
Immunomodulatory Expression of Cathelicidins Peptides in Pulp Inflammation and Regeneration: An Update

 , , , , , and
, , , , , and 
Abstract
:1. Introduction
2. Brief Literature Search
3. Methods and Materials
4. Results
5. Discussion
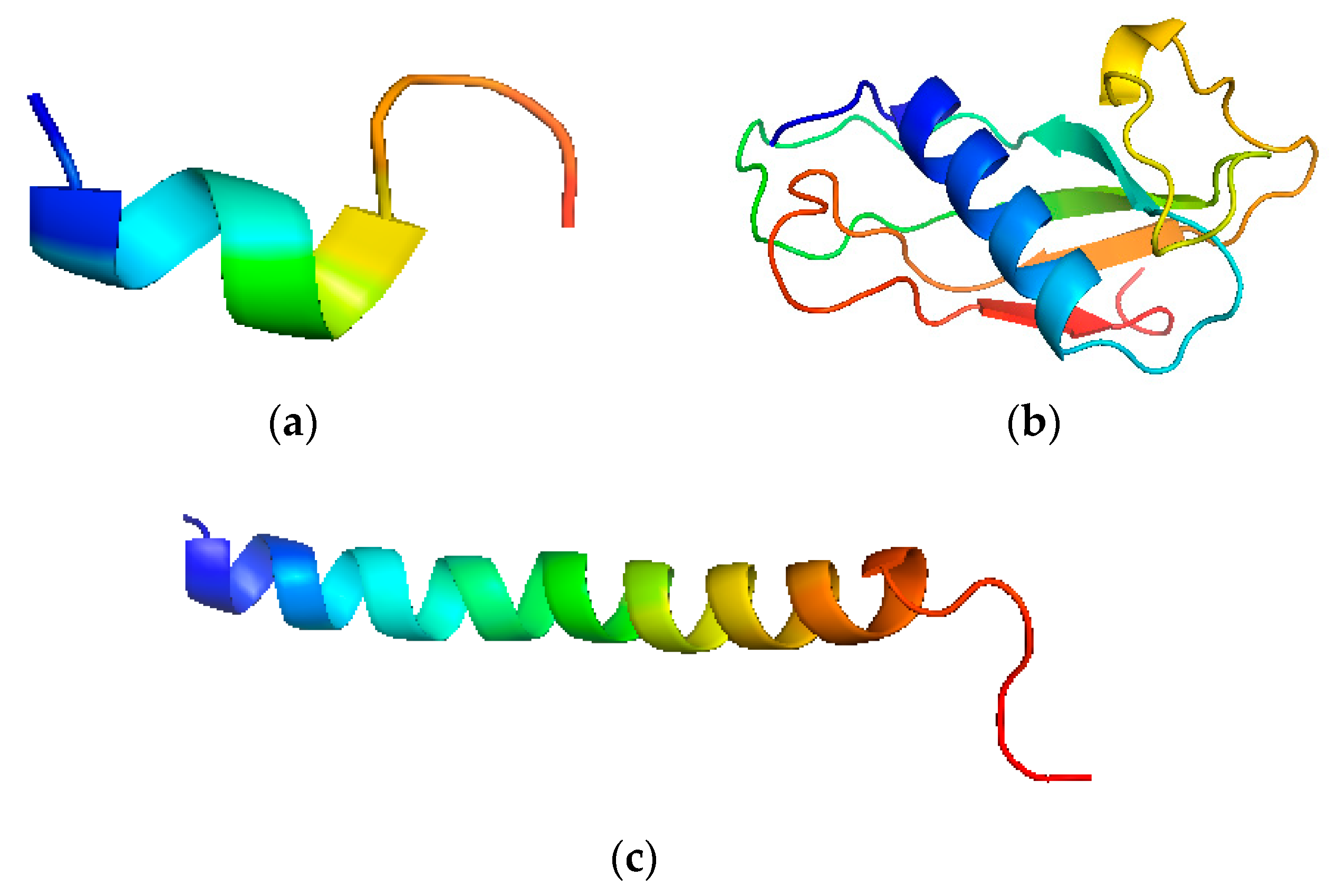
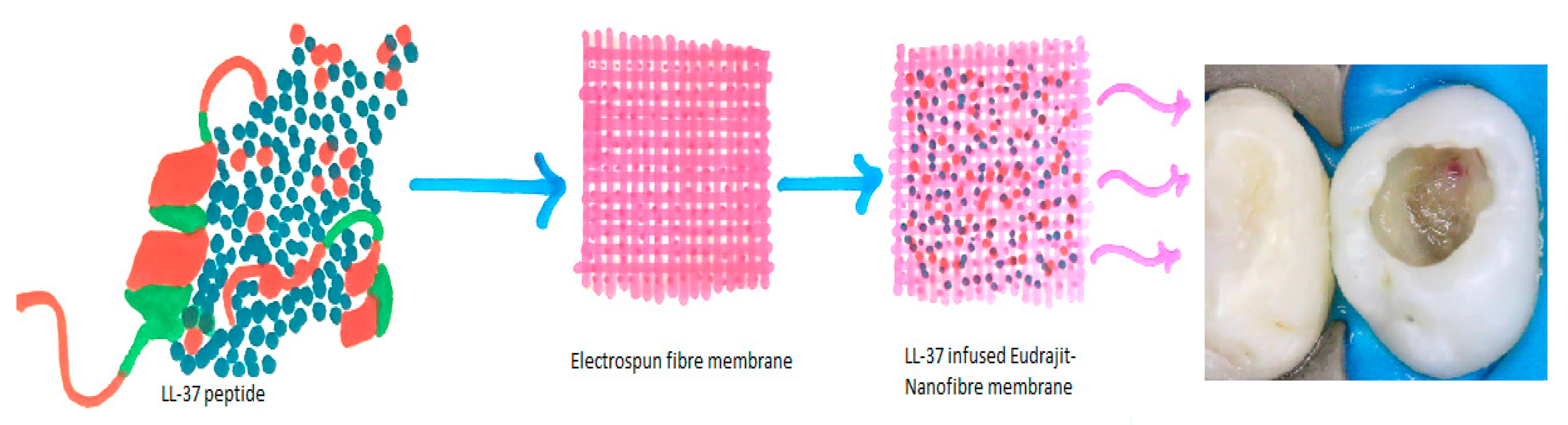
5.1. Structure and Function of LL-37
5.2. Antimicrobial Peptide Cathelicidins and Caries
5.3. Dental Pulp, Innate Immunity and Cathelicidins
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schmalz, G.; Smith, A.J. Pulp development, repair, and regeneration: Challenges of the transition from traditional dentistry to biologically based therapies. J. Endod. 2014, 40, S2–S5. [Google Scholar] [CrossRef]
- Shabbir, J.; Farooq, I.; Ali, S.; Mohammed, F.; Bugshan, A.; Khurram, S.A.; Khan, E. Dental Pulp. Illus. Guide Oral Histol. 2021, 69–79. [Google Scholar] [CrossRef]
- Park, S.H.; Ye, L.; Love, R.M.; Farges, J.C.; Yumoto, H. Inflammation of the Dental Pulp. Mediat. Inflamm. 2015, 2015, 980196. [Google Scholar] [CrossRef]
- Cooper, P.R.; Smith, A.J. Molecular mediators of pulp inflammation and regeneration. Endod. Top. 2013, 28, 90–105. [Google Scholar] [CrossRef]
- Zehra, T.; Qazi, F.; Abidi, Y.A.; Ahmed, S.; Khalili, S.; Saifee, J. Agreement between Two Commonly Used Pulp Tests in Determining Pulp Vitality. Pakistan J. Med. Dent. 2020, 9, 26–31. [Google Scholar] [CrossRef]
- Shabbir, J.; Khurshid, Z.; Qazi, F.; Sarwar, H.; Afaq, H.; Salman, S.; Adanir, N. Effect of Different Host-Related Factors on Postoperative Endodontic Pain in Necrotic Teeth Dressed with Interappointment Intracanal Medicaments: A Multicomparison Study. Eur. J. Dent. 2021, 15, 152–157. [Google Scholar] [PubMed]
- Shabbir, J.; Qazi, F.; Farooqui, W.; Ahmed, S.; Zehra, T.; Khurshid, Z. Effect of Chinese Propolis as an Intracanal Medicament on Post-Operative Endodontic Pain: A Double-Blind Randomized Controlled Trial. Int. J. Environ. Res. Public Health 2020, 17, 445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paris, S.; Wolgin, M.; Kielbassa, A.M.; Pries, A.; Zakrzewicz, A. Gene Expression of Human Beta-defensins in Healthy and Inflamed Human Dental Pulps. J. Endod. 2009, 35, 520–523. [Google Scholar] [CrossRef]
- Khurshid, Z.; Warsi, I.; Moin, S.F.; Slowey, P.D.; Latif, M.; Zohaib, S.; Zafar, M.S. Biochemical analysis of oral fluids for disease detection. In Advances in Clinical Chemistry; Academic Press: Cambridge, MA, USA, 2021; Volume 100, pp. 205–253. ISBN 9780128239223. [Google Scholar]
- Khurshid, Z.; Moin, S.F.; Khan, R.S.; Agwan, M.A.S.; Alwadaani, A.H.; Zafar, M.S. Human salivary protein extraction from RNAPro·SALTM, Pure·SALTM, and passive drooling method. Eur. J. Dent. 2017, 11, 385–389. [Google Scholar] [PubMed]
- Fatima, T.; Khurshid, Z.; Rehman, A.; Imran, E.; Srivastava, K.C.; Shrivastava, D. Gingival Crevicular Fluid (GCF): A Diagnostic Tool for the Detection of Periodontal Health and Diseases. Molecules 2021, 26, 1208. [Google Scholar]
- Hancock, R.E.W.; Sahl, H.G. Antimicrobial and host-defense peptides as new anti-infective therapeutic strategies. Nat. Biotechnol. 2006, 24, 1551–1557. [Google Scholar] [CrossRef] [PubMed]
- Khurshid, Z.; Naseem, M.; Asiri, F.Y.I.; Mali, M.; Khan, R.S.; Sahibzada, H.A.; Zafar, M.S.; Moin, S.F.; Khan, E. Significance and diagnostic role of antimicrobial cathelicidins (LL-37) peptides in oral health. Biomolecules 2017, 7, 80. [Google Scholar] [CrossRef] [Green Version]
- Khurshid, Z.; Naseem, M.; Sheikh, Z.; Najeeb, S.; Shahab, S.; Zafar, M.S. Oral antimicrobial peptides: Types and role in the oral cavity. Saudi Pharm. J. 2016, 24, 515–524. [Google Scholar] [CrossRef] [Green Version]
- Hancock, R.E.W.; Lehrer, R. Cationic peptides: A new source of antibiotics. Trends Biotechnol. 1998, 16, 82–88. [Google Scholar] [CrossRef]
- Liu, P.T.; Stenger, S.; Li, H.; Wenzel, L.; Tan, B.H.; Krutzik, S.R.; Ochoa, M.T.; Schauber, J.; Wu, K.; Meinken, C.; et al. Toll-like receptor triggering of a vitamin D-mediated human antimicrobial response. Science 2006, 311, 1770–1773. [Google Scholar] [CrossRef]
- Bals, R.; Weiner, D.J.; Moscioni, A.D.; Meegalla, R.L.; Wilson, J.M. Augmentation of innate host defense by expression of a cathelicidin antimicrobial peptide. Infect. Immun. 1999, 67, 6084–6089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarmiento, B.F.; Aminoshariae, A.; Bakkar, M.; Bonfield, T.; Ghosh, S.; Montagnese, T.A.; Mickel, A.K. The Expression of the Human Cathelicidin LL-37 in the Human Dental Pulp: An In vivo Study. Int. J. Pharm. 2016, 1, 5. [Google Scholar]
- Mojsoska, B.; Jenssen, H. Peptides and Peptidomimetics for Antimicrobial Drug Design. Pharmaceuticals 2015, 8, 366–415. [Google Scholar] [CrossRef]
- Winfred, S.B.; Meiyazagan, G.; Panda, J.J.; Nagendrababu, V.; Deivanayagam, K.; Chauhan, V.S.; Venkatraman, G. Antimicrobial activity of cationic peptides in endodontic procedures. Eur. J. Dent. 2014, 8, 254–260. [Google Scholar] [CrossRef]
- Davidopoulou, S.; Diza, E.; Menexes, G.; Kalfas, S. Salivary concentration of the antimicrobial peptide LL-37 in children. Arch. Oral Biol. 2012, 57, 865–869. [Google Scholar] [CrossRef]
- Ouhara, K.; Komatsuzawa, H.; Yamada, S.; Shiba, H.; Fujiwara, T.; Ohara, M.; Sayama, K.; Hashimoto, K.; Kurihara, H.; Sugai, M. Susceptibilities of periodontopathogenic and cariogenic bacteria to antibacterial peptides, β-defensins and LL37, produced by human epithelial cells. J. Antimicrob. Chemother. 2005, 55, 888–896. [Google Scholar] [CrossRef] [PubMed]
- Tao, R.; Jurevic, R.J.; Coulton, K.K.; Tsutsui, M.T.; Roberts, M.C.; Kimball, J.R.; Wells, N.; Berndt, J.; Dale, B.A. Salivary antimicrobial peptide expression and dental caries experience in children. Antimicrob. Agents Chemother. 2005, 49, 3883–3888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Q.; Zeng, K.; Kang, Q.; Qian, W.; Zhang, W.; Gan, Q.; Xia, W. The Antimicrobial Peptide LL-37 Promotes Migration and Odonto/Osteogenic Differentiation of Stem Cells from the Apical Papilla through the Akt/Wnt/β-catenin Signaling Pathway. J. Endod. 2020, 46, 964–972. [Google Scholar] [CrossRef] [PubMed]
- Khung, R.; Shiba, H.; Kajiya, M.; Kittaka, M.; Ouhara, K.; Takeda, K.; Mizuno, N.; Fujita, T.; Komatsuzawa, H.; Kurihara, H. LL37 induces VEGF expression in dental pulp cells through ERK signalling. Int. Endod. J. 2015, 48, 673–679. [Google Scholar] [CrossRef]
- Rechenberg, D.K.; Galicia, J.C.; Peters, O.A. Biological markers for pulpal inflammation: A systematic review. PLoS ONE 2016, 11, e0167289. [Google Scholar]
- Gennaro, R.; Skerlavaj, B.; Romeo, D. Purification, composition, and activity of two bactenecins, antibacterial peptides of bovine neutrophils. Infect. Immun. 1989, 57, 3142–3146. [Google Scholar] [CrossRef] [Green Version]
- Shaykhiev, R.; Beißwenger, C.; Kändler, K.; Senske, J.; Püchner, A.; Damm, T.; Behr, J.; Bals, R. Human endogenous antibiotic LL-37 stimulates airway epithelial cell proliferation and wound closure. Am. J. Physiol. Lung Cell. Mol. Physiol. 2005, 289, L842–L848. [Google Scholar] [CrossRef]
- Dürr, U.H.N.; Sudheendra, U.S.; Ramamoorthy, A. LL-37, the only human member of the cathelicidin family of antimicrobial peptides. Biochim. Biophys. Acta Biomembr. 2006, 1758, 1408–1425. [Google Scholar] [CrossRef] [Green Version]
- Nagaoka, I.; Hirata, M.; Sugimoto, K.; Tsutsumi-Ishii, Y.; Someya, A.; Saionji, K.; Igari, J. Evaluation of the expression of human CAP18 gene during neutrophil maturation in the bone marrow. J. Leukoc. Biol. 1998, 64, 845–852. [Google Scholar] [CrossRef]
- Seil, M.; Nagant, C.; Dehaye, J.-P.; Vandenbranden, M.; Lensink, M.F. Spotlight on Human LL-37, an Immunomodulatory Peptide with Promising Cell-Penetrating Properties. Pharmaceuticals 2010, 3, 3435–3460. [Google Scholar] [CrossRef] [Green Version]
- Vandamme, D.; Landuyt, B.; Luyten, W.; Schoofs, L. A Comprehensive Summary of LL-37, the Factotum Human Cathelicidin Peptide. Cell. Immunol. 2012, 280, 22–35. [Google Scholar] [CrossRef]
- Duplantier, A.J.; van Hoek, M.L. The Human Cathelicidin Antimicrobial Peptide LL-37 as a Potential Treatment for Polymicrobial Infected Wounds. Front. Immunol. 2013, 4, 143. [Google Scholar] [CrossRef] [Green Version]
- Larrick, J.W.; Morgan, J.G.; Palings, I.; Hirata, M.; Yen, M.H. Complementary D.N.A. sequence of rabbit CAP18-A unique lipopolysaccharide binding protein. Biochem. Biophys. Res. Commun. 1991, 179, 170–175. [Google Scholar] [CrossRef]
- Clarke, J.K. On the Bacterial Factor in the Aetiology of Dental Caries. Br. J. Exp. Pathol. 1924, 5, 141–147. [Google Scholar]
- Hicks, J.; Garcia-Godoy, F.; Flaitz, C. Biological factors in dental caries: Role of saliva and dental plaque in the dynamic process of demineralization and remineralization (part 1). J. Clin. Pediatr. Dent. 2003, 28, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Vitorino, R.; Lobo, M.J.C.; Duarte, J.R.; Ferrer-Correia, A.J.; Domingues, P.M.; Amado, F.M.L. The role of salivary peptides in dental caries. Biomed. Chromatogr. 2005, 19, 214–222. [Google Scholar] [CrossRef] [PubMed]
- Mizukawa, N.; Sugiyama, K.; Mishima, T.U.; Takagi, S.; Sugahara, T. Levels of human defensin-1, an antimicrobial peptide, in saliva of patients with oral inflammation. Oral Surg. Oral Med. Oral Pathol. Oral Radiol. Endod. 1999, 87, 539–543. [Google Scholar] [CrossRef]
- Hancock, R.E.W.; Diamond, G. The role of cationic antimicrobial peptides in innate host defences. Trends Microbiol. 2000, 8, 402–410. [Google Scholar] [CrossRef]
- Bonass, W.A.; High, A.S.; Owen, P.J.; Devine, D.A. Expression of β-defensin genes by human salivary glands. Oral Microbiol. Immunol. 1999, 14, 371–374. [Google Scholar] [CrossRef]
- Staquet, M.J.; Carrouel, F.; Keller, J.F.; Baudouin, C.; Msika, P.; Bleicher, F.; Kufer, T.A.; Farges, J.C. Pattern-recognition receptors in pulp defense. Adv. Dent. Res. 2011, 23, 296–301. [Google Scholar] [CrossRef]
- Selders, G.S.; Fetz, A.E.; Radic, M.Z.; Bowlin, G.L. An overview of the role of neutrophils in innate immunity, inflammation and host-biomaterial integration. Regen. Biomater. 2017, 4, 55–68. [Google Scholar] [CrossRef] [PubMed]
- Yumoto, H.; Hirao, K.; Hosokawa, Y.; Kuramoto, H.; Takegawa, D.; Nakanishi, T.; Matsuo, T. The roles of odontoblasts in dental pulp innate immunity. Jpn. Dent. Sci. Rev. 2018, 54, 105–117. [Google Scholar] [CrossRef] [PubMed]
- Durand, S.H.; Flacher, V.; Roméas, A.; Carrouel, F.; Colomb, E.; Vincent, C.; Magloire, H.; Couble, M.-L.; Bleicher, F.; Staquet, M.-J.; et al. Lipoteichoic Acid Increases T.L.R. and Functional Chemokine Expression while Reducing Dentin Formation in In Vitro Differentiated Human Odontoblasts. J. Immunol. 2006, 176, 2880–2887. [Google Scholar] [CrossRef] [PubMed]
- Palosaari, H.; Wahlgren, J.; Larrnas, M.; Rönkä, H.; Sorsa, T.; Sato, T.; Tjäderhane, L. The expression of mmp-8 in human odontoblasts and dental pulp cells is down-regulated by tgf-β 1. J. Dent. Res. 2000, 79, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Arana-Chavez, V.E.; Massa, L.F. Odontoblasts: The cells forming and maintaining dentine. Int. J. Biochem. Cell Biol. 2004, 36, 1367–1373. [Google Scholar] [CrossRef]
- Cekici, A.; Kantarci, A.; Hasturk, H.; Van Dyke, T.E. Inflammatory and immune pathways in the pathogenesis of periodontal disease. Periodontol. 2000 2014, 64, 57–80. [Google Scholar] [CrossRef] [Green Version]
- Mogensen, T.H. Pathogen recognition and inflammatory signaling in innate immune defenses. Clin. Microbiol. Rev. 2009, 22, 240–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horst, O.V.; Tompkins, K.A.; Coats, S.R.; Braham, P.H.; Darveau, R.P.; Dale, B.A. TGF-β1 inhibits TLR-mediated odontoblast responses to oral bacteria. J. Dent. Res. 2009, 88, 333–338. [Google Scholar] [CrossRef] [Green Version]
- Veerayutthwilai, O.; Byers, M.R.; Pham, T.T.T.; Darveau, R.P.; Dale, B.A. Differential regulation of immune responses by odontoblasts. Oral Microbiol. Immunol. 2007, 22, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Ahlquist, M.L.; Franzen, O.G. Inflammation and dental pain in man. Dent. Traumatol. 1994, 10, 201–209. [Google Scholar] [CrossRef]
- Bergenholtz, G. Inflammatory response of the dental pulp to bacterial irritation. J. Endod. 1981, 7, 100–104. [Google Scholar] [CrossRef]
- Kajiya, M.; Shiba, H.; Komatsuzawa, H.; Ouhara, K.; Fujita, T.; Takeda, K.; Uchida, Y.; Mizuno, N.; Kawaguchi, H.; Kurihara, H. The antimicrobial peptide LL37 induces the migration of human pulp cells: A possible adjunct for regenerative endodontics. J. Endod. 2010, 36, 1009–1013. [Google Scholar] [CrossRef]
- Odlén, K.; Fält, F.; Dahl, S.; Aidoukovitch, A.; Ericson, D.; Nilsson, B.O.; Hedenbjörk-Lager, A. Odontoblast-like MDPC-23 cells produce proinflammatory IL-6 in response to lipoteichoic acid and express the antimicrobial peptide CRAMP. Acta Odontol. Scand. 2020, 78, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Borregaard, N.; Cowland, J.B. Granules of the human neutrophilic polymorphonuclear leukocyte. Blood 1997, 89, 3503–3521. [Google Scholar] [CrossRef]
- Bülow, E.; Bengtsson, N.; Calafat, J.; Gullberg, U.; Olsson, I. Sorting of neutrophil-specific granule protein human cathelicidin, hCAP-18, when constitutively expressed in myeloid cells. J. Leukoc. Biol. 2002, 72, 147–153. [Google Scholar] [PubMed]
- Koczulla, R.; Von Degenfeld, G.; Kupatt, C.; Krötz, F.; Zahler, S.; Gloe, T.; Issbrücker, K.; Unterberger, P.; Zaiou, M.; Lebherz, C.; et al. An angiogenic role for the human peptide antibiotic LL-37/hCAP-18. J. Clin. Investig. 2003, 111, 1665–1672. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, K.; Suzuki, S.; Kawada-Matsuo, M.; Nakanishi, J.; Hirata-Tsuchiya, S.; Komatsuzawa, H.; Yamada, S.; Shiba, H. Heparin–LL37 complexes are less cytotoxic for human dental pulp cells and have undiminished antimicrobial and LPS-neutralizing abilities. Int. Endod. J. 2019, 52, 1327–1343. [Google Scholar] [CrossRef]
- Chang, W.K.; Wimley, W.C.; Searson, P.C.; Hristova, K.; Merzlyakov, M. Characterization of antimicrobial peptide activity by electrochemical impedance spectroscopy. Biochim. Biophys. Acta Biomembr. 2008, 1778, 2430–2436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pütsep, K.; Carlsson, G.; Boman, H.G.; Andersson, M. Deficiency of antibacterial peptides in patients with morbus Kostmann: An observation study. Lancet 2002, 360, 1144–1149. [Google Scholar] [CrossRef]
- Esfandiyari, R.; Halabian, R.; Behzadi, E.; Sedighian, H.; Jafari, R.; Imani Fooladi, A.A. Performance evaluation of antimicrobial peptide ll-37 and hepcidin and β-defensin-2 secreted by mesenchymal stem cells. Heliyon 2019, 5, e02652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Chen, L.; Chen, Z.; Wu, L.; Feng, J.; Wang, F.; Shoff, L.; Li, X.; Donly, K.J.; MacDougall, M.; et al. Dentin sialoprotein facilitates dental mesenchymal cell differentiation and dentin formation. Sci. Rep. 2017, 7, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Pober, J.S. Cytokine-mediated activation of vascular endothelium. Physiology and pathology. Am. J. Pathol. 1988, 133, 426–433. [Google Scholar] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Author | Experimental Area | Methodology | Outcome | Conclusion | Remarks |
|---|---|---|---|---|---|
| Sarmiento et al., 2016 [18] | To identify LL-37 levels in normal and inflamed pulp | 10 diagnosed symptomatic irreversible pulpitis with apical periodontitis and 10 Healthy pulp were chosen. Estimation of LL-37 was done by ELISA method. | Mean pulpal concentration of LL-37 in symptomatic irreversible pulp is 1.5 ± 1.2 ng/mL and in normal pulp is 0.2 ± 0.6 ng/mL. | Inflammation of dental pulp induces expression of LL-37 ad therefore LL-37 plays a major role in innate immunity. | Small sample size selection might be a bias to standardize the levels of LL-37 in inflamed and normal pulp. The degree of inflammation of pulp required to stimulate LL-37 expression is another factor that needs to be assessed. |
| Tao et al., 2005 [23] | To identify possible relationship between Antimicrobial peptide levels in saliva and caries experience in middle school children | Saliva of 144 children under the age group of 11–15 yrs were analyzed for LL-37 and hBD3, HNP1-3. | The mean of LL-37 showed extensive variation between individuals. The level of (α-defensins) HNP1-3 were inversely correlated with caries. | Children with caries have significantly lower levels of α-defensins and low salivary levels of H.N.P 1-3 was inversely correlated to caries | The study showed alpha-defensins can be potential markers for caries risk. Also mentioned that synthetic peptides can be developed to enhance peptide expression and can be therapeutic agent. |
| Ouhara et al., 2005 [22] | Anti microbial peptide hBD1, hBD2, hBD3 and LL-37 were evaluated for their antimicrobial activity against oral bacteria | Synthetic peptides were treated with various oral bacteria Aggregatibacter actinomycetemcomitans, Porphyromonas gingivalis, Prevotella intermedia, Fusobacterium nucleatum, Streptococcus mutans, Streptococcus sobrinus, Streptococcus salivarius, Streptococcus sanguis, Streptococcus mitis and Lactobacillus casei | Antibacterial action of hBD1 was lower than other peptides F. nucleatum was highly susceptible to hBD 3 and LL-37. S. mutans was highly susceptible to hBD3 | β-defensins and LL-37 have potential antibacterial action against oral bacteria | The net charge on the bacterial surface is another factor that can be considered, also selective inhibition by these peptides can be a contradictory factor that can be evaluated through invivo studies. |
| Cheng et al., 2020 [24] | Evaluate the effect of LL-37 on proliferation, migration and differentiation of Stem cells from apical papilla (SCAP) | SCAPs were isolated from third molars of age group 16–20 Y, cultured, characterized. The cell viability was analyzed with cell-commet assay kit. Dentin Sialophosphoprotein, dentin matrix protein 1 were assessed by quantitative polymerase chain reaction and western blot. | 2.5 µg/mL of LL-37 regulated odonto/osteogenic markers. LL-37 promoted alkaline phosphatase activity | LL-37 at 2.5 µg/mL promoted the migration and odonto/osteo differential SCAPs by activating the AKt/Wnt/β-catenin signalling | In vivo assay could have been a better option, to understand therapeutic action of LL-37 on apical lesions. |
| Khung et al., 2015 [25] | To examine in-vitro effects of LL-37 on expression of vascular endothelial growth factors in human pulp cells | Pulp cells at passage 6 were treated with 10 µg/mL−1 synthesized LL-37 and inhibition assay was performed with MAPk or NF-kB inhibits VEGF, mRNA, VEGF protein andphophorylated ERK were determined byRT PCk, ELISA and western blot | LL-37 significantly increased both mRNA and protein levels VEGF in pulp cell. | LL-37 activated the ERK pathway to boost VEGF secretion from human pulp cells. These for can be a potent pulp capping agent. | This article clearly suggests the role of LL-37 in pulpal cell migration and thus a potent pulp capping agent. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nireeksha; Varma, S.R.; Damdoum, M.; Alsaegh, M.A.; Hegde, M.N.; Kumari, S.N.; Ramamurthy, S.; Narayanan, J.; Imran, E.; Shabbir, J.; et al. Immunomodulatory Expression of Cathelicidins Peptides in Pulp Inflammation and Regeneration: An Update. Curr. Issues Mol. Biol. 2021, 43, 116-126. https://doi.org/10.3390/cimb43010010
Nireeksha, Varma SR, Damdoum M, Alsaegh MA, Hegde MN, Kumari SN, Ramamurthy S, Narayanan J, Imran E, Shabbir J, et al. Immunomodulatory Expression of Cathelicidins Peptides in Pulp Inflammation and Regeneration: An Update. Current Issues in Molecular Biology. 2021; 43(1):116-126. https://doi.org/10.3390/cimb43010010
Chicago/Turabian StyleNireeksha, Sudhir Rama Varma, Marah Damdoum, Mohammed Amjed Alsaegh, Mithra N. Hegde, Suchetha N. Kumari, Srinivasan Ramamurthy, Jayaraj Narayanan, Eisha Imran, Juzer Shabbir, and et al. 2021. "Immunomodulatory Expression of Cathelicidins Peptides in Pulp Inflammation and Regeneration: An Update" Current Issues in Molecular Biology 43, no. 1: 116-126. https://doi.org/10.3390/cimb43010010
APA StyleNireeksha, Varma, S. R., Damdoum, M., Alsaegh, M. A., Hegde, M. N., Kumari, S. N., Ramamurthy, S., Narayanan, J., Imran, E., Shabbir, J., & Khurshid, Z. (2021). Immunomodulatory Expression of Cathelicidins Peptides in Pulp Inflammation and Regeneration: An Update. Current Issues in Molecular Biology, 43(1), 116-126. https://doi.org/10.3390/cimb43010010