

Development of Syringaldehyde as an Agonist of the GLP-1 Receptor to Alleviate Diabetic Disorders in Animal Models
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
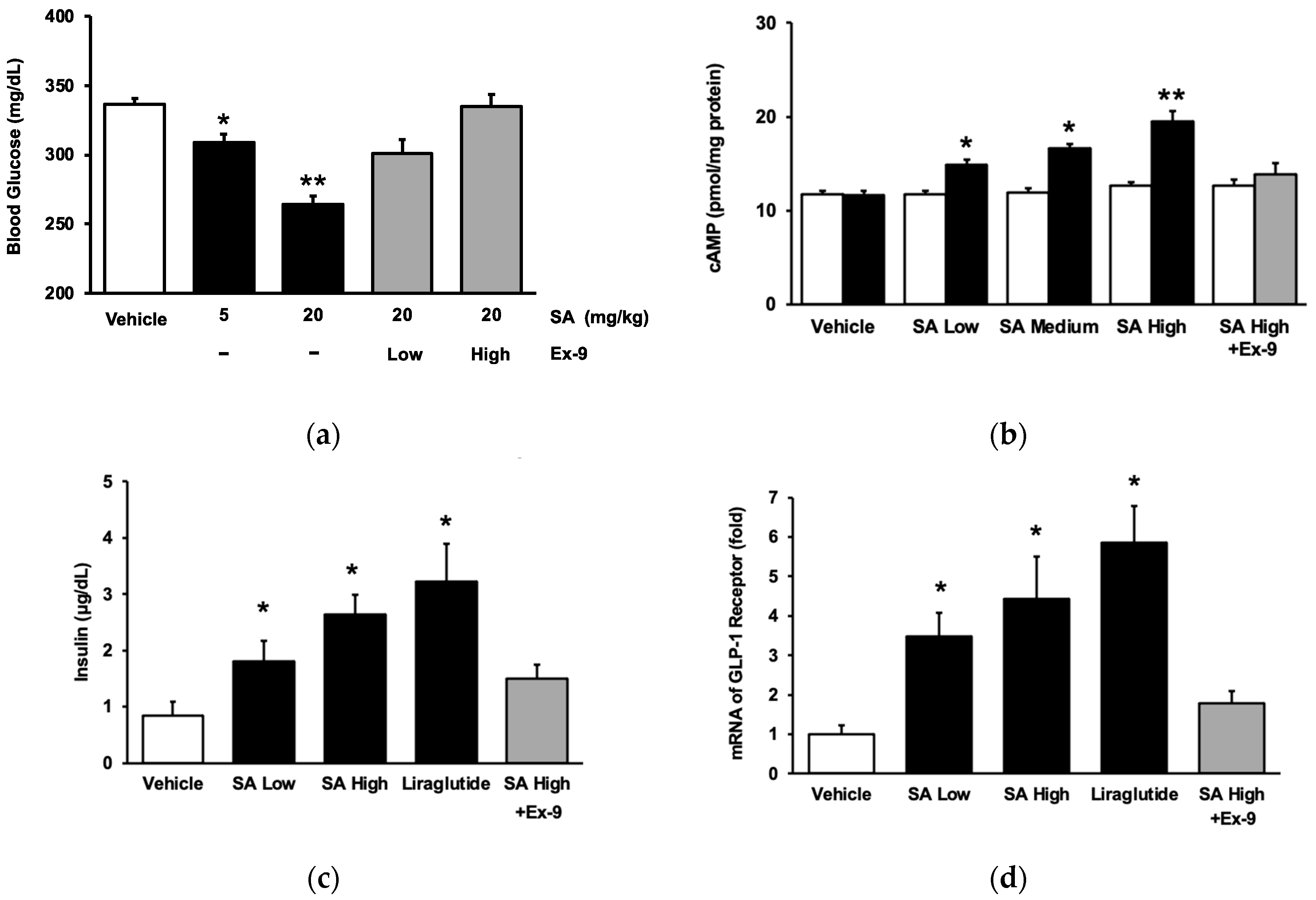
2.1. Activation of the GLP-1 Receptor Using SA
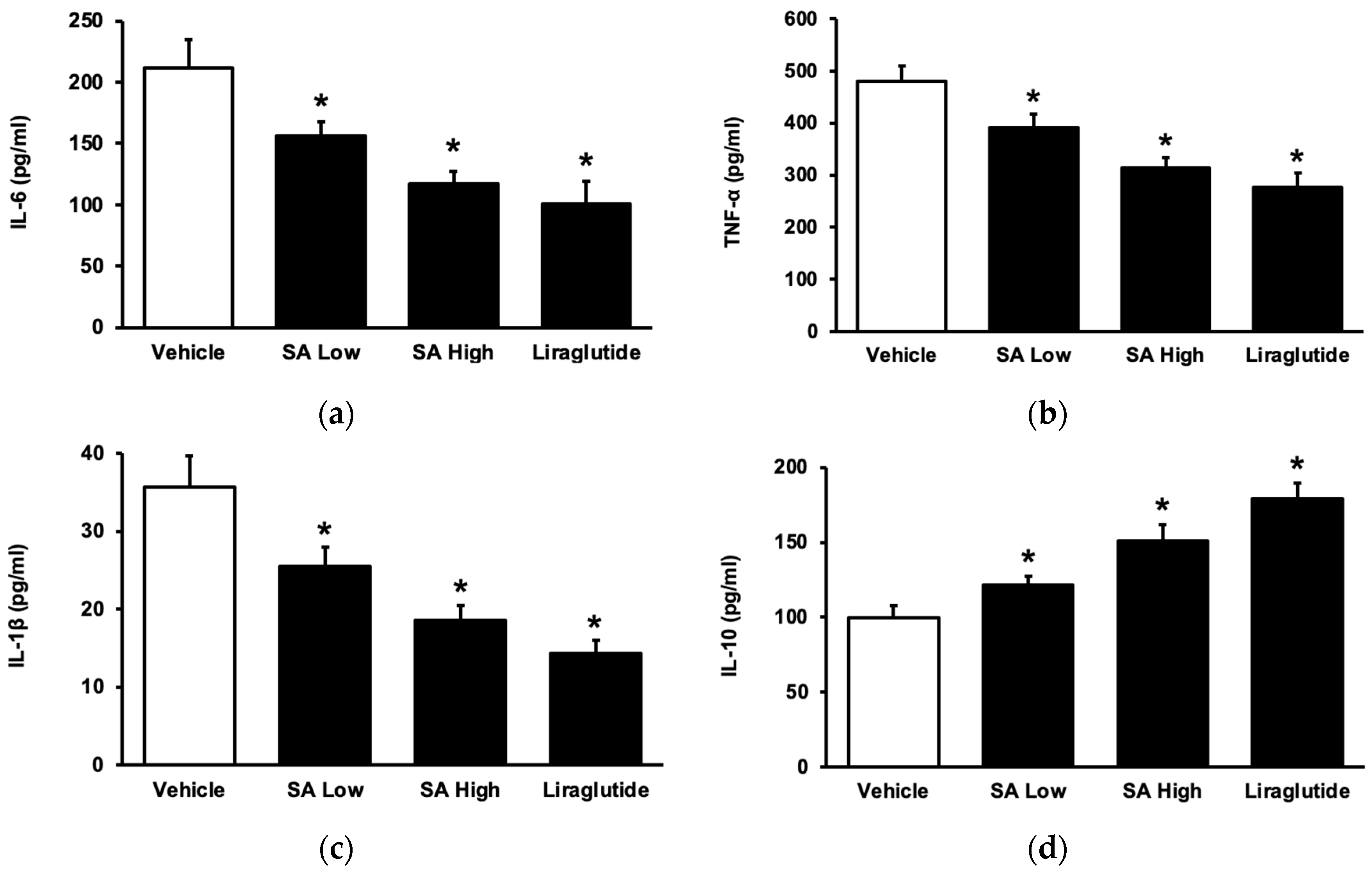
2.2. Inhibitory Effects of SA on Inflammation in Animals
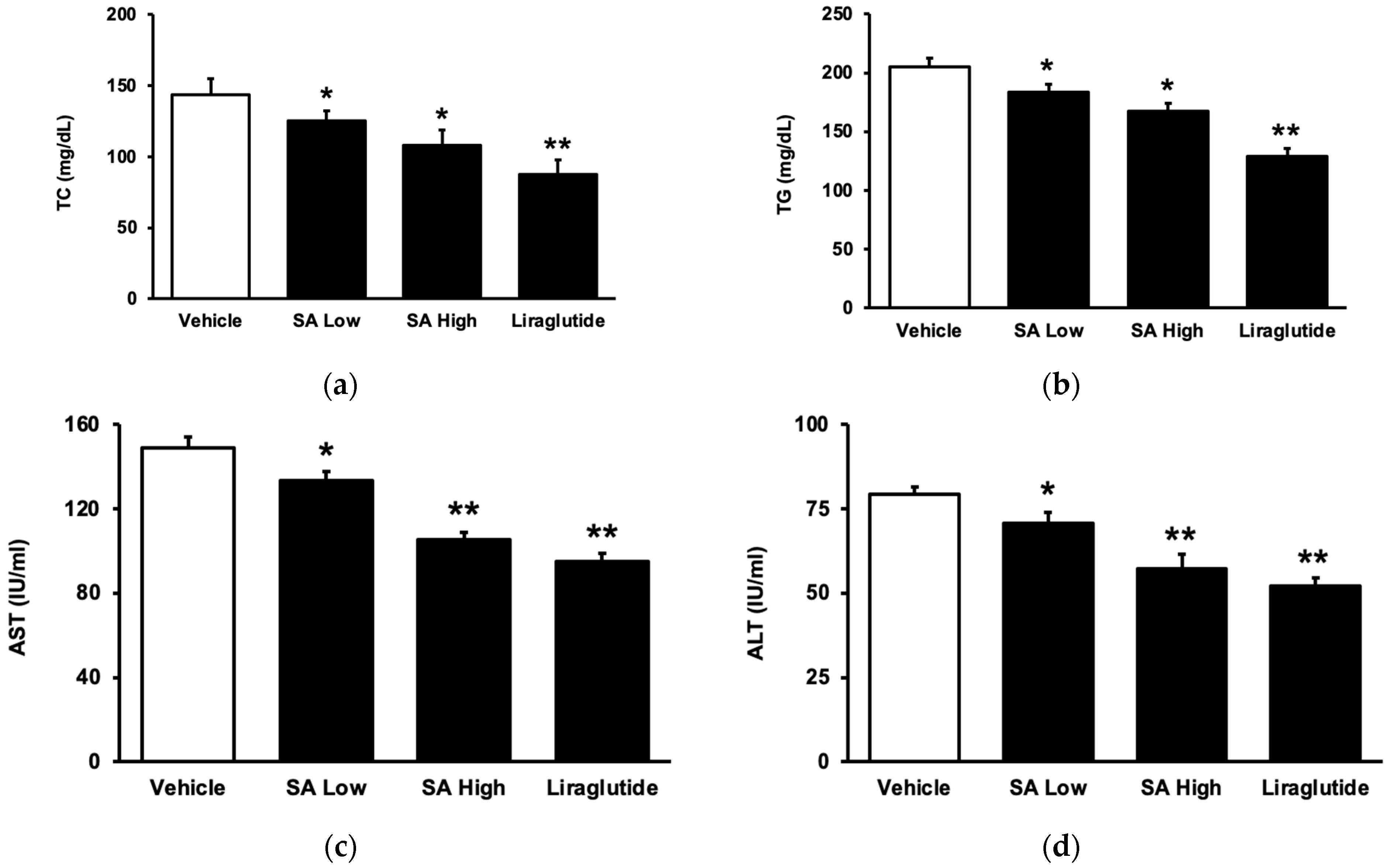
2.3. Inhibitory Effects of SA on Lipid Profiles and Hepatic Disorders in Animals
2.4. SA Improved Diabetic Nephropathy but Not the Body Weight in Animals
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animal Model
4.3. Experimental Protocol
4.4. Laboratory Determinations
4.5. The Cultured Cells
4.5.1. CHO-K1 Cells
4.5.2. Min-6 Cells
4.5.3. H9c2 Cells
4.6. Quantitative Reverse-Transcription Polymerase Chain Reaction (qRT-PCR)
- GLP-1 Receptor:
- forward: 5′-AGTGCGAAGAGTCCAAGCAA-3′
- reverse: 5′-TTGAGGGCAGCGTCTTTGAT-3′
- β-Actin:
- forward: 5′-CATCCAGGCTGTGTTGTCCC-3′
- reverse: 5′-CACGCACGATTTCCCTCTCA-3′
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nauck, M.A. Incretin-based therapies for type 2 diabetes mellitus: Properties, functions, and clinical implications. Am. J. Med. 2011, 124, S3–S18. [Google Scholar] [CrossRef] [PubMed]
- Alsalim, W.; Lindgren, O.; Ahren, B. Glucose-dependent insulinotropic polypeptide and glucagon-like peptide-1 secretion in humans: Characteristics and regulation. J. Diabetes Investig. 2023, 14, 354–361. [Google Scholar] [CrossRef] [PubMed]
- Seino, Y.; Yabe, D. Glucose-dependent insulinotropic polypeptide and glucagon-like peptide-1: Incretin actions beyond the pancreas. J. Diabetes Investig. 2013, 4, 108–130. [Google Scholar] [CrossRef]
- Waldrop, G.; Zhong, J.; Peters, M.; Rajagopalan, S. Incretin-Based Therapy for Diabetes: What a Cardiologist Needs to Know. J. Am. Coll. Cardiol. 2016, 67, 1488–1496. [Google Scholar] [CrossRef] [PubMed]
- Mori, H.; Okada, Y.; Arao, T.; Tanaka, Y. Sitagliptin improves albuminuria in patients with type 2 diabetes mellitus. J. Diabetes Investig. 2014, 5, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Nauck, M.A.; Meier, J.J. Pioneering oral peptide therapy for patients with type 2 diabetes. Lancet Diabetes Endocrinol. 2019, 7, 500–502. [Google Scholar] [CrossRef]
- Lingvay, I.; Hansen, T.; Macura, S.; Marre, M.; Nauck, M.A.; de la Rosa, R.; Woo, V.; Yildirim, E.; Wilding, J. Superior weight loss with once-weekly semaglutide versus other glucagon-like peptide-1 receptor agonists is independent of gastrointestinal adverse events. BMJ Open Diabetes Res. Care 2020, 8, e001706. [Google Scholar] [CrossRef]
- Thomas, M.K.; Nikooienejad, A.; Bray, R.; Cui, X.; Wilson, J.; Duffin, K.; Milicevic, Z.; Haupt, A.; Robins, D.A. Dual GIP and GLP-1 Receptor Agonist Tirzepatide Improves Beta-cell Function and Insulin Sensitivity in Type 2 Diabetes. J. Clin. Endocrinol. Metab. 2021, 106, 388–396. [Google Scholar] [CrossRef]
- Dutta, D.; Surana, V.; Singla, R.; Aggarwal, S.; Sharma, M. Efficacy and safety of novel twincretin tirzepatide a dual GIP and GLP-1 receptor agonist in the management of type-2 diabetes: A Cochrane meta-analysis. Indian. J. Endocrinol. Metab. 2021, 25, 475–489. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Hira, T.; Trakooncharoenvit, A.; Taguchi, H.; Hara, H. Improvement of Glucose Tolerance by Food Factors Having Glucagon-Like Peptide-1 Releasing Activity. Int. J. Mol. Sci. 2021, 22, 6623. [Google Scholar] [CrossRef] [PubMed]
- Stull, A.J. Blueberries’ Impact on Insulin Resistance and Glucose Intolerance. Antioxidants 2016, 5, 44. [Google Scholar] [CrossRef] [PubMed]
- Hui, H.; Tang, G.; Go, V.L. Hypoglycemic herbs and their action mechanisms. Chin. Med. 2009, 4, 11. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.X.; Xia, Z.N.; Gao, X.; Yin, F.; Liu, J.H. Glucagon-like peptide 1 receptor plays a critical role in geniposide-regulated insulin secretion in INS-1 cells. Acta Pharmacol. Sin. 2012, 33, 237–241. [Google Scholar] [CrossRef]
- Kuo, S.C.; Chung, H.H.; Huang, C.H.; Cheng, J.T. Decrease of hyperglycemia by syringaldehyde in diabetic rats. Horm. Metab. Res. 2014, 46, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Yao, D.; Yang, H.; Wei, Y.; Peng, Y.; Ding, Y.; Shu, L. Puerarin Protects Pancreatic beta-Cells in Obese Diabetic Mice via Activation of GLP-1R Signaling. Mol. Endocrinol. 2016, 30, 361–371. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, X.; Yi, X.; Liu, C.; Kong, D.; Zhang, J.; Gong, M. Myricetin: A potent approach for the treatment of type 2 diabetes as a natural class B GPCR agonist. FASEB J. 2017, 31, 2603–2611. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Wu, H.Y.; Liu, H.; Gong, N.; Wang, Y.R.; Wang, Y.X. Morroniside, a secoiridoid glycoside from Cornus officinalis, attenuates neuropathic pain by activation of spinal glucagon-like peptide-1 receptors. Br. J. Pharmacol. 2017, 174, 580–590. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Xing, H.J.; Wang, B.; Fu, C.; Zhang, Y.S.; Qiao, X.; Guo, C.; Zhang, X.L.; Hu, B.; Zhao, X.; et al. Cinchonine, a Potential Oral Small-Molecule Glucagon-Like Peptide-1 Receptor Agonist, Lowers Blood Glucose and Ameliorates Non-Alcoholic Steatohepatitis. Drug Des. Devel Ther. 2023, 17, 1417–1432. [Google Scholar] [CrossRef]
- Huang, C.H.; Chen, M.F.; Chung, H.H.; Cheng, J.T. Antihyperglycemic effect of syringaldehyde in streptozotocin-induced diabetic rats. J. Nat. Prod. 2012, 75, 1465–1468. [Google Scholar] [CrossRef]
- Weng, L.; Chen, T.H.; Zheng, Q.; Weng, W.H.; Huang, L.; Lai, D.; Fu, Y.S.; Weng, C.F. Syringaldehyde promoting intestinal motility with suppressing alpha-amylase hinders starch digestion in diabetic mice. Biomed. Pharmacother. 2021, 141, 111865. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Fu, Y.S.; Lin, K.; Huang, X.; Chen, Y.J.; Lai, D.; Kang, N.; Huang, L.; Weng, C.F. A narrative review: The pharmaceutical evolution of phenolic syringaldehyde. Biomed. Pharmacother. 2022, 153, 113339. [Google Scholar] [CrossRef] [PubMed]
- Esposito, K.; Nappo, F.; Marfella, R.; Giugliano, G.; Giugliano, F.; Ciotola, M.; Quagliaro, L.; Ceriello, A.; Giugliano, D. Inflammatory cytokine concentrations are acutely increased by hyperglycemia in humans: Role of oxidative stress. Circulation 2002, 106, 2067–2072. [Google Scholar] [CrossRef] [PubMed]
- Fujisaka, S.; Usui, I.; Bukhari, A.; Ikutani, M.; Oya, T.; Kanatani, Y.; Tsuneyama, K.; Nagai, Y.; Takatsu, K.; Urakaze, M.; et al. Regulatory mechanisms for adipose tissue M1 and M2 macrophages in diet-induced obese mice. Diabetes 2009, 58, 2574–2582. [Google Scholar] [CrossRef] [PubMed]
- Hogan, A.E.; Gaoatswe, G.; Lynch, L.; Corrigan, M.A.; Woods, C.; O’Connell, J.; O’Shea, D. Glucagon-like peptide 1 analogue therapy directly modulates innate immune-mediated inflammation in individuals with type 2 diabetes mellitus. Diabetologia 2014, 57, 781–784. [Google Scholar] [CrossRef] [PubMed]
- Tsalamandris, S.; Antonopoulos, A.S.; Oikonomou, E.; Papamikroulis, G.A.; Vogiatzi, G.; Papaioannou, S.; Deftereos, S.; Tousoulis, D. The Role of Inflammation in Diabetes: Current Concepts and Future Perspectives. Eur. Cardiol. 2019, 14, 50–59. [Google Scholar] [CrossRef] [PubMed]
- Elkattawy, H.A.; Elsherbini, D.M.; Ebrahim, H.A.; Abdullah, D.M.; Al-Zahaby, S.A.; Noser, Y.; Hassan, A.E. Rho-kinase inhibition ameliorates non-alcoholic fatty liver disease in type 2 diabetic rats. Physiol. Res. 2022, 71, 615–630. [Google Scholar] [CrossRef] [PubMed]
- Ruhl, C.E.; Everhart, J.E. Determinants of the association of overweight with elevated serum alanine aminotransferase activity in the United States. Gastroenterology 2003, 124, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Iacobellis, G.; Mohseni, M.; Bianco, S.D.; Banga, P.K. Liraglutide causes large and rapid epicardial fat reduction. Obesity 2017, 25, 311–316. [Google Scholar] [CrossRef]
- Verges, B.; Duvillard, L.; Pais de Barros, J.P.; Bouillet, B.; Baillot-Rudoni, S.; Rouland, A.; Sberna, A.L.; Petit, J.M.; Degrace, P.; Demizieux, L. Liraglutide Reduces Postprandial Hyperlipidemia by Increasing ApoB48 (Apolipoprotein B48) Catabolism and by Reducing ApoB48 Production in Patients With Type 2 Diabetes Mellitus. Arterioscler. Thromb. Vasc. Biol. 2018, 38, 2198–2206. [Google Scholar] [CrossRef]
- Tanaka, A.; Node, K. Evidence-based and tailored medication in type 2 diabetes: A pathway learned from clinical trials. Cardiovasc. Diabetol. 2019, 18, 19. [Google Scholar] [CrossRef]
- Nauck, M.A.; Meier, J.J. Management of endocrine disease: Are all GLP-1 agonists equal in the treatment of type 2 diabetes? Eur. J. Endocrinol. 2019, 181, R211–R234. [Google Scholar] [CrossRef]
- Umanath, K.; Lewis, J.B. Diabetic Kidney Disease: The Tiger May Have New Stripes. Am. J. Kidney Dis. 2018, 72, 631–633. [Google Scholar] [CrossRef]
- Warren, A.M.; Knudsen, S.T.; Cooper, M.E. Diabetic nephropathy: An insight into molecular mechanisms and emerging therapies. Expert. Opin. Ther. Targets 2019, 23, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Yuan, L.; Cao, S. Endogenous GLP-1 as a key self-defense molecule against lipotoxicity in pancreatic islets. Int. J. Mol. Med. 2015, 36, 173–185. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.H.; Han, J.H.; Park, H.S.; Lee, D.H.; Kim, S.J.; Cho, H.S.; Kang, J.S.; Myung, C.S. Effects of unaltered and bioconverted mulberry leaf extracts on cellular glucose uptake and antidiabetic action in animals. BMC Complement. Altern. Med. 2019, 19, 55. [Google Scholar] [CrossRef]
- Li, Y.X.; Cheng, K.C.; Liu, I.M.; Niu, H.S. Myricetin Increases Circulating Adropin Level after Activation of Glucagon-like Peptide 1 (GLP-1) Receptor in Type-1 Diabetic Rats. Pharmaceuticals 2022, 15, 173. [Google Scholar] [CrossRef]
- Lee, Y.S.; Jun, H.S. Anti-Inflammatory Effects of GLP-1-Based Therapies beyond Glucose Control. Mediators Inflamm. 2016, 2016, 3094642. [Google Scholar] [CrossRef] [PubMed]
- Lumeng, C.N.; Bodzin, J.L.; Saltiel, A.R. Obesity induces a phenotypic switch in adipose tissue macrophage polarization. J. Clin. Invest. 2007, 117, 175–184. [Google Scholar] [CrossRef]
- Bruen, R.; Curley, S.; Kajani, S.; Crean, D.; O’Reilly, M.E.; Lucitt, M.B.; Godson, C.G.; McGillicuddy, F.C.; Belton, O. Liraglutide dictates macrophage phenotype in apolipoprotein E null mice during early atherosclerosis. Cardiovasc. Diabetol. 2017, 16, 143. [Google Scholar] [CrossRef]
- Parthsarathy, V.; Holscher, C. Chronic treatment with the GLP1 analogue liraglutide increases cell proliferation and differentiation into neurons in an AD mouse model. PLoS ONE 2013, 8, e58784. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Lin, C.; Cheng, Y.; Zhuo, X.; Li, Q.; Xu, W.; Zhao, L.; Yang, L. Liraglutide Alleviates Hepatic Steatosis and Liver Injury in T2MD Rats via a GLP-1R Dependent AMPK Pathway. Front. Pharmacol. 2020, 11, 600175. [Google Scholar] [CrossRef] [PubMed]
- Verges, B.; Duvillard, L.; Pais de Barros, J.P.; Bouillet, B.; Baillot-Rudoni, S.; Rouland, A.; Petit, J.M.; Degrace, P.; Demizieux, L. Liraglutide Increases the Catabolism of Apolipoprotein B100-Containing Lipoproteins in Patients with Type 2 Diabetes and Reduces Proprotein Convertase Subtilisin/Kexin Type 9 Expression. Diabetes Care 2021, 44, 1027–1037. [Google Scholar] [CrossRef] [PubMed]
- Martinovic, N.; Abramovic, H.; Poklar Ulrih, N. Inhibition of copper-induced lipid peroxidation by sinapic acid and its derivatives in correlation to their effect on the membrane structural properties. Biochim. Biophys. Acta Biomembr. 2019, 1861, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, S.; Mateen, S.; Kausar, T.; Naeem, S.S.; Hasan, A.; Abidi, M.; Nayeem, S.M.; Faizy, A.F.; Moin, S. Effect of syringic acid and syringaldehyde on oxidative stress and inflammatory status in peripheral blood mononuclear cells from patients of myocardial infarction. Naunyn Schmiedebergs Arch. Pharmacol. 2020, 393, 691–704. [Google Scholar] [CrossRef] [PubMed]
- Perreault, L.; Skyler, J.S.; Rosenstock, J. Novel therapies with precision mechanisms for type 2 diabetes mellitus. Nat. Rev. Endocrinol. 2021, 17, 364–377. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.A.; Mells, J.; Dunham, R.M.; Grakoui, A.; Handy, J.; Saxena, N.K.; Anania, F.A. Glucagon-like peptide-1 receptor is present on human hepatocytes and has a direct role in decreasing hepatic steatosis in vitro by modulating elements of the insulin signaling pathway. Hepatology 2010, 51, 1584–1592. [Google Scholar] [CrossRef]
- Barriere, D.A.; Noll, C.; Roussy, G.; Lizotte, F.; Kessai, A.; Kirby, K.; Belleville, K.; Beaudet, N.; Longpre, J.M.; Carpentier, A.C.; et al. Combination of high-fat/high-fructose diet and low-dose streptozotocin to model long-term type-2 diabetes complications. Sci. Rep. 2018, 8, 424. [Google Scholar] [CrossRef]
- Bifari, F.; Manfrini, R.; Dei Cas, M.; Berra, C.; Siano, M.; Zuin, M.; Paroni, R.; Folli, F. Multiple target tissue effects of GLP-1 analogues on non-alcoholic fatty liver disease (NAFLD) and non-alcoholic steatohepatitis (NASH). Pharmacol. Res. 2018, 137, 219–229. [Google Scholar] [CrossRef]
- Imprialos, K.P.; Stavropoulos, K.; Doumas, M. Liraglutide and Renal Outcomes in Type 2 Diabetes. N. Engl. J. Med. 2017, 377, 2196. [Google Scholar] [CrossRef]
- Mali, N.; Su, F.; Ge, J.; Fan, W.X.; Zhang, J.; Ma, J. Efficacy of liraglutide in patients with diabetic nephropathy: A meta-analysis of randomized controlled trials. BMC Endocr. Disord. 2022, 22, 93. [Google Scholar] [CrossRef]
- Song, T.; Jia, Y.; Li, Z.; Wang, F.; Ren, L.; Chen, S. Effects of Liraglutide on Nonalcoholic Fatty Liver Disease in Patients with Type 2 Diabetes Mellitus: A Systematic Review and Meta-Analysis. Diabetes Ther. 2021, 12, 1735–1749. [Google Scholar] [CrossRef] [PubMed]
- Raun, K.; von Voss, P.; Gotfredsen, C.F.; Golozoubova, V.; Rolin, B.; Knudsen, L.B. Liraglutide, a long-acting glucagon-like peptide-1 analog, reduces body weight and food intake in obese candy-fed rats, whereas a dipeptidyl peptidase-IV inhibitor, vildagliptin, does not. Diabetes 2007, 56, 8–15. [Google Scholar] [CrossRef]
- Hsu, T.M.; Hahn, J.D.; Konanur, V.R.; Lam, A.; Kanoski, S.E. Hippocampal GLP-1 receptors influence food intake, meal size, and effort-based responding for food through volume transmission. Neuropsychopharmacology 2015, 40, 327–337. [Google Scholar] [CrossRef] [PubMed]
- Bozkurt, A.A.; Mustafa, G.; Tarik, A.; Adile, O.; Murat, S.H.; Mesut, K.; Yildiray, K.; Coskun, S.; Murat, C. Syringaldehyde exerts neuroprotective effect on cerebral ischemia injury in rats through anti-oxidative and anti-apoptotic properties. Neural Regen. Res. 2014, 9, 1884–1890. [Google Scholar] [CrossRef]
- Le, T.D.V.; Fathi, P.; Watters, A.B.; Ellis, B.J.; Besing, G.K.; Bozadjieva-Kramer, N.; Perez, M.B.; Sullivan, A.I.; Rose, J.P.; Baggio, L.L.; et al. Fibroblast growth factor-21 is required for weight loss induced by the glucagon-like peptide-1 receptor agonist liraglutide in male mice fed high carbohydrate diets. Mol. Metab. 2023, 72, 101718. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wu, Q.; Chen, Y.; Gu, G.; Gao, R.; Peng, B.; Wang, Y.; Li, A.; Guo, J.; Xu, X.; et al. Updated Pharmacological Effects, Molecular Mechanisms, and Therapeutic Potential of Natural Product Geniposide. Molecules 2022, 27, 3319. [Google Scholar] [CrossRef] [PubMed]
- Niisato, N.; Marunaka, Y. Therapeutic potential of multifunctional myricetin for treatment of type 2 diabetes mellitus. Front. Nutr. 2023, 10, 1175660. [Google Scholar] [CrossRef]
- Yang, H.Z.; Dong, R.; Jia, Y.; Li, Y.; Luo, G.; Li, T.; Long, Y.; Liang, S.; Li, S.; Jin, X.; et al. Morroniside ameliorates glucocorticoid-induced osteoporosis and promotes osteoblastogenesis by interacting with sodium-glucose cotransporter 2. Pharm. Biol. 2023, 61, 416–426. [Google Scholar] [CrossRef]
- Furman, B.L. Streptozotocin-Induced Diabetic Models in Mice and Rats. Curr. Protoc. Pharmacol. 2015, 70, 5.47.1–5.47.20. [Google Scholar] [CrossRef]
- Udumula, M.P.; Mangali, S.; Kalra, J.; Dasari, D.; Goyal, S.; Krishna, V.; Bollareddy, S.R.; Sriram, D.; Dhar, A.; Bhat, A. High fructose and streptozotocin induced diabetic impairments are mitigated by Indirubin-3-hydrazone via downregulation of PKR pathway in Wistar rats. Sci. Rep. 2021, 11, 12924. [Google Scholar] [CrossRef] [PubMed]
- Niu, C.S.; Chen, L.J.; Niu, H.S. Antihyperglycemic action of rhodiola-aqeous extract in type1-like diabetic rats. BMC Complement. Altern. Med. 2014, 14, 20. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Cheng, K.C.; Liu, I.M.; Cheng, J.T. Identification of Andrographolide as an Agonist of Bile Acid TGR5 Receptor in a Cell Line to Demonstrate the Reduction in Hyperglycemia in Type-1 Diabetic Rats. Pharmaceuticals 2023, 16, 1417. [Google Scholar] [CrossRef]
- Liu, I.M.; Chang, C.K.; Juang, S.W.; Kou, D.H.; Tong, Y.C.; Cheng, K.C.; Cheng, J.T. Role of hyperglycaemia in the pathogenesis of hypotension observed in type-1 diabetic rats. Int. J. Exp. Pathol. 2008, 89, 292–300. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.; Li, Y.; Cheng, J.-T.; Liu, I.-M.; Cheng, K.-C. Development of Syringaldehyde as an Agonist of the GLP-1 Receptor to Alleviate Diabetic Disorders in Animal Models. Pharmaceuticals 2024, 17, 538. https://doi.org/10.3390/ph17040538
Lee J, Li Y, Cheng J-T, Liu I-M, Cheng K-C. Development of Syringaldehyde as an Agonist of the GLP-1 Receptor to Alleviate Diabetic Disorders in Animal Models. Pharmaceuticals. 2024; 17(4):538. https://doi.org/10.3390/ph17040538
Chicago/Turabian StyleLee, Jenpei, Yingxiao Li, Juei-Tang Cheng, I-Min Liu, and Kai-Chun Cheng. 2024. "Development of Syringaldehyde as an Agonist of the GLP-1 Receptor to Alleviate Diabetic Disorders in Animal Models" Pharmaceuticals 17, no. 4: 538. https://doi.org/10.3390/ph17040538
APA StyleLee, J., Li, Y., Cheng, J. -T., Liu, I. -M., & Cheng, K. -C. (2024). Development of Syringaldehyde as an Agonist of the GLP-1 Receptor to Alleviate Diabetic Disorders in Animal Models. Pharmaceuticals, 17(4), 538. https://doi.org/10.3390/ph17040538