Differentially Expressed Extracellular Vesicle-Contained microRNAs before and after Transurethral Resection of Bladder Tumors

 and
and
Abstract
:1. Introduction
2. Results
2.1. Patient Characteristics
2.2. Biosource Separation Based on Principal Component Analysis
2.3. EV Characterization
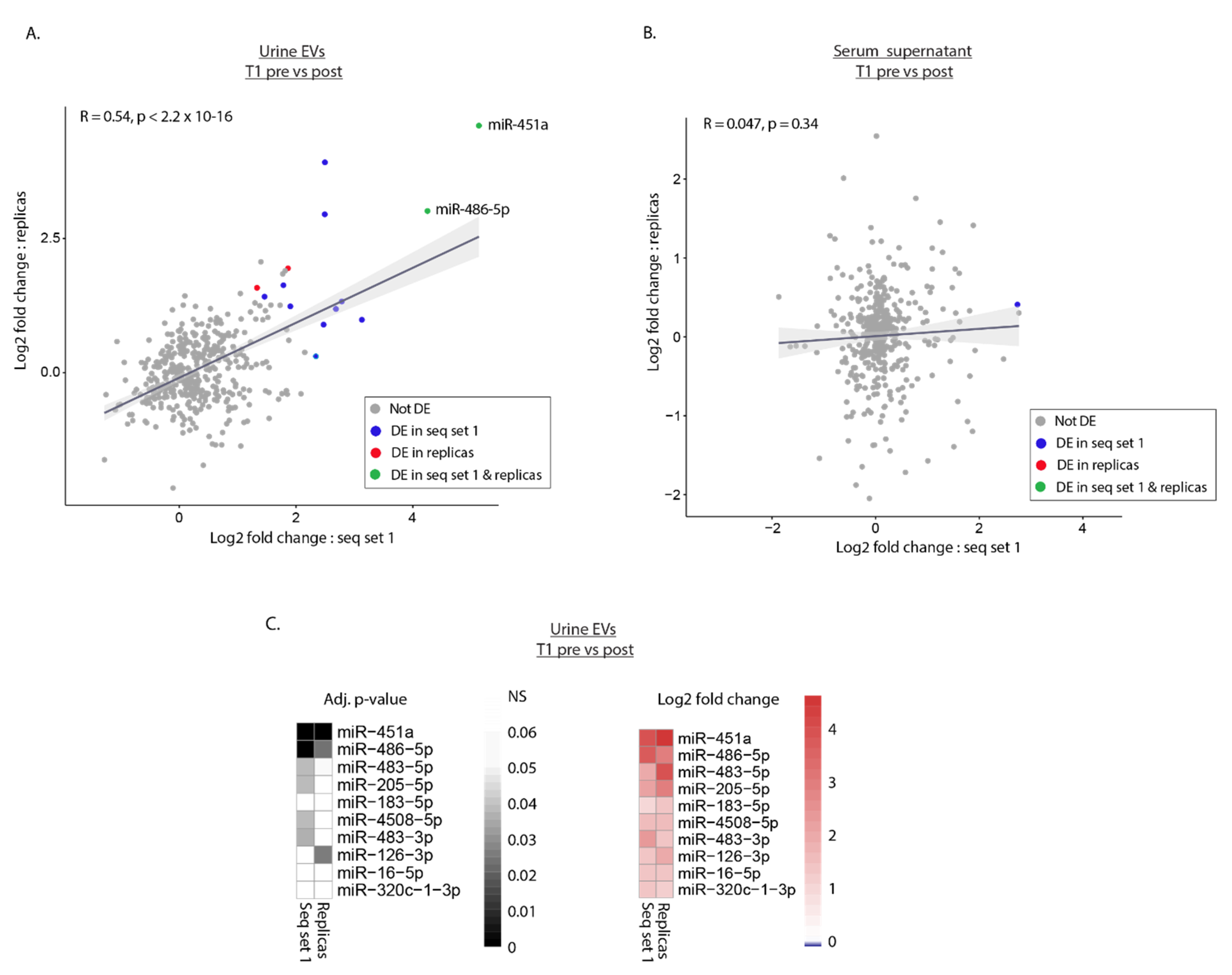
2.4. Differential Expression Analysis Reveals Potential MiRNA Biomarkers for T1 BC Patients
2.5. Replica Sequencing Confirms Differential Expression of miR-451a and miR-486-5p in Urinary EVs
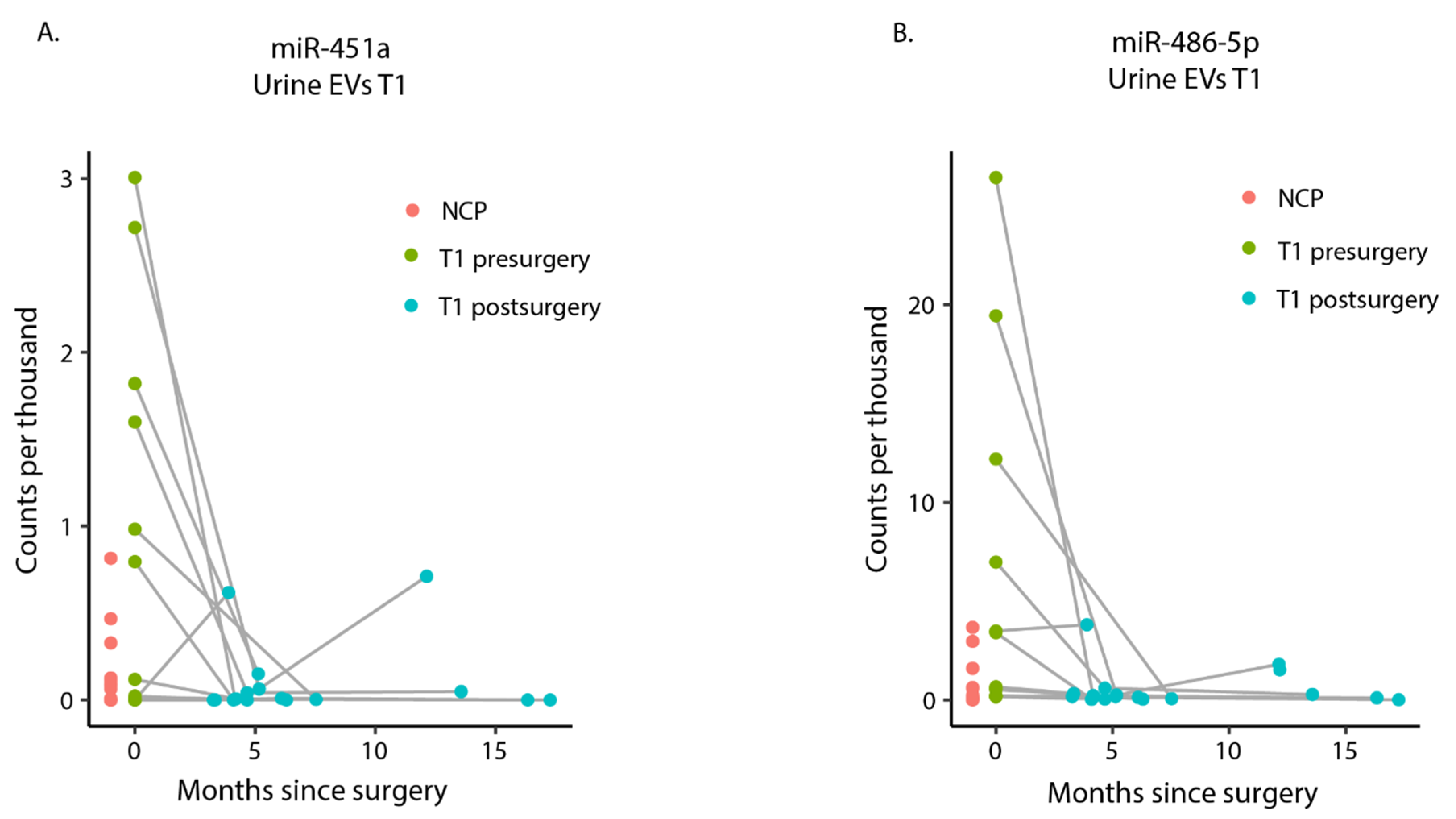
2.6. miR-451a and miR-486-5p Are Highly Expressed in Presurgery Urinary EVs among T1 Patients
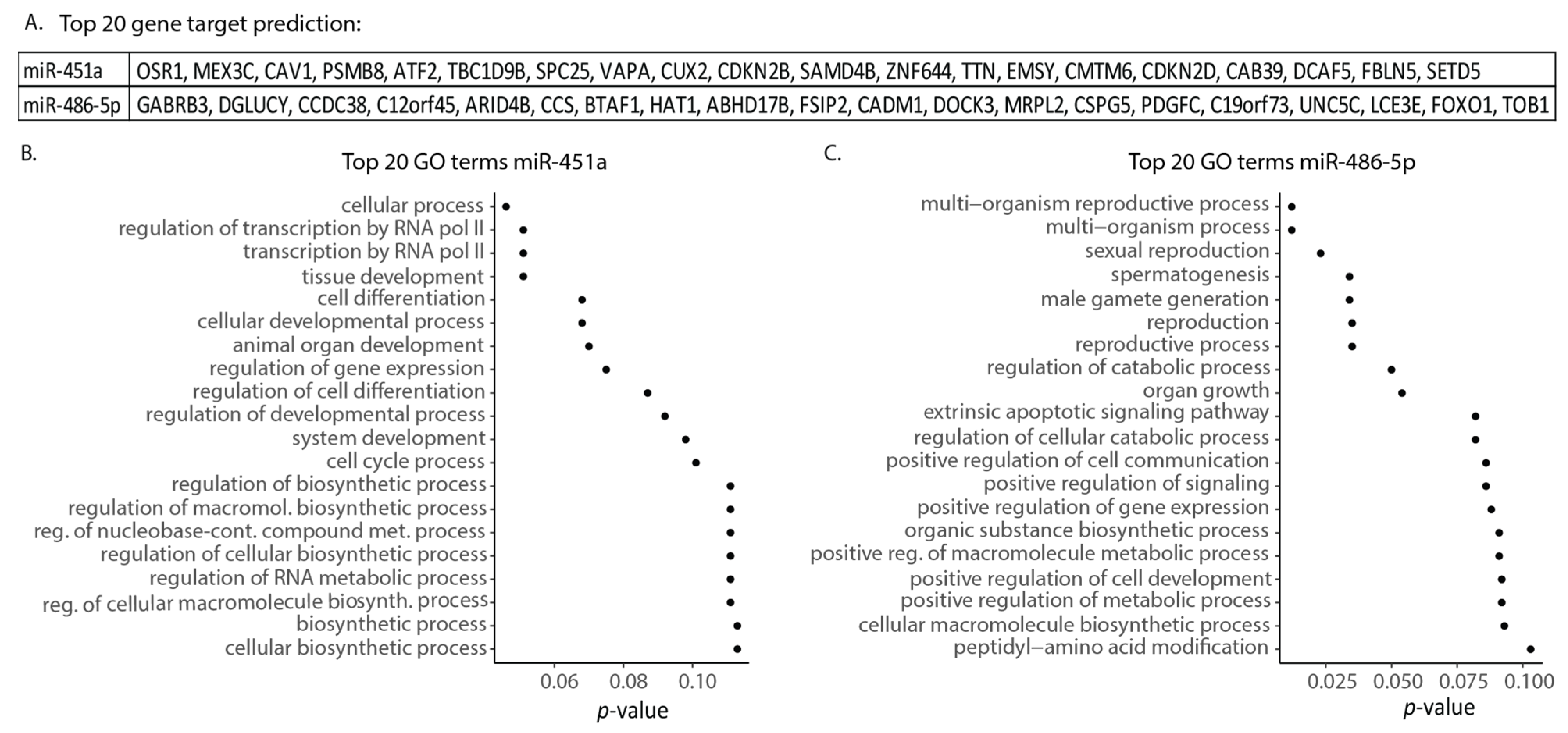
2.7. miR-451a and miR-486-5p Are Involved in Transcription Regulation, Cell Differentiation, and Apoptotic Signaling
3. Discussion
4. Materials and Methods
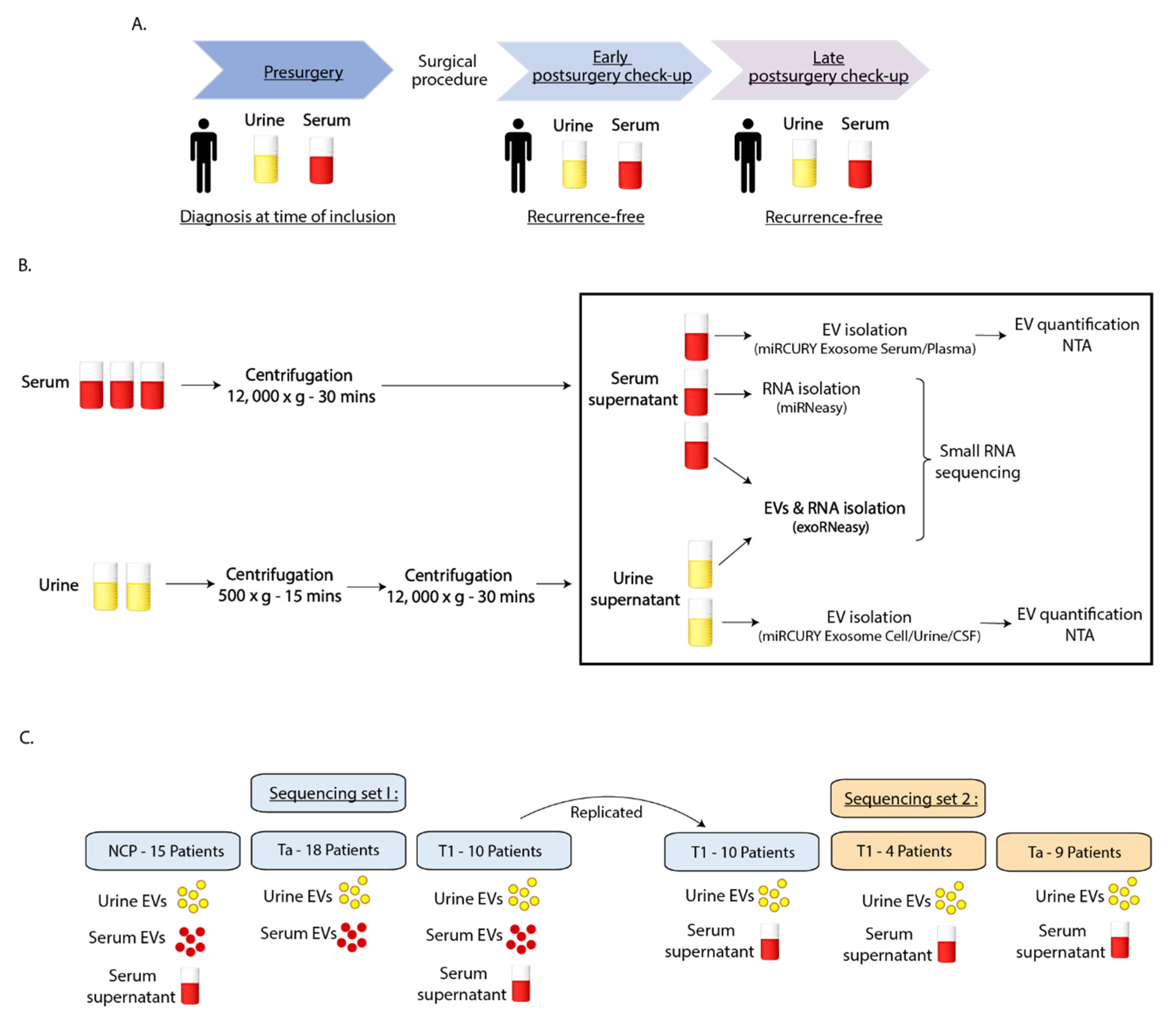
4.1. Clinical Samples
4.2. Biobanking Procedure
4.3. Sequencing Study Design
4.4. Isolation of EVs and Small Non-Coding RNAs
4.5. Nanoparticle Tracking Analysis
4.6. Small RNA Sequencing
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Turan, H.; Balci, U.; Erdinc, F.S.; Tülek, N.; Germiyanoglu, C. Bacteriuria, pyuria and bacteremia frequency following outpatient cystoscopy. Int. J. Urol. 2006, 13, 25–28. [Google Scholar] [CrossRef]
- Biardeau, X.; Lam, O.; Ba, V.; Campeau, L.; Corcos, J. Prospective evaluation of anxiety, pain, and embarrassment associated with cystoscopy and urodynamic testing in clinical practice. Can. Urol. Assoc. J. 2017, 11, 104. [Google Scholar] [CrossRef] [PubMed]
- Planz, B.; Jochims, E.; Deix, T.; Caspers, H.; Jakse, G.; Boecking, A. The role of urinary cytology for detection of bladder cancer. Eur. J. Surg. Oncol. (EJSO) 2005, 31, 304–308. [Google Scholar] [CrossRef] [PubMed]
- Kehinde, E.O.; Al-Mulla, F.; Kapila, K.; Anim, J.T. Comparison of the sensitivity and specificity of urine cytology, urinary nuclear matrix protein-22 and multitarget fluorescence in situ hybridization assay in the detection of bladder cancer. Scand. J. Urol. Nephrol. 2010, 45, 113–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McIntire, P.J.; Khan, R.; Hussain, H.; Pambuccian, S.E.; Wojcik, E.M.; Barkan, G.A. Negative predictive value and sensitivity of urine cytology prior to implementation of The Paris System for Reporting Urinary Cytology. Cancer Cytopathol. 2019, 127, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Kowal, J.; Tkach, M.; Théry, C. Biogenesis and secretion of exosomes. Curr. Opin. Cell Biol. 2014, 29, 116–125. [Google Scholar] [CrossRef] [Green Version]
- Colombo, M.; Raposo, G.; Théry, C. Biogenesis, secretion, and intercellular interactions of exosomes and other extracellular vesicles. Annu. Rev. Cell Dev. Biol. 2014, 30, 255–289. [Google Scholar] [CrossRef]
- Bucki, R.; Bachelot-Loza, C.; Zachowski, A.; Giraud, F.; Sulpice, J.-C. Calcium Induces Phospholipid Redistribution and Microvesicle Release in Human Erythrocyte Membranes by Independent Pathways. Biochemistry 1998, 37, 15383–15391. [Google Scholar] [CrossRef]
- Cocucci, E.; Racchetti, G.; Meldolesi, J. Shedding microvesicles: Artefacts no more. Trends Cell Biol. 2009, 19, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Turchinovich, A.; Drapkina, O.; Tonevitsky, A. Transcriptome of Extracellular Vesicles: State-of-the-Art. Front. Immunol. 2019, 10, 202. [Google Scholar] [CrossRef] [Green Version]
- Sork, H.; Corso, G.; Krjutskov, K.; Johansson, H.J.; Nordin, J.Z.; Wiklander, O.P.B.; Lee, Y.X.F.; Westholm, J.O.; Lehtiö, J.; Wood, M.J.A.; et al. Heterogeneity and interplay of the extracellular vesicle small RNA transcriptome and proteome. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Kim, K.M.; Abdelmohsen, K.; Mustapic, M.; Kapogiannis, D.; Gorospe, M. RNA in extracellular vesicles. Wiley Interdiscip. Rev. RNA 2017, 8, e1413. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Liao, K.; Zhou, W. Exosomes Regulate the Transformation of Cancer Cells in Cancer Stem Cell Homeostasis. Stem Cells Int. 2018, 2018, 1–16. [Google Scholar] [CrossRef]
- Tkach, M.; Théry, C. Communication by Extracellular Vesicles: Where We Are and Where We Need to Go. Cell 2016, 164, 1226–1232. [Google Scholar] [CrossRef] [Green Version]
- Boukouris, S.; Mathivanan, S. Exosomes in bodily fluids are a highly stable resource of disease biomarkers. Proteom. Clin. Appl. 2015, 9, 358–367. [Google Scholar] [CrossRef] [Green Version]
- Soung, Y.H.; Ford, S.; Zhang, V.; Chung, J. Exosomes in Cancer Diagnostics. Cancers 2017, 9, 8. [Google Scholar] [CrossRef] [Green Version]
- Oeyen, E.; Hoekx, L.; De Wachter, S.; Baldewijns, M.; Ameye, F.; Mertens, I. Bladder Cancer Diagnosis and Follow-Up: The Current Status and Possible Role of Extracellular Vesicles. Int. J. Mol. Sci. 2019, 20, 821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugawara, S.; Yamada, Y.; Arai, T.; Okato, A.; Idichi, T.; Kato, M.; Koshizuka, K.; Ichikawa, T.; Seki, N. Dual strands of the miR-223 duplex (miR-223-5p and miR-223-3p) inhibit cancer cell aggressiveness: Targeted genes are involved in bladder cancer pathogenesis. J. Hum. Genet. 2018, 63, 657–668. [Google Scholar] [CrossRef] [PubMed]
- Li, P.; Yang, X.; Yuan, W.; Yang, C.; Zhang, X.; Han, J.; Wang, J.; Deng, X.; Yang, H.; Li, P.; et al. CircRNA-Cdr1as Exerts Anti-Oncogenic Functions in Bladder Cancer by Sponging MicroRNA-135a. Cell Physiol. Biochem. 2018, 46, 1606–1616. [Google Scholar] [CrossRef]
- Valadi, H.; Ekstrom, K.; Bossios, A.; Sjostrand, M.; Lee, J.J.; Lotvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujita, Y.; Yoshioka, Y.; Ochiya, T. Extracellular vesicle transfer of cancer pathogenic components. Cancer Sci. 2016, 107, 385–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.; Shi, K.; Yang, S.; Liu, J.; Zhou, Q.; Wang, G.; Song, J.; Li, Z.; Zhang, Z.; Yuan, W. Effect of exosomal miRNA on cancer biology and clinical applications. Mol. Cancer 2018, 17, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Bhome, R.; Del Vecchio, F.; Lee, G.-H.; Bullock, M.D.; Primrose, J.N.; Sayan, A.E.; Mirnezami, A.H. Exosomal microRNAs (exomiRs): Small molecules with a big role in cancer. Cancer Lett. 2018, 420, 228–235. [Google Scholar] [CrossRef]
- Salehi, M.; Sharifi, M. Exosomal miRNAs as novel cancer biomarkers: Challenges and opportunities. J. Cell. Physiol. 2018, 233, 6370–6380. [Google Scholar] [CrossRef]
- Baumgart, S.; Hölters, S.; Ohlmann, C.-H.; Bohle, R.; Stöckle, M.; Ostenfeld, M.S.; Dyrskjøt, L.; Junker, K.; Heinzelmann, J. Exosomes of invasive urothelial carcinoma cells are characterized by a specific miRNA expression signature. Oncotarget 2017, 8, 58278–58291. [Google Scholar] [CrossRef] [Green Version]
- Andreu, Z.; Oshiro, R.O.; Redruello, A.; López-Martín, S.; Gutiérrez-Vázquez, C.; Morato, E.; Marina, A.I.; Gómez, C.O.; Yáñez-Mó, M. Extracellular vesicles as a source for non-invasive biomarkers in bladder cancer progression. Eur. J. Pharm. Sci. 2017, 98, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Matsuzaki, K.; Fujita, K.; Jingushi, K.; Kawashima, A.; Ujike, T.; Nagahara, A.; Ueda, Y.; Tanigawa, G.; Yoshioka, I.; Ueda, K.; et al. MiR-21-5p in urinary extracellular vesicles is a novel biomarker of urothelial carcinoma. Oncotarget 2017, 8, 24668–24678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.-R.; Ortiz-Bonilla, C.J.; Lee, Y.-F. Extracellular Vesicles in Bladder Cancer: Biomarkers and Beyond. Int. J. Mol. Sci. 2018, 19, 2822. [Google Scholar] [CrossRef] [Green Version]
- Piao, X.-M.; Cha, E.-J.; Yun, S.J.; Kim, W.-J. Role of Exosomal miRNA in Bladder Cancer: A Promising Liquid Biopsy Biomarker. Int. J. Mol. Sci. 2021, 22, 1713. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Miao, T.; Feng, T.; Jiang, Z.; Li, M.; Zhou, L.; Li, H. miR-451a Inhibited Cell Proliferation and Enhanced Tamoxifen Sensitive in Breast Cancer via Macrophage Migration Inhibitory Factor. BioMed. Res. Int. 2015, 2015, 207684. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.-Y.; Cui, J.-Y.; Yuan, J.; Wang, X. MiR-451a suppressed cell migration and invasion in non-small cell lung cancer through targeting ATF2. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 5554–5561. [Google Scholar] [PubMed]
- Minna, E.; Romeo, P.; Dugo, M.; De Cecco, L.; Todoerti, K.; Pilotti, S.; Perrone, F.; Seregni, E.; Agnelli, L.; Neri, A.; et al. miR-451a is underexpressed and targets AKT/mTOR pathway in papillary thyroid carcinoma. Oncotarget 2016, 7, 12731–12747. [Google Scholar] [CrossRef] [PubMed]
- Zeng, T.; Peng, L.; Chao, C.; Fu, B.; Wang, G.; Wang, Y.; Zhu, X. miR-451 inhibits invasion and proliferation of bladder cancer by regulating EMT. Int. J. Clin. Exp. Pathol. 2014, 7, 7653–7662. [Google Scholar]
- Wang, J.; Zhao, X.; Shi, J.; Pan, Y.; Chen, Q.; Leng, P.; Wang, Y. miR-451 suppresses bladder cancer cell migration and invasion via directly targeting c-Myc. Oncol. Rep. 2016, 36, 2049–2058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Itesako, T.; Seki, N.; Yoshino, H.; Chiyomaru, T.; Yamasaki, T.; Hidaka, H.; Yonezawa, T.; Nohata, N.; Kinoshita, T.; Nakagawa, M.; et al. The microRNA expression signature of bladder cancer by deep sequencing: The functional significance of the miR-195/497 cluster. PLoS ONE 2014, 9, e84311. [Google Scholar] [CrossRef]
- Guo, Y.; Ying, L.; Tian, Y.; Yang, P.; Zhu, Y.; Wang, Z.; Qiu, F.; Lin, J. miR-144 downregulation increases bladder cancer cell proliferation by targeting EZH2 and regulating Wnt signaling. FEBS J. 2013, 280, 4531–4538. [Google Scholar] [CrossRef]
- Pardini, B.; Cordero, F.; Naccarati, A.; Viberti, C.; Birolo, G.; Oderda, M.; Di Gaetano, C.; Arigoni, M.; Martina, F.; Calogero, R.; et al. microRNA profiles in urine by next-generation sequencing can stratify bladder cancer subtypes. Oncotarget 2018, 9, 20658–20669. [Google Scholar] [CrossRef]
- Dickman, C.T.; Lawson, J.; Jabalee, J.; MacLellan, S.A.; LePard, N.E.; Bennewith, K.L.; Garnis, C. Selective extracellular vesicle exclusion of miR-142-3p by oral cancer cells promotes both internal and extracellular malignant phenotypes. Oncotarget 2017, 8, 15252–15266. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Qin, F.; Hu, F.; Xu, H.; Sun, G.; Han, G.; Wang, T.; Guo, M. Characterization and selective incorporation of small non-coding RNAs in non-small cell lung cancer extracellular vesicles. Cell Biosci. 2018, 8, 2. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Li, L.-X.; Diao, Y.-J.; Wang, J.; Ye, Y.; Hao, X.-K. Identification of Urinary Exosomal miRNAs for the Non-Invasive Diagnosis of Prostate Cancer. Cancer Manag. Res. 2021, 13, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Kanaoka, R.; Iinuma, H.; Dejima, H.; Sakai, T.; Uehara, H.; Matsutani, N.; Kawamura, M. Usefulness of Plasma Exosomal MicroRNA-451a as a Noninvasive Biomarker for Early Prediction of Recurrence and Prognosis of Non-Small Cell Lung Cancer. Oncology 2018, 94, 311–323. [Google Scholar] [CrossRef]
- Goto, T.; Fujiya, M.; Konishi, H.; Sasajima, J.; Fujibayashi, S.; Hayashi, A.; Utsumi, T.; Sato, H.; Iwama, T.; Ijiri, M.; et al. An elevated expression of serum exosomal microRNA-191,− 21,− 451a of pancreatic neoplasm is considered to be efficient diagnostic marker. BMC Cancer 2018, 18, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Tian, X.; Han, R.; Zhang, X.; Wang, X.; Shen, H.; Xue, L.; Liu, Y.; Yan, X.; Shen, J.; et al. Downregulation of miR-486-5p contributes to tumor progression and metastasis by targeting protumorigenic ARHGAP5 in lung cancer. Oncogene 2013, 33, 1181–1189. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Mou, Q.; Li, P.; Yang, Z.; Wang, Z.; Niu, J.; Liu, Y.; Sun, Z.; Lv, S.; Zhang, B.; et al. MiR-486-5p inhibits IL-22-induced epithelial-mesenchymal transition of breast cancer cell by repressing Dock1. J. Cancer 2019, 10, 4695–4706. [Google Scholar] [CrossRef]
- Gao, Z.-J.; Yuan, W.-D.; Yuan, J.-Q.; Yuan, K.; Wang, Y. miR-486-5p functions as an oncogene by targeting PTEN in non-small cell lung cancer. Pathol. Res. Pr. 2018, 214, 700–705. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Shen, Y.-Q.; Li, Y.-L.; Liang, C.; Zhang, B.-J.; Lu, S.-D.; He, Y.-Y.; Wang, P.; Sun, Q.-L.; Jin, Y.-X.; et al. Direct repression of the oncogene CDK4 by the tumor suppressor miR-486-5p in non-small cell lung cancer. Oncotarget 2016, 7, 34011–34021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Zhang, T.; Yang, K.; Zhang, M.; Wang, K. miR-486-5p suppresses prostate cancer metastasis by targeting Snail and regulating epithelial–mesenchymal transition. OncoTargets Ther. 2016, 9, 6909–6914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Ji, C.; Guo, S.; Su, X.; Zhao, X.; Zhang, S.; Liu, G.; Qiu, X.; Zhang, Q.; Guo, H.; et al. The miR-486-5p plays a causative role in prostate cancer through negative regulation of multiple tumor suppressor pathways. Oncotarget 2017, 8, 72835–72846. [Google Scholar] [CrossRef] [Green Version]
- Salimian, J.; Baradaran, B.; Jamalkandi, S.A.; Moridikia, A.; Ahmadi, A. MiR-486-5p enhances cisplatin sensitivity of human muscle-invasive bladder cancer cells by induction of apoptosis and down-regulation of metastatic genes. Urol. Oncol. Semin. Orig. Investig. 2020, 38, 738.e9–738.e21. [Google Scholar] [CrossRef]
- Okamura, S.; Yoshino, H.; Kuroshima, K.; Tsuruda, M.; Osako, Y.; Sakaguchi, T.; Yonemori, M.; Yamada, Y.; Tatarano, S.; Nakagawa, M.; et al. EHHADH contributes to cisplatin resistance through regulation by tumor-suppressive microRNAs in bladder cancer. BMC Cancer 2021, 21, 1–13. [Google Scholar] [CrossRef]
- Kim, Y.; Roh, S.; Lawler, S.; Friedman, A. miR451 and AMPK Mutual Antagonism in Glioma Cell Migration and Proliferation: A Mathematical Model. PLoS ONE 2011, 6, e28293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Godlewski, J.; Bronisz, A.; Nowicki, M.O.; Chiocca, E.A.; Lawler, S. microRNA-451: A conditional switch controlling glioma cell proliferation and migration. Cell Cycle 2010, 9, 2742–2820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Xu, Z.; Kong, L.; Gao, H.; Zhang, Y.; Zheng, Y.; Wan, Y. miRNA-486-5p Promotes COPD Progression by Targeting HAT1 to Regulate the TLR4-Triggered Inflammatory Response of Alveolar Macrophages. Int. J. Chronic Obstr. Pulm. Dis. 2020, 15, 2991–3001. [Google Scholar] [CrossRef]
- Sabo, A.A.; Birolo, G.; Naccarati, A.; Dragomir, M.P.; Aneli, S.; Allione, A.; Oderda, M.; Allasia, M.; Gontero, P.; Sacerdote, C.; et al. Small Non-Coding RNA Profiling in Plasma Extracellular Vesicles of Bladder Cancer Patients by Next-Generation Sequencing: Expression Levels of miR-126-3p and piR-5936 Increase with Higher Histologic Grades. Cancers 2020, 12, 1507. [Google Scholar] [CrossRef] [PubMed]
- De Long, J.; Sullivan, T.B.; Humphrey, J.; Logvinenko, T.; A Summerhayes, K.; Kozinn, S.; Harty, N.; Summerhayes, I.C.; A Libertino, J.; Holway, A.H.; et al. A non-invasive miRNA based assay to detect bladder cancer in cell-free urine. Am. J. Transl. Res. 2015, 7, 2500–2509. [Google Scholar]
- Rekker, K.; Saare, M.; Roost, A.M.; Kubo, A.-L.; Zarovni, N.; Chiesi, A.; Salumets, A.; Peters, M. Comparison of serum exosome isolation methods for microRNA profiling. Clin. Biochem. 2014, 47, 135–138. [Google Scholar] [CrossRef]
- Liu, B.; Sun, W.; Gao, W.; Li, L.; Cao, Z.; Yang, X.; Liu, J.; Guo, Y. microRNA-451a promoter methylation regulated by DNMT3B expedites bladder cancer development via the EPHA2/PI3K/AKT axis. BMC Cancer 2020, 20, 1019. [Google Scholar] [CrossRef] [PubMed]
- Vu, L.; Ragupathy, V.; Kulkarni, S.; Atreya, C. Analysis of Argonaute 2-microRNA complexes in ex vivo stored red blood cells. Transfusion 2017, 57, 2995–3000. [Google Scholar] [CrossRef] [PubMed]
- Gebert, D.; Hewel, C.; Rosenkranz, D. unitas: The universal tool for annotation of small RNAs. BMC Genom. 2017, 18, 1–14. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genom. Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, G.E.; Schadt, E.E. variancePartition: Interpreting drivers of variation in complex gene expression studies. BMC Bioinform. 2016, 17, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sex | Age | Smoking Status | Primary Diagnosis | Diagnosis at TOI | |
|---|---|---|---|---|---|
| Patient 1 | M | 74 | PREVIOUS | TaG1 | TaG1 |
| Patient 2 | M | 76 | PREVIOUS | TaG1 | TaG1 |
| Patient 3 | F | 77 | PREVIOUS | TaG1 (2015) | TaG1 |
| Patient 4 | M | 61 | PREVIOUS | TaG1 | TaG1 |
| Patient 5 | M | 62 | NEVER | TaG1 | TaG1 |
| Patient 6 | M | 59 | PREVIOUS | TaG1 | TaG1 |
| Patient 7 | F | 61 | PREVIOUS | TaG2 | TaG2 |
| Patient 8 | M | 52 | PREVIOUS | TaG2 | TaG2 |
| Patient 9 | M | 81 | NEVER | TaG2 (2016) | TaG2 |
| Patient 10 | M | 81 | PREVIOUS | TaG2 (2012) | TaG2 |
| Patient 11 | M | 82 | PREVIOUS | TaG2 | TaG2 |
| Patient 12 | M | 72 | PREVIOUS | TaG1 | TaG1 |
| Patient 13 | M | 72 | PREVIOUS | TaG1 (1990) | TaG1 |
| Patient 14 | M | 65 | PREVIOUS | TaG1 | TaG1 |
| Patient 15 | M | 65 | PREVIOUS | TaG2 (2016) | TaG2 |
| Patient 16 | M | 72 | PREVIOUS | TaG2 (2011) | T1G3 |
| Patient 17 | M | 72 | PREVIOUS | T1G3 | T1G3 |
| Patient 18 | M | 74 | NEVER | T1G2 | T1G2 |
| Patient 19 | M | 70 | NEVER | T1G3 | T1G3 |
| Patient 20 | M | 82 | NEVER | T1G3 | T1G3 |
| Patient 21 | M | 47 | NEVER | T1G3/Carcinoma in-situ | T1G3/Carcinoma in-situ |
| Patient 22 | M | 72 | PREVIOUS | T1G3 | T1G3 |
| Patient 23 | F | 70 | NEVER | T1G3 | T1G3 |
| Patient 24 | F | 70 | NEVER | T1G2 | T1G2 |
| Patient 25 | M | 81 | PREVIOUS | TaG2 (2005) | TaG2 |
| Patient 26 | M | 81 | PREVIOUS | TaG3 | TaG3 |
| Patient 27 | M | 75 | PREVIOUS | T1G2 | T1G2 |
| Patient 28 | F | 80 | PREVIOUS | TaG2 (2012) | TaG2 |
| Patient 29 | F | 65 | PREVIOUS | TaG1 | TaG1 |
| Patient 30 | M | 82 | NEVER | TaG2 | TaG2 |
| Patient 31 | F | 83 | NEVER | TaG1 | TaG1 |
| Patient 32 | F | 71 | NEVER | TaG2 | TaG2 |
| Patient 33 | F | 77 | PREVIOUS | TaG2 (2008) | TaG2 |
| Patient 34 | M | 75 | NEVER | TaG2 | TaG2 |
| Patient 35 | M | 79 | CURRENT | TaG2 | TaG2 |
| Patient 36 | M | 79 | PREVIOUS | TaG1 | TaG1 |
| Patient 37 | M | 69 | PREVIOUS | TaG2 | TaG2 |
| Patient 38 | M | 70 | PREVIOUS | TaG3 (2018) | T1G3 |
| Patient 39 | M | 64 | PREVIOUS | T1G3 | T1G3 |
| Patient 40 | M | 78 | PREVIOUS | T1G3 | T1G3 |
| Patient 41 | M | 89 | PREVIOUS | T1G2 | T1G2 |
| (A) Urine EVs | (B) Serum Supernatant | ||
|---|---|---|---|
| miR-486-5p | 12 | miR-3158-3p | 9 |
| miR-451a | 12 | miR-3921-3p | 7 |
| miR-205-5p | 11 | miR-4654-3p | 9 |
| miR-1246 | 10 | miR-922-3p | 11 |
| miR-483-3p | 12 | miR-612-3p | 7 |
| miR-1323 | 14 | miR-4453-3p | 8 |
| miR-6073-3p | 11 | miR-942-3p | 7 |
| miR-202-3p | 8 | miR-4734-3p | 9 |
| miR-4508-5p | 8 | miR-185-3p | 8 |
| miR-483-5p | 9 | miR-6851-5p | 9 |
| miR-6768-5p | 8 | miR-135a-1-3p | 3 |
| miR-3153-3p | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Strømme, O.; Heck, K.A.; Brede, G.; Lindholm, H.T.; Otterlei, M.; Arum, C.-J. Differentially Expressed Extracellular Vesicle-Contained microRNAs before and after Transurethral Resection of Bladder Tumors. Curr. Issues Mol. Biol. 2021, 43, 286-300. https://doi.org/10.3390/cimb43010024
Strømme O, Heck KA, Brede G, Lindholm HT, Otterlei M, Arum C-J. Differentially Expressed Extracellular Vesicle-Contained microRNAs before and after Transurethral Resection of Bladder Tumors. Current Issues in Molecular Biology. 2021; 43(1):286-300. https://doi.org/10.3390/cimb43010024
Chicago/Turabian StyleStrømme, Olaf, Kathleen A. Heck, Gaute Brede, Håvard T. Lindholm, Marit Otterlei, and Carl-Jørgen Arum. 2021. "Differentially Expressed Extracellular Vesicle-Contained microRNAs before and after Transurethral Resection of Bladder Tumors" Current Issues in Molecular Biology 43, no. 1: 286-300. https://doi.org/10.3390/cimb43010024
APA StyleStrømme, O., Heck, K. A., Brede, G., Lindholm, H. T., Otterlei, M., & Arum, C. -J. (2021). Differentially Expressed Extracellular Vesicle-Contained microRNAs before and after Transurethral Resection of Bladder Tumors. Current Issues in Molecular Biology, 43(1), 286-300. https://doi.org/10.3390/cimb43010024