2.1. List of CPP5s
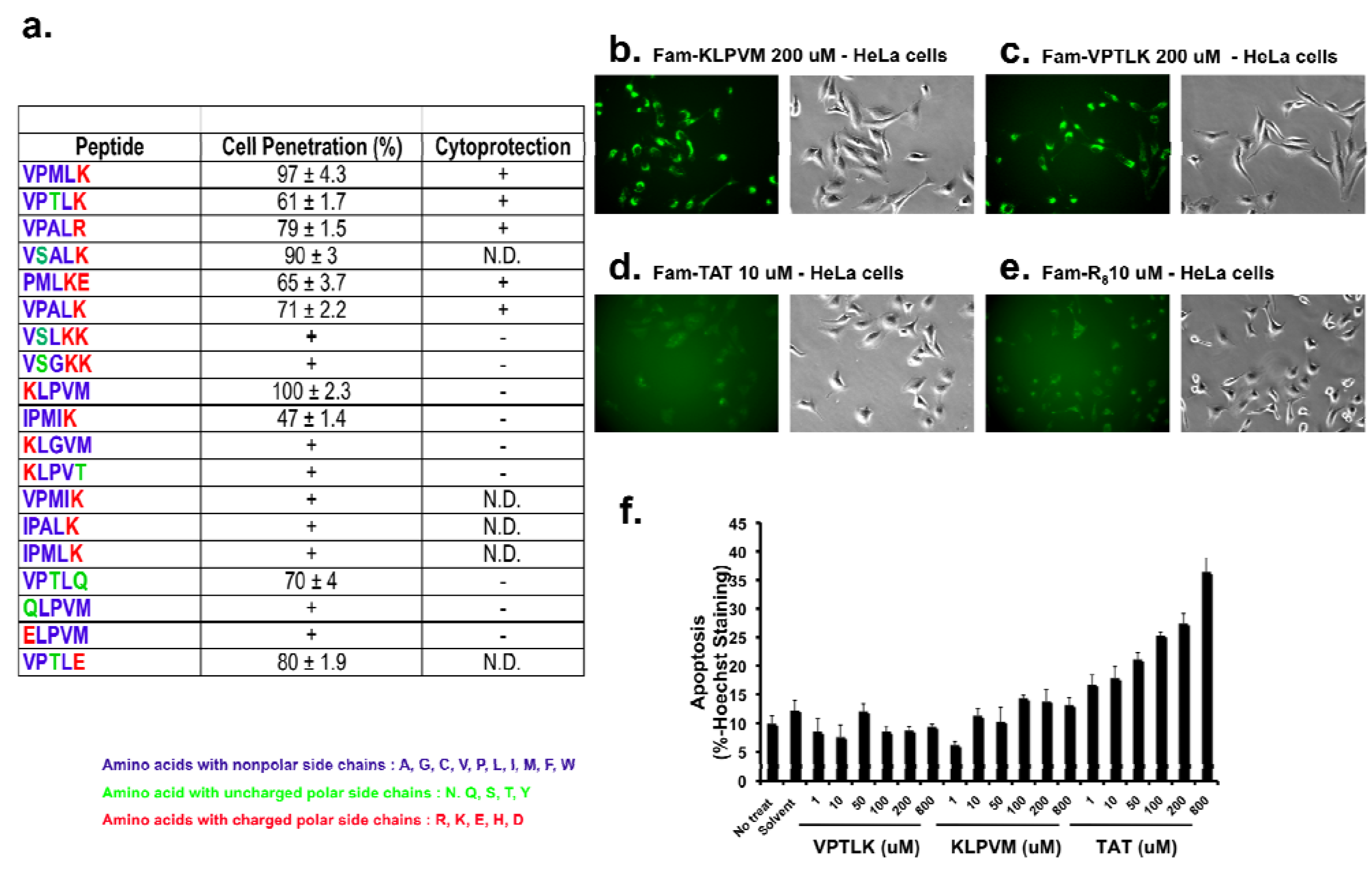
The amino acid sequences of characterized CPP5s are shown in
Figure 1a. CPP5s that were designed based on the Bax-binding domain of Ku70 of several species are named Bax-Inhibiting Peptides (BIPs); they are VPMLK, PMLKE, VPTLK, VPALR, and VPALK. VPMLK and PMLKE were designed from human Ku70, and the same sequence exists in the Ku70 homologues of dog and monkey. VPTLK and VPALR are based on mouse and rat Ku70, respectively. VPALK was designed from cattle Ku70, and the same sequence exists in Ku70 of the African clawed frog. Cytoprotective activities of these BIPs have been previously reported, both in cell culture and animal models [
6,
22,
23,
25,
27,
28]. Among the CPP5s, KLPVM was found to have the best cell-penetrating activity when assessed by flow cytometric analysis of the cellular fluorescence intensity of Fam (5,6-carboxyfluorescein)-labeled CPP5s (
Figure 1a). The observed variation in cell-entering activity of Fam-labeled-CPP5s is shown in
Figure 1a as a percentage of the activity of KLPVM. In all cases, Fam was attached to the N-terminal amino acid. For some CPP5s, cell-penetration activity was confirmed by detection of an intracellular fluorescent signal by microscopy. These latter results are indicated with ‘+’ symbols, and a quantitative comparison with the activity of KLPVM was not performed. Representative images of the cell entry of Fam-labeled KLPVM and VPTLK are shown in
Figure 1b and c. As positive controls, cells cultured in the presence of Fam-TAT and Fam-octa-arginine (R8) are also shown (
Figure 1d and
Figure 1e). As previously reported [
6,
29], Fam-labeled CPP5s showed detectable cell-penetrating activity when the peptides were added to the culture medium at concentrations greater than 1 µM. In this study, we used KLPVM and VPTLK as representative CPP5s for further analysis of the cell-penetrating activity.
2.3. Mechanism of cell entry of CPP5s
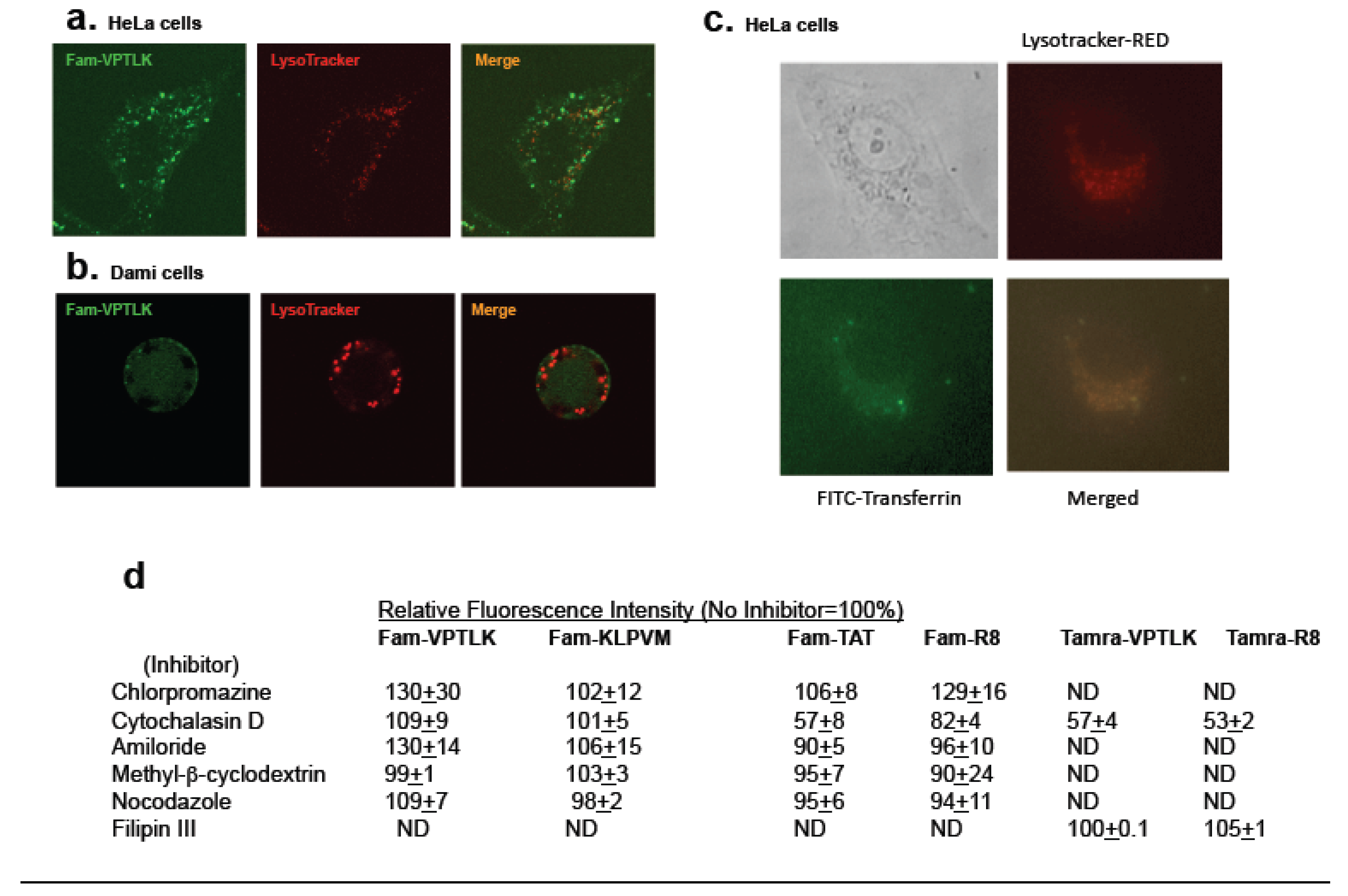
Lysosomes are known to contain molecules which have entered the cell through pinocytosis [
30]. To determine whether CPP5s use pinocytotic pathways for their cell entry, the localizations of Fam-labeled CPP5s and lysosomes (labeled by LysoTracker® Red) were compared. Only a subset of Fam-VPTLK peptides co-localized with lysosomes while the majority did not (
Figure 2a and b). In contrast, FITC-labeled transferrin and LysoTracker-Red co-localized extensively, which is consistent with the understanding that transferrin enters cells through receptor-mediated endocytosis (
Figure 2c). Partial co-localization of Fam-VPTLK with LysoTracker suggests that the cell penetration of CPP5s is achieved through both pinocytosis-dependent and -independent pathways.
To further examine the role of pinocytosis, various pinocytosis inhibitors were tested as shown in
Figure 2d. HeLa cells were incubated with each inhibitor in serum-free DMEM for 30 min, followed by incubation for 1 h with Fam-labeled CPP5 peptides (30 μM). Then, the cells were extensively washed with Hanks’ Buffered Salt Solution (HBSS) and complete DMEM, prior to the detection of fluorescence by microscopy and FACS. None of amiloride, chlorpromazine, cytochalasin D, Filipin III, nocodazole and methyl-beta-cyclodextrin blocked the entry of Fam-labeled VPTLK and KLPVM into HeLa cells. However, when Tamra-labeled VPTLK was used, cytochalasin D significantly attenuated the cell entry of VPTLK (57±4% of the non-treated control). As a control experiment to validate the effectiveness of these inhibitors, FITC-transferrin (for chlorpromazine) and rhodamine-labeled EGF (for cytochalasin D and amiloride) were used. These inhibitors significantly suppressed the cell entry of transferrin and EGF, as HeLa cells were unable to take up detectable FITC-transferrin and rhodamine-EGF (not shown). To compare the cell-penetrating mechanism between CPP5s and arginine-rich CPPs, the uptake of TAT and R8 was also examined in the presence of these inhibitors. Only cytochalasin D had a suppressive effect on the cell-penetrating activity of TAT and R8, consistent with a previous report [
31].
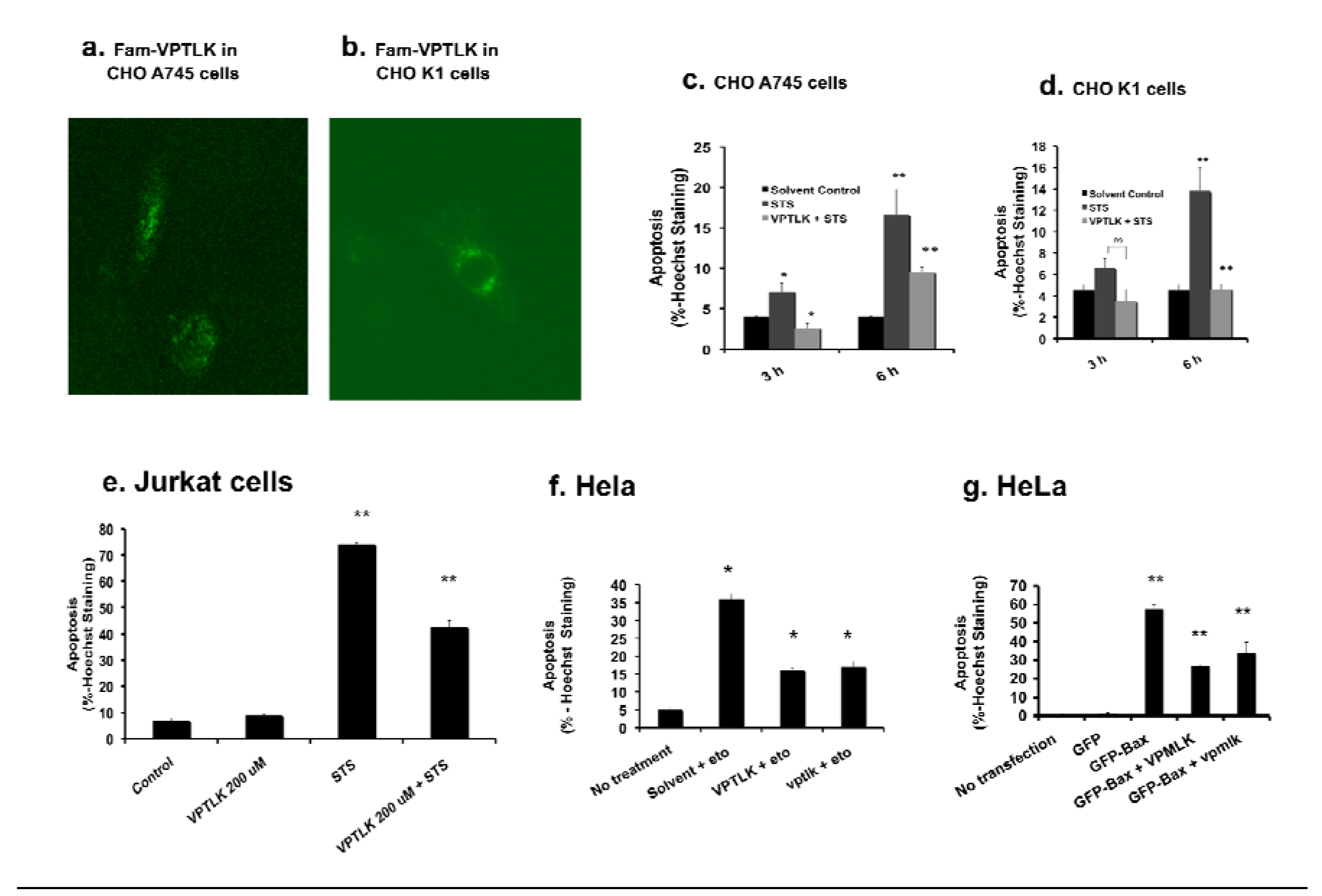
Chinese hamster ovary (CHO) A745 cells lack proteoglycans in the plasma membrane [
32], and this cell line has been used to determine whether proteoglycans are required for CPP entry [
32]. In CHO A745, as well as in its parental cell line CHO K1 that expresses proteoglycans, Fam-VPTLK and Fam-KLPVM (not shown) both showed cell-penetrating activity (
Figure 3a and b), and VPTLK showed cytoprotective activity [
i.e., VPTLK protected these cells from the apoptosis-inducing chemical staurosporine (STS)] (
Figure 3c and d). The cell-penetrating activity of CPP5s was also examined in Jurkat cells that are reported to lack caveolin-mediated endocytotic activity [
33]. Fam-VPTLK and -KLPVM were able to enter Jurkat cells (not shown) and VPTLK showed cytoprotective activity in this cell type (
Figure 3e). These results suggest that CPP5s do not require proteoglycan- and caveolin-mediated endocytosis for their cell-penetrating activity.
Figure 2.
Mechanism of cell entry of CPP5s. a and b. VPTLK partially co-localizes with lysosomes. Microscopic analysis of the co-localization of Fam-VPTLK with lysosome-staining dye (LysoTracker™ red DND-99, L7528 Invitrogen). Hela cells (a) and DAMI cells (b) were incubated with Fam-VPTLK (50 μM) and LysoTracker (100 nM) for 1 h at 37 °C. c. FITC-labeled Transferin (FITC-Transferin) co-localized with lysosome. HeLa cells were incubaled with FITC-transferin (100 nM) and LysoTracker (100 nM) for 1 h at 37 °C. d. Effects of inhibitors of pinocytosis and endocytosis on the cell-penetrating activity of CPP5s. HeLa cells were pre-treated with various pinocytosis inhibitors for 30 min. Then, Fam- or Tamra-labeled peptides were added to the culture medium. Cells were incubated with peptides for 1 h and the cellular fluorescence intensity was measured by fluorescence microscopy equipped with image analysis software (for Fam-labeled peptides) or FACS (for Tamra-labeled peptides) as described in Experimental Procedures. VPTLK and KLPVM were used at 30 μM and TAT and RRRRRRRR (R8) were used at 10 μM. The results are shown as percentages of the fluorescence intensity of inhibitor-treated cells (the intensity of non-treated cells is designated as 100%). Each value represents mean of three experiments and standard error. The pinocytosis inhibitors used were: Chlorpromazine (an inhibitor of chlathrin-mediated endocytosis), cytochalasin D (an inhibitor of F-actin elongation, resulting in inhibition of macropinocytosis and caveolin-mediated endocytosis), amiloride (an inhibitor of macropinocytosis), methyl beta cyclodextrin (it depletes cholesterol from the plasma membrane, thus inhibiting cell entry dependent on lipid rafts), nocodazole (an inhibitor of microtubule formation), and Filipin III (an inhibitor of macropinocytosis).
Figure 2.
Mechanism of cell entry of CPP5s. a and b. VPTLK partially co-localizes with lysosomes. Microscopic analysis of the co-localization of Fam-VPTLK with lysosome-staining dye (LysoTracker™ red DND-99, L7528 Invitrogen). Hela cells (a) and DAMI cells (b) were incubated with Fam-VPTLK (50 μM) and LysoTracker (100 nM) for 1 h at 37 °C. c. FITC-labeled Transferin (FITC-Transferin) co-localized with lysosome. HeLa cells were incubaled with FITC-transferin (100 nM) and LysoTracker (100 nM) for 1 h at 37 °C. d. Effects of inhibitors of pinocytosis and endocytosis on the cell-penetrating activity of CPP5s. HeLa cells were pre-treated with various pinocytosis inhibitors for 30 min. Then, Fam- or Tamra-labeled peptides were added to the culture medium. Cells were incubated with peptides for 1 h and the cellular fluorescence intensity was measured by fluorescence microscopy equipped with image analysis software (for Fam-labeled peptides) or FACS (for Tamra-labeled peptides) as described in Experimental Procedures. VPTLK and KLPVM were used at 30 μM and TAT and RRRRRRRR (R8) were used at 10 μM. The results are shown as percentages of the fluorescence intensity of inhibitor-treated cells (the intensity of non-treated cells is designated as 100%). Each value represents mean of three experiments and standard error. The pinocytosis inhibitors used were: Chlorpromazine (an inhibitor of chlathrin-mediated endocytosis), cytochalasin D (an inhibitor of F-actin elongation, resulting in inhibition of macropinocytosis and caveolin-mediated endocytosis), amiloride (an inhibitor of macropinocytosis), methyl beta cyclodextrin (it depletes cholesterol from the plasma membrane, thus inhibiting cell entry dependent on lipid rafts), nocodazole (an inhibitor of microtubule formation), and Filipin III (an inhibitor of macropinocytosis).
![Pharmaceuticals 03 03594 g002]()
Figure 3.
Protection of cells from apoptosis by VPTLK. a-d. Proteoglycan is not required for VPTLK cell entry. Microscopic analysis of cell entry of Fam-VPTLK in Chinese Hamster Ovary (CHO) A745 cells lacking proteoglycans in the plasma membrane (a) and CHO K1 cells that express proteoglycans (b). CHO A745 and K1 cells were incubated with Fam-VPTLK (200 μM) for 3 h at 37 °C. VPTLK (3 h pre-incubation) protected both cell lines from 100 nM staurosporine (STS) treatment (3, and 6 h) (c and d). Apoptosis was determined by using Hoechst dye nuclear staining. Each bar represents the mean of triplicate samples and standard error, and statistical significance was determined by an unpaired student t test; “ns”: not significant, *P < 0.05, and **P < 0.01. e. VPTLK protected Jurkat cells from STS-induced apoptosis. Jurkat cells were pre-incubated with VPTLK 200 μM for 3 h prior to STS treatment (100 nM, 3 or 6 h). Apoptosis was determined by Hoechst dye nuclear staining. Each bar represents the mean of triplicate samples and standard error, and statistical significance was determined by an unpaired student t test: **P < 0.01. f. VPTLK and D-type VPTLK (vptlkD) protected HeLa cells from etoposide-induced apoptosis. HeLa cells were pretreated with 200 μM VPTLK or vptlkD for 3 h prior to etoposide treatment (10 μM, 24 h). Apoptosis was determined by Hoechst dye nuclear staining. Each bar represents the mean of triplicate samples and standard error. ** P < 0.01 g. VPTLK and vptlkD inhibited Bax-induced apoptosis in HeLa cells. HeLa cells were pretreated with VPTLK and vptlkD 200 μM for 3 h prior to pEGFP-Bax plasmid transfection. Apoptosis was assayed 24 h after the transfection as described in Experimental Procedures. Each bar represents the mean of triplicate samples and standard error. ** P < 0.01
Figure 3.
Protection of cells from apoptosis by VPTLK. a-d. Proteoglycan is not required for VPTLK cell entry. Microscopic analysis of cell entry of Fam-VPTLK in Chinese Hamster Ovary (CHO) A745 cells lacking proteoglycans in the plasma membrane (a) and CHO K1 cells that express proteoglycans (b). CHO A745 and K1 cells were incubated with Fam-VPTLK (200 μM) for 3 h at 37 °C. VPTLK (3 h pre-incubation) protected both cell lines from 100 nM staurosporine (STS) treatment (3, and 6 h) (c and d). Apoptosis was determined by using Hoechst dye nuclear staining. Each bar represents the mean of triplicate samples and standard error, and statistical significance was determined by an unpaired student t test; “ns”: not significant, *P < 0.05, and **P < 0.01. e. VPTLK protected Jurkat cells from STS-induced apoptosis. Jurkat cells were pre-incubated with VPTLK 200 μM for 3 h prior to STS treatment (100 nM, 3 or 6 h). Apoptosis was determined by Hoechst dye nuclear staining. Each bar represents the mean of triplicate samples and standard error, and statistical significance was determined by an unpaired student t test: **P < 0.01. f. VPTLK and D-type VPTLK (vptlkD) protected HeLa cells from etoposide-induced apoptosis. HeLa cells were pretreated with 200 μM VPTLK or vptlkD for 3 h prior to etoposide treatment (10 μM, 24 h). Apoptosis was determined by Hoechst dye nuclear staining. Each bar represents the mean of triplicate samples and standard error. ** P < 0.01 g. VPTLK and vptlkD inhibited Bax-induced apoptosis in HeLa cells. HeLa cells were pretreated with VPTLK and vptlkD 200 μM for 3 h prior to pEGFP-Bax plasmid transfection. Apoptosis was assayed 24 h after the transfection as described in Experimental Procedures. Each bar represents the mean of triplicate samples and standard error. ** P < 0.01
![Pharmaceuticals 03 03594 g003]()
Previously, D-type amino acid-based peptides were shown to have better stability inside cells because they are resistant to proteases [
34]. Therefore, we expected that D-type VPTLK might show better cytoprotective activity because of its higher intracellular stability. Interestingly, both L-type and D-type VPTLK showed similar cytoprotective activity against etoposide treatment and Bax overexpression (
Figure 3f and g). These results suggest that the cell-entry pathway of CPP5s is not selective for L- or D-type amino acids.
2.4. Quantification of the cell-penetrating activity of VPTLK by mass spectrometric analysis
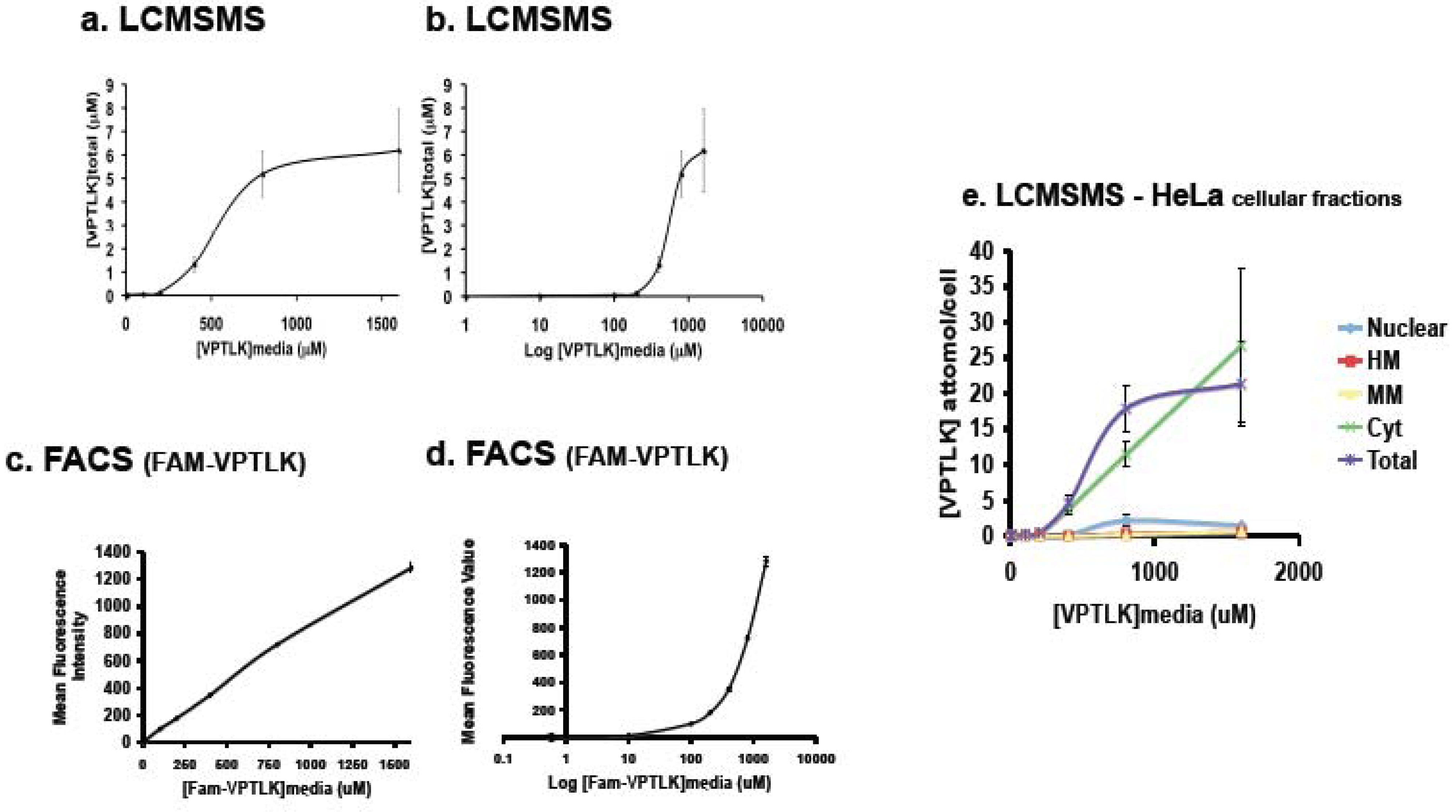
The amount of VPTLK that penetrates into HeLa cells was measured by using a mass spectrometric [LC-MS/MS (Liquid Chromatography Mass Spectrometer)] method. Isotopically labeled VPTLK [VPTL(
13C
15N)K] was added to the cell lysate at a known concentration prior to the peptide extraction steps so that VPTL(
13C
15N)K could be used as an internal standard. HeLa cells were cultured in medium containing concentrations of VPTLK ranging from 0.5 μM to 1.6 mM. The amount of VPTLK that entered into HeLa cells increased in a dose-dependent manner (
Figure 4a and b). However, it reached an intracellular plateau of approximately 6 μM when the VPTLK concentration in the culture medium was greater than 800 μM. These results suggest that the cell entry of VPTLK is not merely by a simple diffusion mechanism. Interestingly, the cell entry of Fam-VPTLK showed a linear increase up to the maximum concentration examined (1.6 mM) (
Figure 4c and d), indicating that Fam-VPTLK and VPTLK use different cell-entry pathways, at least when the extracellular VPTLK concentration is above 800 μM.
Cells were subjected to subcellular fractionation, and the distribution of VPTLK in each fraction was determined (
Figure 4e). VPTLK localized mainly in the cytosol and did not accumulate in the nucleus, nor in the heavy membrane fraction (HM) that contains mitochondria, nor in the endoplasmic reticulum (ER). The cytosolic localization of VPTLK was also observed when fluorescence microscopy was used to detect Fam-VPTLK (
Figure 1c), which is consistent with the result of LC-MS/MS analysis of non-labeled VPTLK.
2.5. CPP5s have protein-transduction activity
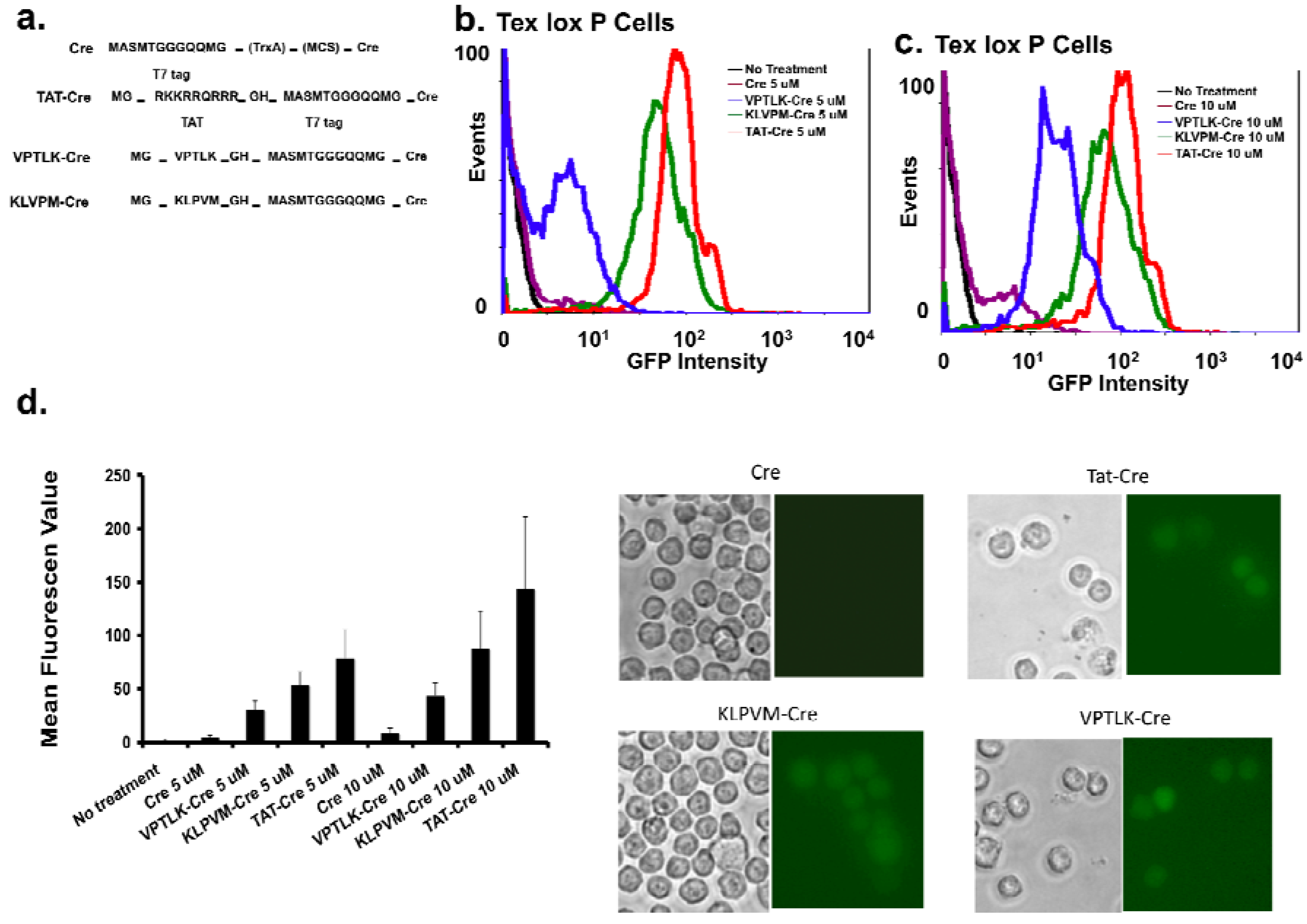
To determine the protein-transduction activity of VPTLK and KLPVM, these peptides were added to the N-terminus of Cre DNA recombinase and the resulting CPP5-Cre fusion proteins were added separately to the culture medium of Tex-loxP-EG cells, which are engineered to express GFP through Cre-dependent excision of a chromosomal lox P site. This method was developed by S. Dowdy’s group to determine the protein-transduction activity of TAT peptide [
11]. As shown in
Figure 5a, the TAT sequence of the TAT-Cre expression plasmid was replaced with VPTLK and KLPVM and these plasmids were used to express recombinant proteins.
Figure 4.
Quantification of VPTLK concentration in HeLa cells by mass spectrometry (MS). a and b. VPTLK concentration in HeLa cells (y-axis) incubated in various concentrations of VPTLK in the culture medium (x-axis). HeLa cells were incubated for 24 h in VPTLK-containing medium. Then the cells were collected and the VPTLK concentration was measured using MS as described in the Experimental Procedures. Each value represents the mean of triplicate samples and standard error. b. Semi-Logarithmic presentation of the data in (a). c and d. Fluorescence-Activated Cell Sorting (FACS) analysis of the cell penetration of Fam-VPTLK into HeLa cells. Cells were incubated for 3 h in various concentrations of VPTLK in the medium. Each value represents the mean of triplicate samples and standard error. d. Semi-Logarithmic profile of the data shown in (c). e. VPTLK concentration in each subcellular fraction of HeLa cells cultured in various concentrations of VPTLK. The graph shows the VPTLK concentration in the cytolsolic fraction (Cyt), heavy membrane (HM), nucleus, or microsomal membrane (MM). HeLa cells were incubated for 24 h with various concentrations of VPTLK, and subcellular fractions were analyzed by MS as described in Expeimental Procedures. Each value represents the mean of triplicate samples and standard error.
Figure 4.
Quantification of VPTLK concentration in HeLa cells by mass spectrometry (MS). a and b. VPTLK concentration in HeLa cells (y-axis) incubated in various concentrations of VPTLK in the culture medium (x-axis). HeLa cells were incubated for 24 h in VPTLK-containing medium. Then the cells were collected and the VPTLK concentration was measured using MS as described in the Experimental Procedures. Each value represents the mean of triplicate samples and standard error. b. Semi-Logarithmic presentation of the data in (a). c and d. Fluorescence-Activated Cell Sorting (FACS) analysis of the cell penetration of Fam-VPTLK into HeLa cells. Cells were incubated for 3 h in various concentrations of VPTLK in the medium. Each value represents the mean of triplicate samples and standard error. d. Semi-Logarithmic profile of the data shown in (c). e. VPTLK concentration in each subcellular fraction of HeLa cells cultured in various concentrations of VPTLK. The graph shows the VPTLK concentration in the cytolsolic fraction (Cyt), heavy membrane (HM), nucleus, or microsomal membrane (MM). HeLa cells were incubated for 24 h with various concentrations of VPTLK, and subcellular fractions were analyzed by MS as described in Expeimental Procedures. Each value represents the mean of triplicate samples and standard error.
![Pharmaceuticals 03 03594 g004]()
First, cells were cultured for 3 h in serum-free medium in the presence of 5 or 10 µM Cre, TAT-Cre, KLPVM-Cre, or VPTLK-Cre. GFP expression was measured 48 h after the incubation with the CPP-Cre proteins. As shown in
Figure 5b-d, KLPVM and VPTLK were able to induce GFP expression similar to TAT did, whereas Cre itself did not. In serum-free medium, KLPVM and TAT showed similar levels of protein-transduction activity, and VPTLK showed lower activity than the other CPPs. Second, Cre fusion proteins (5 µM) were added to the culture medium containing serum (10%), and GFP expression was examined 48 h after the treatment (
Figure 5 e). All of TAT-Cre, VPTLK-Cre, and KLPVM-Cre were able to induce GFP expression, suggesting that all these CPPs are able to deliver the cargo protein in 10% serum-containing medium. We also sought to examine the addition of 10 μM Cre-fusion proteins to serum-containing medium; however, precipitation of serum proteins occurred and cell viability became very low under these conditions. Probably, the high-salt solubilization buffer (10 mM phosphate and 500 mM NaCl, pH 6.8) of the fusion proteins caused the serum proteins to precipitate.
2.6. Discussion
The present study showed that two CPP5s (KLPVM and VPTLK) have protein-transduction activity, as determined by the delivery of Cre protein into cells from the culture medium. Previously, we reported that these CPP5s were able to deliver GFP from the medium into the cell [
6]. CPP5 was attached to the N-terminus of Cre (
Figure 5 a) and to the C-terminus of GFP [
6]. Both cargo proteins (i.e. Cre and GFP) were delivered into the cell. Therefore, CPP5 is able to express protein transduction activity whether at the N- or C-terminus of the cargo protein. In this study, we focused on analyzing VPTLK and KLPVM for their protein-transduction activities, and it is not yet known whether other CPP5s have such capabilities. Further studies examining the protein-transduction activities of other CPP5s are necessary to find the best CPP5s for protein delivery across the plasma membrane.
Cationic peptides such as TAT and poly-arginine have been widely used as a means for protein transduction [
10,
11,
19,
35]. As shown in
Figure 1, CPP5s have less toxicity than arginine-rich peptide. The low toxicity of CPP5s may be an advantage over TAT in terms of delivery of large amounts of therapeutic cargo molecules into damaged cells to restore their function. Examination of the protein-transduction activity of CPP5s in animal models is clearly the next important experiment to be undertaken.
VPTLK localizes mostly in the cytosol. Since VPTLK binds cytosolic Bax protein [
6,
22,
23], Bax may serve as the cytosolic retention factor for VPTLK. It is also possible that other VPTLK-binding factors exist in the cytosol. Importantly, VPTLK-Cre was able to switch on Cre-inducible GFP gene expression, indicating that VPTLK-Cre was able to reach chromosomal DNA in the nucleus. This means that VPTLK did not interfere with the nuclear-localization signal of Cre protein. Therefore, VPTLK can be utilized for the delivery of nuclear proteins into the cell even though VPTLK itself has a propensity to localize in the cytosol.
By means of a mass spectrometry-based method, we succeeded in performing a quantitative analysis of the cell-penetrating activity of VPTLK. We found that VPTLK reached a concentration of 6 μM in the cytosol when the culture medium contained 800 μM VPTLK (
Figure 4 e); however, the intracellular concentration of VPTLK did not increase further even after the concentration of VPTLK in the medium was increased from 800 μM to 1.6 mM. The intracellular VPTLK concentration showed a sigmoid curve in relation to the extracellular VPTLK concentration. This result suggests that simple diffusion is not the mechanism of VPTLK entry into the cell, but that there is a specific cell-entry pathway for VPTLK such as a receptor-mediated mechanism. In contrast, the intracellular concentration of Fam-VPTLK did not reach a plateau, suggesting that Fam-labeled VPTLK and non-labeled VPTLK use different mechanisms to enter the cell, at least at high concentrations (>800 μM) in the culture medium.
Figure 5.
Protein-Transduction activity (PTDs) of VPTLK and KLPVM determined by the delivery of Cre recombinase into cells.
a. Scheme of Cre protein, TAT-Cre, VPTLK-Cre, and KLPVM-Cre examined in this study.
b-e. Tex-loxP-EG cells were incubated with purified recombinant Cre, VPTLK-Cre, KLPVM-Cre, and TAT-Cre (5 and 10 μM) for 3 h in serum-free
(b-d) or 10% serum-containing
(e) media. After the incubation of cells with CPP-Cre fusion proteins, medium was changed to 10% FCS containing DMEM and cells were culture for additional 45 hrs. The delivery of Cre into Tex-lox P-EG cells was evaluated by GFP expression as previously reported [
11]. GFP expression was analyzed 48 h after the incubation of cells with Cre proteins.
b and c. FACS analysis of GFP expression in cells incubated with 5 μM
(b) or 10 μM
(c) Cre and CPP-Cre fusion proteins in serum-free medium. Representative data from three independent experiments are shown.
d. Mean GFP fluorescence intensity of Tex-loxP-EG cells incubated with 5 or 10 μM Cre recombinant proteins under serum-free conditions. Each bar represents three independent experiments and standard error.
e. Tex-loxP-EG cells incubated with purified recombinant Cre, VPTLK-Cre, KLPVM-Cre, and TAT-Cre (5 μM) in 10% serum-containing medium for 3 hrs. Images were captured 48 h after the incubation to detect GFP expression (right panel).
Figure 5.
Protein-Transduction activity (PTDs) of VPTLK and KLPVM determined by the delivery of Cre recombinase into cells.
a. Scheme of Cre protein, TAT-Cre, VPTLK-Cre, and KLPVM-Cre examined in this study.
b-e. Tex-loxP-EG cells were incubated with purified recombinant Cre, VPTLK-Cre, KLPVM-Cre, and TAT-Cre (5 and 10 μM) for 3 h in serum-free
(b-d) or 10% serum-containing
(e) media. After the incubation of cells with CPP-Cre fusion proteins, medium was changed to 10% FCS containing DMEM and cells were culture for additional 45 hrs. The delivery of Cre into Tex-lox P-EG cells was evaluated by GFP expression as previously reported [
11]. GFP expression was analyzed 48 h after the incubation of cells with Cre proteins.
b and c. FACS analysis of GFP expression in cells incubated with 5 μM
(b) or 10 μM
(c) Cre and CPP-Cre fusion proteins in serum-free medium. Representative data from three independent experiments are shown.
d. Mean GFP fluorescence intensity of Tex-loxP-EG cells incubated with 5 or 10 μM Cre recombinant proteins under serum-free conditions. Each bar represents three independent experiments and standard error.
e. Tex-loxP-EG cells incubated with purified recombinant Cre, VPTLK-Cre, KLPVM-Cre, and TAT-Cre (5 μM) in 10% serum-containing medium for 3 hrs. Images were captured 48 h after the incubation to detect GFP expression (right panel).
![Pharmaceuticals 03 03594 g005]()
We speculate that Fam helps VPTLK to integrate into the plasma membrane at high concentrations because Fam has high affinity for the plasma membrane, as evidenced by its application to visualize histological structure of the retina [
36]. The mechanism of cell entry of CPP5s is still unknown. The present study showed that almost all of the tested inhibitors of pinocytosis and endocytosis were unable to block the cell entry of Fam- or Tamra-labeled CPP5s (
Figure 2 and
Figure 3). To be noted, cytochalasin D was able to attenuate the cell-penetrating activity of Tamra-labeled CPP5s as well as Tamra-labeled R8 (
Figure 2d), though other inhibitors did not. The variation in the effects of inhibitors on the cell-entry activity between Fam- and Tamra-labeled peptides suggests that the fluorescent tag itself may have an influence on the cell-entry activity. Our attempts to determine the mechanism of cell entry by examining the effects of inhibitors did not give a clear answer. The present study, at least, suggests that a cytochalasin D-sensitive endocytic pathway may be involved in the cell-penetrating mechanism of CPP5s. However, since all other inhibitors of endocytosis and pinocytosis were not able to inhibit the cell entry of CPP5, it is possible that mechanisms other than endocytosis and pinocytosis play a major role in the cell-penetrating activity of CPP5s.
As listed in
Figure 1a, there are numerous penta-peptides that show cell-penetrating activity when the activity is measured by the cellular intake of Fam-labeled peptides. These peptides share similar composition, i.e. they contain one or two charged hydrophilic amino acids at the N- or C-terminus, and two to three hydrophobic amino acids additionally. This composition may allow CPP5s to possess affinity for the phospholipid membrane and/or a putative receptor(s) expressed in the membrane. Further studies of the mechanism of binding of CPP5s to the plasma membrane will provide clues to elucidate the mechanism of cell entry of CPP5s.
It is intriguing that TAT at higher concentrations induced apoptosis, which is the energy-dependent cellular response involving activation of specific proteases (Caspases) to induce programmed cell death6. Excess cationic peptides at the cell surface may trigger an apoptotic signal through an unknown mechanism. Identification of the apoptotic signal may help in the development of technologies to reduce the cytotoxicity of TAT. In our previous study, the fusion peptide of TAT and BIP (Bax-Inhibiting Peptide designed from Ku70) reduced the cytotoxicity of TAT in DAMI (a human megakaryocytic cancer cell line) cells6. Since BIP attenuated the toxicity of TAT, Bax may be involved in the TAT-induced apoptosis pathway. The combination of TAT and BIP may be one solution to attenuate the toxic side effects of TAT-dependent drug delivery.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





