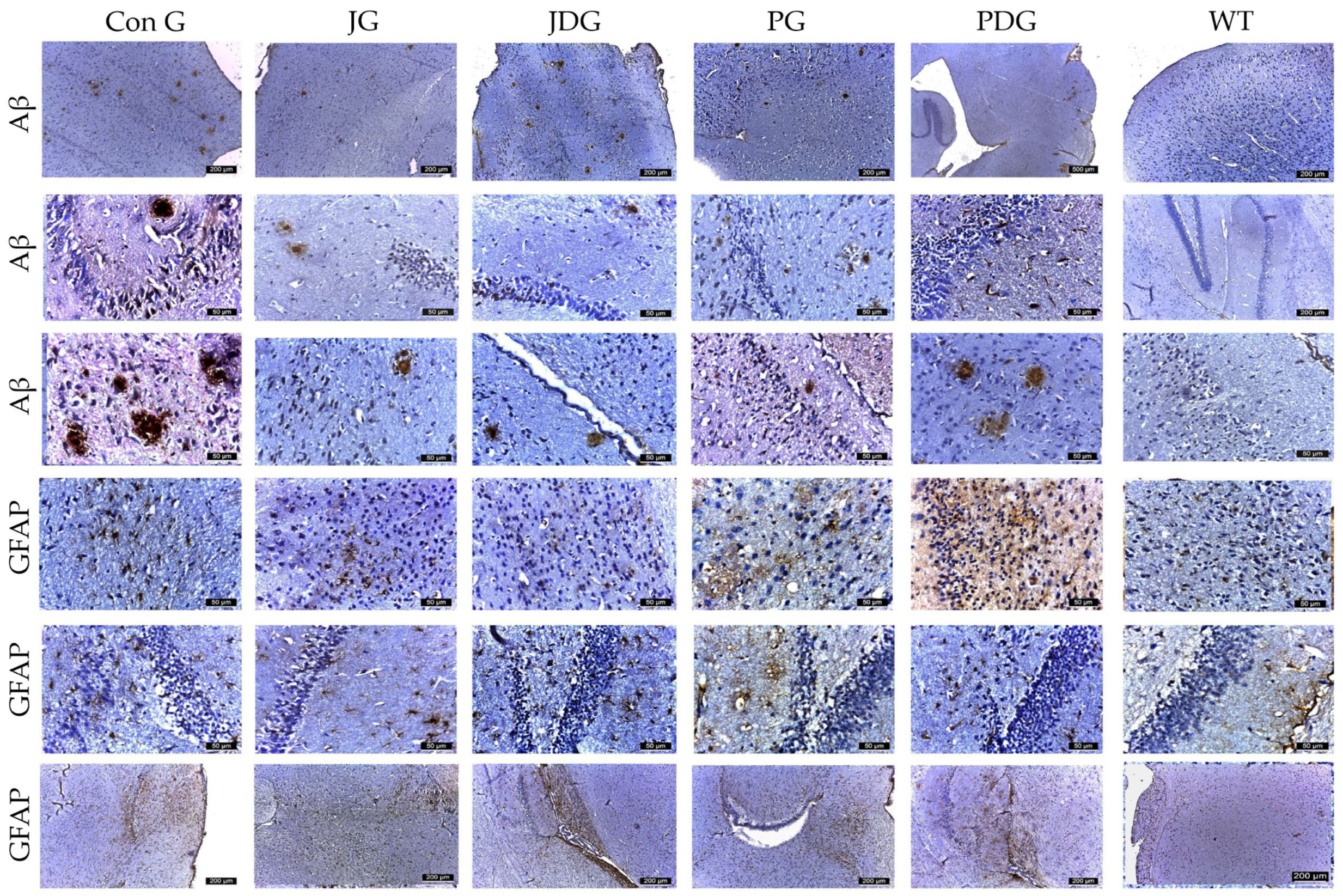
2.4. The Effect of Pharmacological Modulation of the ECS on AD-Related Aβ Burden, Neuroinflammation, and Glial Reactivity in APP/PS1 Mice
3. Discussion
4. Materials and Methods
4.1. Animal Care
4.2. Reagents and Pharmacological Treatment
4.3. Behavioral Evaluation of Cognitive Performance
4.4. PET-MRI
4.4.1. PET-MRI Acquisition and Reconstruction
4.4.2. PET-MRI Data Processing
4.5. Sample Collection, Histology, and Immunohistochemistry Analysis
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, S.; Huang, Y.; Yu, L.; Ji, X.; Wu, J. Current Neuropharmacology Send Orders for Reprints to [email protected] Impact of the Cannabinoid System in Alzheimer’s Disease. Curr. Neuropharmacol. 2023, 21, 715–726. [Google Scholar] [CrossRef] [PubMed]
- Ruver-Martins, A.C.; Bicca, M.A.; de Araujo, F.S.; de Noronha Sales Maia, B.H.L.; Pamplona, F.A.; da Silva, E.G.; Nascimento, F.P. Cannabinoid Extract in Microdoses Ameliorates Mnemonic and Nonmnemonic Alzheimer’s Disease Symptoms: A Case Report. J. Med. Case Rep. 2022, 16, 277. [Google Scholar] [CrossRef] [PubMed]
- Abate, G.; Uberti, D.; Tambaro, S. Potential and Limits of Cannabinoids in Alzheimer’s Disease Therapy. Biology 2021, 10, 542. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K. Cholinesterase Inhibitors as Alzheimer’s Therapeutics (Review). Mol. Med. Rep. 2019, 20, 1479–1487. [Google Scholar] [CrossRef] [PubMed]
- Marucci, G.; Buccioni, M.; Ben, D.D.; Lambertucci, C.; Volpini, R.; Amenta, F. Efficacy of Acetylcholinesterase Inhibitors in Alzheimer’s Disease. Neuropharmacology 2021, 190, 108352. [Google Scholar] [CrossRef]
- Tampi, R.; Gopalakrishna, G.; Gupta, A.; Zhu, F.; Zhu, J.; Shi, M.; Chu, F. Impact of Anti-Amyloid-β Monoclonal Antibodies on the Pathology and Clinical Profile of Alzheimer’s Disease: A Focus on Aducanumab and Lecanemab. Front. Aging Neurosci. 2022, 14, 870517. [Google Scholar] [CrossRef]
- Watt, J.A.; Isaranuwatchai, W.; Grossman, L.; Straus, S.E. Disease-Modifying Drugs for Alzheimer Disease: Implications for People in Canada. Can. Med. Assoc. J. 2023, 195, E146–E1448. [Google Scholar] [CrossRef] [PubMed]
- Frenkel, D. Alzheimer’s Disease: A Need for Personalized Therapeutic Approaches. Drug Dev. Res. 2020, 81, 141–143. [Google Scholar] [CrossRef] [PubMed]
- van der Flier, W.M.; de Vugt, M.E.; Smets, E.M.A.; Blom, M.; Teunissen, C.E. Towards a Future Where Alzheimer’s Disease Pathology Is Stopped before the Onset of Dementia. Nat. Aging 2023, 3, 494–505. [Google Scholar] [CrossRef] [PubMed]
- de Melo Reis, R.A.; Isaac, A.R.; Freitas, H.R.; de Almeida, M.M.; Schuck, P.F.; Ferreira, G.C.; Andrade-da-Costa, B.L.d.S.; Trevenzoli, I.H. Quality of Life and a Surveillant Endocannabinoid System. Front. Neurosci. 2021, 15, 747229. [Google Scholar] [CrossRef]
- Aso, E.; Sánchez-Pla, A.; Vegas-Lozano, E.; Maldonado, R.; Ferrer, I. Cannabis-Based Medicine Reduces Multiple Pathological Processes in AβPP/PS1 Mice. J. Alzheimer’s Dis. 2015, 43, 977–991. [Google Scholar] [CrossRef] [PubMed]
- Berry, A.J.; Zubko, O.; Reeves, S.J.; Howard, R.J. Endocannabinoid System Alterations in Alzheimer’s Disease: A Systematic Review of Human Studies. Brain Res. 2020, 1749, 147135. [Google Scholar] [CrossRef] [PubMed]
- Esposito, G.; De Filippis, D.; Steardo, L.; Scuderi, C.; Savani, C.; Cuomo, V.; Iuvone, T. CB1 Receptor Selective Activation Inhibits Beta-Amyloid-Induced INOS Protein Expression in C6 Cells and Subsequently Blunts Tau Protein Hyperphosphorylation in Co-Cultured Neurons. Neurosci. Lett. 2006, 404, 342–346. [Google Scholar] [CrossRef] [PubMed]
- Komorowska-Müller, J.A.; Schmöle, A.C. CB2 Receptor in Microglia: The Guardian of Self-Control. Int. J. Mol. Sci. 2021, 22, 19. [Google Scholar] [CrossRef]
- Veilleux, A.; Di Marzo, V.; Silvestri, C. The Expanded Endocannabinoid System/Endocannabinoidome as a Potential Target for Treating Diabetes Mellitus. Curr. Diabetes Rep. 2019, 19, 117. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.; Walder, K.; Kloiber, S.; Amminger, P.; Berk, M.; Bortolasci, C.C.; Maes, M.; Puri, B.K.; Carvalho, A.F. The Endocannabinoidome in Neuropsychiatry: Opportunities and Potential Risks. Pharmacol. Res. 2021, 170, 105729. [Google Scholar] [CrossRef]
- Oddo, S. The Role of MTOR Signaling in Alzheimer Disease. Front. Biosci. Sch. 2012, 4, 941–952. [Google Scholar] [CrossRef]
- Rapaka, D.; Bitra, V.R.; Reddy Challa, S.; Adiukwu, P.C. MTOR Signaling as a Molecular Target for the Alleviation of Alzheimer’s Disease Pathogenesis. Neurochem. Int. 2022, 155, 105311. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamilanban, T.; Alsayari, A.; Ramachawolran, G.; Wong, L.S.; Sekar, M.; Gan, S.H.; Subramaniyan, V.; Chinni, S.V.; Izzati Mat Rani, N.N.; et al. Trilateral Association of Autophagy, MTOR and Alzheimer’s Disease: Potential Pathway in the Development for Alzheimer’s Disease Therapy. Front. Pharmacol. 2022, 13, 1094351. [Google Scholar] [CrossRef]
- Chen, Z.R.; Huang, J.B.; Yang, S.L.; Hong, F.F. Role of Cholinergic Signaling in Alzheimer’s Disease. Molecules 2022, 27, 1816. [Google Scholar] [CrossRef] [PubMed]
- Thompson, K.J.; Tobin, A.B. Crosstalk between the M 1 Muscarinic Acetylcholine Receptor and the Endocannabinoid System: A Relevance for Alzheimer’s Disease? Cell Signal. 2020, 70, 109545. [Google Scholar] [CrossRef] [PubMed]
- Filipiuc, L.E.; Ştefănescu, R.; Solcan, C.; Ciorpac, M.; Szilagyi, A.; Cojocaru, D.; Stanciu, G.D.; Creangă, I.; Caratașu, C.C.; Ababei, D.C.; et al. Acute Toxicity and Pharmacokinetic Profile of an EU-GMP-Certified Cannabis sativa L. in Rodents. Pharmaceuticals 2023, 16, 694. [Google Scholar] [CrossRef] [PubMed]
- Shin, C.Y.; Kim, H.; Cha, K.; Won, D.H.; Lee, J.; Jang, S.W.; Sohn, U.D. The Effects of Donepezil, an Acetylcholinesterase Inhibitor, on Impaired Learning and Memory in Rodents. Biomol. Ther. 2018, 26, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Stanciu, G.D.; Ababei, D.C.; Solcan, C.; Bild, V.; Ciobica, A.; Beschea Chiriac, S.I.; Ciobanu, L.M.; Tamba, B.I. Preclinical Studies of Canagliflozin, a Sodium-Glucose Co-Transporter 2 Inhibitor, and Donepezil Combined Therapy in Alzheimer’s Disease. Pharmaceuticals 2023, 16, 1620. [Google Scholar] [CrossRef] [PubMed]
- Aso, E.; Juvés, S.; Maldonado, R.; Ferrer, I. CB2 Cannabinoid Receptor Agonist Ameliorates Alzheimer-like Phenotype in AβPP/PS1 Mice. J. Alzheimer’s Dis. 2013, 35, 847–858. [Google Scholar] [CrossRef] [PubMed]
- Martín-Moreno, A.M.; Brera, B.; Spuch, C.; Carro, E.; García-García, L.; Delgado, M.; Pozo, M.A.; Innamorato, N.G.; Cuadrado, A.; de Ceballos, M.L. Prolonged Oral Cannabinoid Administration Prevents Neuroinflammation, Lowers β-Amyloid Levels and Improves Cognitive Performance in Tg APP 2576 Mice. J. Neuroinflamm. 2012, 9, 8. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Bie, B.; Yang, H.; Xu, J.J.; Brown, D.L.; Naguib, M. Activation of the CB2 Receptor System Reverses Amyloid-Induced Memory Deficiency. Neurobiol. Aging 2013, 34, 791–804. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.; Zhang, J.; Fan, N.; Teng, Z.Q.; Wu, Y.; Yang, H.; Tang, Y.P.; Sun, H.; Song, Y.; Chen, C. XΔ9-THC-Caused Synaptic and Memory Impairments Are Mediated through COX-2 Signaling. Cell 2013, 155, 1154. [Google Scholar] [CrossRef] [PubMed]
- Filipiuc, L.E.; Ababei, D.C.; Alexa-Stratulat, T.; Pricope, C.V.; Bild, V.; Stefanescu, R.; Stanciu, G.D.; Tamba, B.I. Major Phytocannabinoids and Their Related Compounds: Should We Only Search for Drugs That Act on Cannabinoid Receptors? Pharmaceutics 2021, 13, 1823. [Google Scholar] [CrossRef] [PubMed]
- Fakhoury, M. Microglia and Astrocytes in Alzheimer’s Disease: Implications for Therapy. Curr. Neuropharmacol. 2017, 15, 508–518. [Google Scholar] [CrossRef] [PubMed]
- Giusti, V.; Kaur, G.; Giusto, E.; Civiero, L. Brain Clearance of Protein Aggregates: A Close-up on Astrocytes. Mol. Neurodegener. 2024, 19, 5. [Google Scholar] [CrossRef] [PubMed]
- Deture, M.A.; Dickson, D.W. The Neuropathological Diagnosis of Alzheimer’s Disease. Mol. Neurodegener. 2019, 5, 32. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.T.; Nguyen, T.D.; Nguyen, T.K.O.; Vo, T.K.; Vo, V.G. Advances in Developing Therapeutic Strategies for Alzheimer’s Disease. Biomed. Pharmacother. 2021, 139, 111623. [Google Scholar] [CrossRef] [PubMed]
- López, A.; Aparicio, N.; Pazos, M.R.; Grande, M.T.; Barreda-Manso, M.A.; Benito-Cuesta, I.; Vázquez, C.; Amores, M.; Ruiz-Pérez, G.; García-García, E.; et al. Cannabinoid CB 2 Receptors in the Mouse Brain: Relevance for Alzheimer’s Disease. J. Neuroinflamm. 2018, 15, 158. [Google Scholar] [CrossRef] [PubMed]
- Kamaruzzaman, M.A.; Romli, M.H.; Abas, R.; Vidyadaran, S.; Hidayat Baharuldin, M.T.; Nasaruddin, M.L.; Thirupathirao, V.; Sura, S.; Warsito, K.; Mohd Nor, N.H.; et al. Regulatory Role of the Endocannabinoid System on Glial Cells toward Cognitive Function in Alzheimer’s Disease: A Systematic Review and Meta-Analysis of Animal Studies. Front. Pharmacol. 2023, 14, 1053680. [Google Scholar] [CrossRef] [PubMed]
- Çakır, M.; Tekin, S.; Doğanyiğit, Z.; Erden, Y.; Soytürk, M.; Çiğremiş, Y.; Sandal, S. Cannabinoid Type 2 Receptor Agonist JWH-133, Attenuates Okadaic Acid Induced Spatial Memory Impairment and Neurodegeneration in Rats. Life Sci. 2019, 217, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Ivy, D.; Palese, F.; Vozella, V.; Fotio, Y.; Yalcin, A.; Ramirez, G.; Mears, D.; Wynn, G.; Piomelli, D. Cannabinoid CB 2 Receptors Mediate the Anxiolytic-like Effects of Monoacylglycerol Lipase Inhibition in a Rat Model of Predator-Induced Fear. Neuropsychopharmacology 2020, 45, 1330–1338. [Google Scholar] [CrossRef] [PubMed]
- Kasten, C.R.; Zhang, Y.; Boehm, S.L. Acute and Long-Term Effects of Δ9-Tetrahydrocannabinol on Object Recognition and Anxiety-like Activity Are Age-and Strain-Dependent in Mice. Pharmacol. Biochem. Behav. 2017, 163, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Cadoni, C.; Valentini, V.; Di Chiara, G. Behavioral Sensitization to Δ9-Tetrahydrocannabinol and Cross-Sensitization with Morphine: Differential Changes in Accumbal Shell and Core Dopamine Transmission. J. Neurochem. 2008, 106, 1586–1593. [Google Scholar] [CrossRef]
- Cohen, K.; Mama, Y.; Rosca, P.; Pinhasov, A.; Weinstein, A. Chronic Use of Synthetic Cannabinoids Is Associated with Impairment in Working Memory and Mental Flexibility. Front. Psychiatry 2020, 11, 00602. [Google Scholar] [CrossRef] [PubMed]
- Luo, F.; Rustay, N.R.; Ebert, U.; Hradil, V.P.; Cole, T.B.; Llano, D.A.; Mudd, S.R.; Zhang, Y.; Fox, G.B.; Day, M. Characterization of 7- and 19-Month-Old Tg2576 Mice Using Multimodal in Vivo Imaging: Limitations as a Translatable Model of Alzheimer’s Disease. Neurobiol. Aging 2012, 33, 933–944. [Google Scholar] [CrossRef] [PubMed]
- Poisnel, G.; Hérard, A.S.; El Tannir El Tayara, N.; Bourrin, E.; Volk, A.; Kober, F.; Delatour, B.; Delzescaux, T.; Debeir, T.; Rooney, T.; et al. Increased Regional Cerebral Glucose Uptake in an APP/PS1 Model of Alzheimer’s Disease. Neurobiol. Aging 2012, 33, 1995–2005. [Google Scholar] [CrossRef]
- Brendel, M.; Probst, F.; Jaworska, A.; Overhoff, F.; Korzhova, V.; Albert, N.L.; Beck, R.; Lindner, S.; Gildehaus, F.J.; Baumann, K.; et al. Glial Activation and Glucose Metabolism in a Transgenic Amyloid Mouse Model: A Triple-Tracer PET Study. J. Nucl. Med. 2016, 57, 954–960. [Google Scholar] [CrossRef] [PubMed]
- Rojas, S.; Herance, J.R.; Gispert, J.D.; Abad, S.; Torrent, É.; Jiménez, X.; Pareto, D.; Perpiña, U.; Sarroca, S.; Rodríguez, E.; et al. In Vivo Evaluation of Amyloid Deposition and Brain Glucose Metabolism of 5XFAD Mice Using Positron Emission Tomography. Neurobiol. Aging 2013, 34, 1790–1798. [Google Scholar] [CrossRef] [PubMed]
- Tolón, R.M.; Núñez, E.; Pazos, M.R.; Benito, C.; Castillo, A.I.; Antonio Martínez-Orgado, J.; Romero, J. The Activation of Cannabinoid CB 2 Receptors Stimulates in Situ and in Vitro Beta-Amyloid Removal by Human Macrophages. Brain Res. 2009, 1283, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Koppel, J.; Vingtdeux, V.; Marambaud, P.; d’Abramo, C.; Jimenez, H.; Stauber, M.; Friedman, R.; Davies, P. CB2 Receptor Deficiency Increases Amyloid Pathology and Alters Tau Processing in a Transgenic Mouse Model of Alzheimer’s Disease. Mol. Med. 2014, 20, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Escartin, C.; Galea, E.; Lakatos, A.; O’Callaghan, J.P.; Petzold, G.C.; Serrano-Pozo, A.; Steinhäuser, C.; Volterra, A.; Carmignoto, G.; Agarwal, A.; et al. Reactive Astrocyte Nomenclature, Definitions, and Future Directions. Nat. Neurosci. 2021, 24, 312–325. [Google Scholar] [CrossRef] [PubMed]
- Escartin, C.; Guillemaud, O.; Carrillo-de Sauvage, M.A. Questions and (Some) Answers on Reactive Astrocytes. Glia 2019, 67, 2221–2247. [Google Scholar] [CrossRef] [PubMed]
- Smit, T.; Deshayes, N.A.C.; Borchelt, D.R.; Kamphuis, W.; Middeldorp, J.; Hol, E.M. Reactive Astrocytes as Treatment Targets in Alzheimer’s Disease—Systematic Review of Studies Using the APPswePS1dE9 Mouse Model. Glia 2021, 69, 1852–1881. [Google Scholar] [CrossRef] [PubMed]
- Van Tijn, P.; Dennissen, F.J.A.; Gentier, R.J.G.; Hobo, B.; Hermes, D.; Steinbusch, H.W.M.; Van Leeuwen, F.W.; Fischer, D.F. Mutant Ubiquitin Decreases Amyloid β Plaque Formation in a Transgenic Mouse Model of Alzheimer’s Disease. Neurochem. Int. 2012, 61, 739–748. [Google Scholar] [CrossRef]
- Rodríguez, J.J.; Olabarria, M.; Chvatal, A.; Verkhratsky, A. Astroglia in Dementia and Alzheimer’s Disease. Cell Death Differ. 2008, 16, 378–385. [Google Scholar] [CrossRef] [PubMed]
- Ceyzériat, K.; Ben Haim, L.; Denizot, A.; Pommier, D.; Matos, M.; Guillemaud, O.; Palomares, M.A.; Abjean, L.; Petit, F.; Gipchtein, P.; et al. Modulation of Astrocyte Reactivity Improves Functional Deficits in Mouse Models of Alzheimer’s Disease. Acta Neuropathol. Commun. 2018, 6, 104. [Google Scholar] [CrossRef]
- Liddelow, S.A.; Barres, B.A. Reactive Astrocytes: Production, Function, and Therapeutic Potential. Immunity 2017, 46, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Sofroniew, M.V.; Vinters, H.V. Astrocytes: Biology and Pathology. Acta Neuropathol. 2010, 119, 7–35. [Google Scholar] [CrossRef] [PubMed]
- Navarrete, M.; Araque, A. Endocannabinoids Mediate Neuron-Astrocyte Communication. Neuron 2008, 57, 883–893. [Google Scholar] [CrossRef] [PubMed]
- Valori, C.F.; Possenti, A.; Brambilla, L.; Rossi, D. Challenges and Opportunities of Targeting Astrocytes to Halt Neurodegenerative Disorders. Cells 2021, 10, 2019. [Google Scholar] [CrossRef] [PubMed]
- Eraso-Pichot, A.; Pouvreau, S.; Olivera-Pinto, A.; Gomez-Sotres, P.; Skupio, U.; Marsicano, G. Endocannabinoid Signaling in Astrocytes. Glia 2023, 71, 44. [Google Scholar] [CrossRef] [PubMed]
- Haense, C.; Kalbe, E.; Herholz, K.; Hohmann, C.; Neumaier, B.; Krais, R.; Heiss, W.D. Cholinergic System Function and Cognition in Mild Cognitive Impairment. Neurobiol. Aging 2012, 33, 867–877. [Google Scholar] [CrossRef] [PubMed]
- Drummond, E.; Wisniewski, T. Alzheimer’s Disease: Experimental Models and Reality. Acta Neuropathol. 2016, 133, 155–175. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, M.; Fan, K.; Li, T.; Yan, T.; Wu, B.; Bi, K.; Jia, Y. Protective Effects of Alpinae Oxyphyllae Fructus Extracts on Lipopolysaccharide-Induced Animal Model of Alzheimer’s Disease. J. Ethnopharmacol. 2018, 217, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Lamarque, S.; Taghzouti, K.; Simon, H. Chronic Treatment with Δ9-Tetrahydrocannabinol Enhances the Locomotor Response to Amphetamine and Heroin. Implications for Vulnerability to Drug Addiction. Neuropharmacology 2001, 41, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Mouro, F.M.; Ribeiro, J.A.; Sebastião, A.M.; Dawson, N. Chronic, Intermittent Treatment with a Cannabinoid Receptor Agonist Impairs Recognition Memory and Brain Network Functional Connectivity. J. Neurochem. 2018, 147, 71–83. [Google Scholar] [CrossRef] [PubMed]
- Copeland, J.; Rooke, S.; Swift, W. Changes in Cannabis Use among Young People: Impact on Mental Health. Curr. Opin. Psychiatry 2013, 26, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Lovelace, J.W.; Corches, A.; Vieira, P.A.; Mackie, K.; Korzus, E. An Animal Model of Female Adolescent Cannabinoid Exposure Elicits a Long-Lasting Deficit in Presynaptic Long-Term Plasticity. Neuropharmacology 2015, 99, 242–255. [Google Scholar] [CrossRef] [PubMed]
- Magaki, S.; Hojat, S.A.; Wei, B.; So, A.; Yong, W.H. An Introduction to the Performance of Immunohistochemistry. Methods Mol. Biol. 2019, 1897, 289–298. [Google Scholar] [CrossRef] [PubMed]
- Guerin, C.J. Using Antibodies in Microscopy: A Guide to Immunohistochemistry. Part 2: IHC Staining Protocols. Microsc. Today 2023, 31, 34–39. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biomarkers | Experimental Groups | |||||
|---|---|---|---|---|---|---|
| ConG | JG | JDG | PG | PDG | WT | |
| GFAP H | ++ | ++ | ++ | ++++ | +++ | ++++ |
| GFAP SC | ++ | ++ | ++ | +++ | ++ | +++ |
| S100 H | ++ | ++ | ++ | ++++ | +++ | +++ |
| S100 SC | ++ | ++ | ++ | +++ | ++ | ++++ |
| M1 AChR H | + | +++ | +++ | +++ | ++ | ++++ |
| M1 AchR SC | + | +++ | ++ | ++ | +++ | ++++ |
| mTOR H | +++ | ++ | ++ | ++ | ++ | +++ |
| mTOR SC | ++++ | + | + | ++ | ++ | ++ |
| CB2 H | ++++ | ++ | ++ | +++ | +++ | + |
| CB2 SC | +++ | ++ | ++ | +++ | ++ | + |
| Primary Antibody | Dilution | Secondary Antibody | Dilution |
|---|---|---|---|
| Anti-beta amyloid antibody (ab201060), Abcam (Cambridge, UK) | 1:1000 | Goat anti-rabbit | 1:1000 |
| Anti-canabinoid receptor antibodies (ACR-002), Alomone Labs (Jerusalem, Israel) | 1:100 | Goat anti-rabbit | 1:100 |
| GFAP (SYSY cat. no. 173002) SYSY Antibodies (Göttingen, Germany) | 1:1000 | Goat anti-rabbit | 1:1000 |
| Beta S100 ca48942 sab antibody, Sigma-Aldrich (St. Louis, MO, USA) | 1:100 | Goat anti-mouse | 1:100 |
| Anti-mTOR (ab109268), Abcam (Cambridge, UK) | 1:100 | Goat anti-rabbit | 1:500 |
| CHRM1 polyclonal antibody (PA5-90876), Thermo Fisher Scientific (Waltham, MA, USA) | 1:100 | Goat anti-rabbit | 1:1000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stanciu, G.D.; Ababei, D.-C.; Solcan, C.; Uritu, C.-M.; Craciun, V.-C.; Pricope, C.-V.; Szilagyi, A.; Tamba, B.-I. Exploring Cannabinoids with Enhanced Binding Affinity for Targeting the Expanded Endocannabinoid System: A Promising Therapeutic Strategy for Alzheimer’s Disease Treatment. Pharmaceuticals 2024, 17, 530. https://doi.org/10.3390/ph17040530
Stanciu GD, Ababei D-C, Solcan C, Uritu C-M, Craciun V-C, Pricope C-V, Szilagyi A, Tamba B-I. Exploring Cannabinoids with Enhanced Binding Affinity for Targeting the Expanded Endocannabinoid System: A Promising Therapeutic Strategy for Alzheimer’s Disease Treatment. Pharmaceuticals. 2024; 17(4):530. https://doi.org/10.3390/ph17040530
Chicago/Turabian StyleStanciu, Gabriela Dumitrita, Daniela-Carmen Ababei, Carmen Solcan, Cristina-Mariana Uritu, Vlad-Constantin Craciun, Cosmin-Vasilica Pricope, Andrei Szilagyi, and Bogdan-Ionel Tamba. 2024. "Exploring Cannabinoids with Enhanced Binding Affinity for Targeting the Expanded Endocannabinoid System: A Promising Therapeutic Strategy for Alzheimer’s Disease Treatment" Pharmaceuticals 17, no. 4: 530. https://doi.org/10.3390/ph17040530
APA StyleStanciu, G. D., Ababei, D. -C., Solcan, C., Uritu, C. -M., Craciun, V. -C., Pricope, C. -V., Szilagyi, A., & Tamba, B. -I. (2024). Exploring Cannabinoids with Enhanced Binding Affinity for Targeting the Expanded Endocannabinoid System: A Promising Therapeutic Strategy for Alzheimer’s Disease Treatment. Pharmaceuticals, 17(4), 530. https://doi.org/10.3390/ph17040530