
Three-Dimensional Reconstructed Bone Marrow Matrix Culture Improves the Viability of Primary Myeloma Cells In-Vitro via a STAT3-Dependent Mechanism

 ,
, 
Abstract
:1. Introduction
2. Results
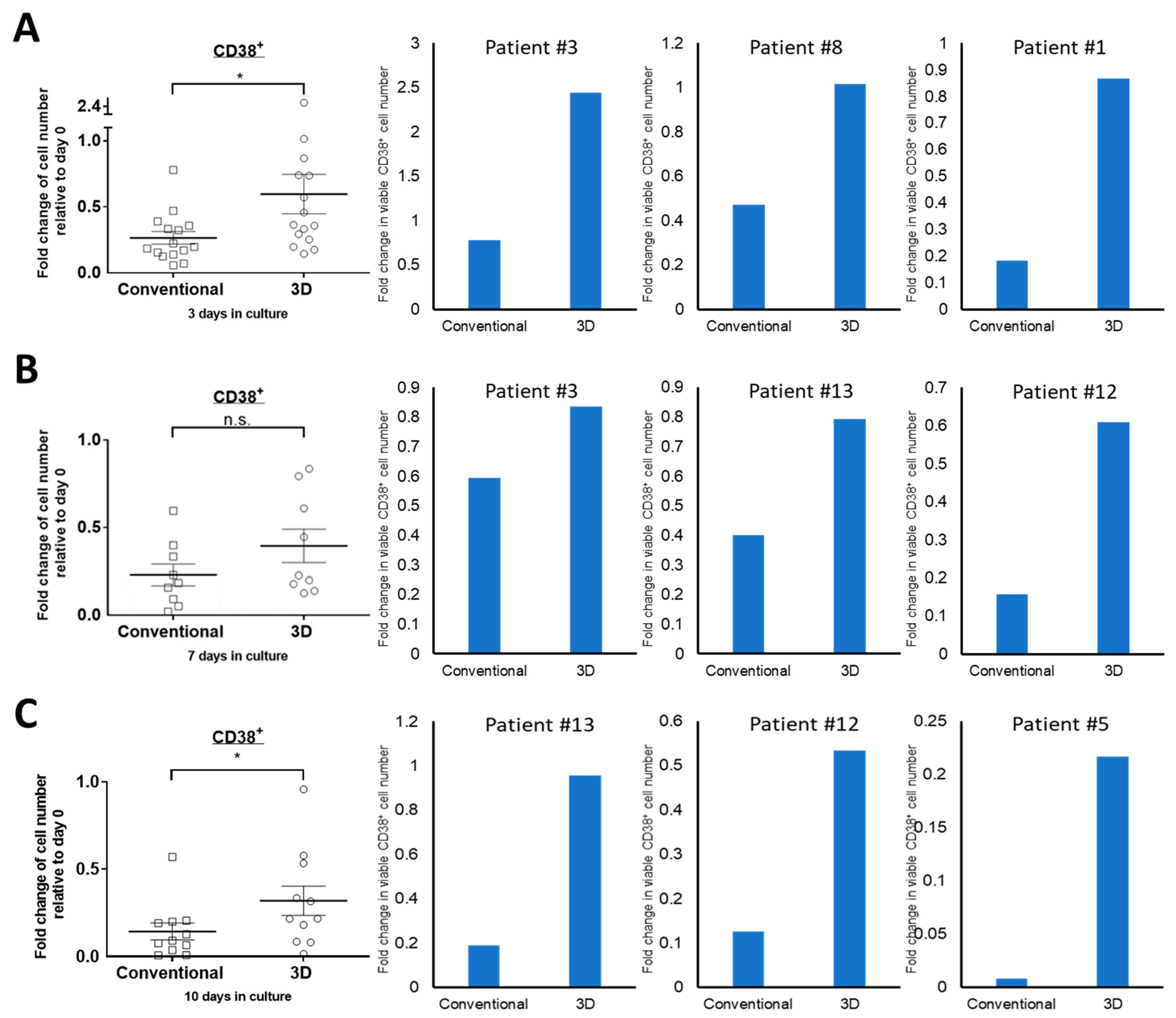
2.1. The Viability of Primary MM Cells Is Higher in 3D Culture
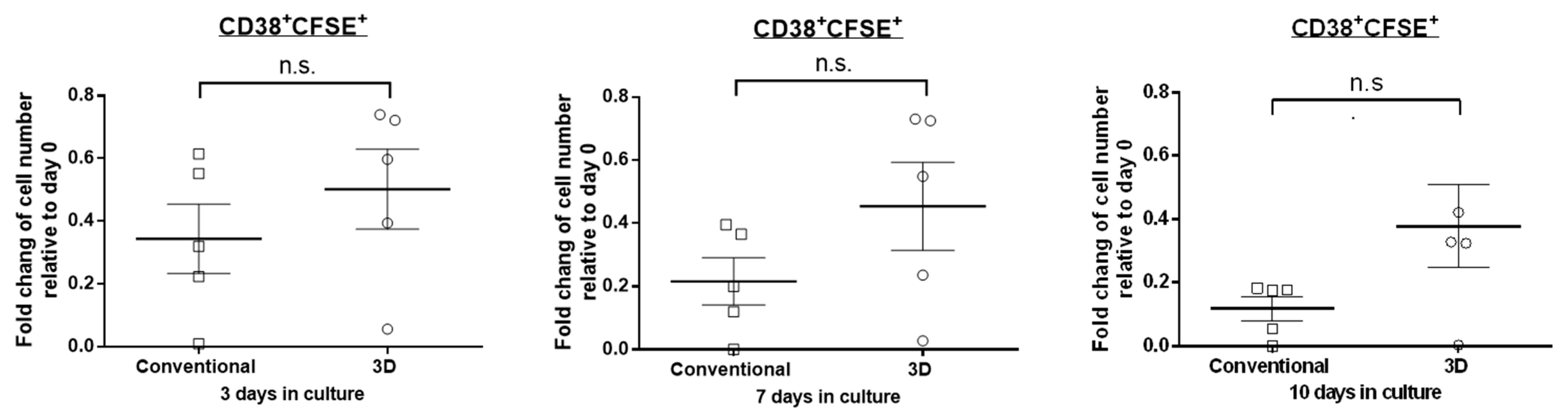
2.2. The Proliferation Rate of MM Cells Is Similar between 3D and Conventional Culture
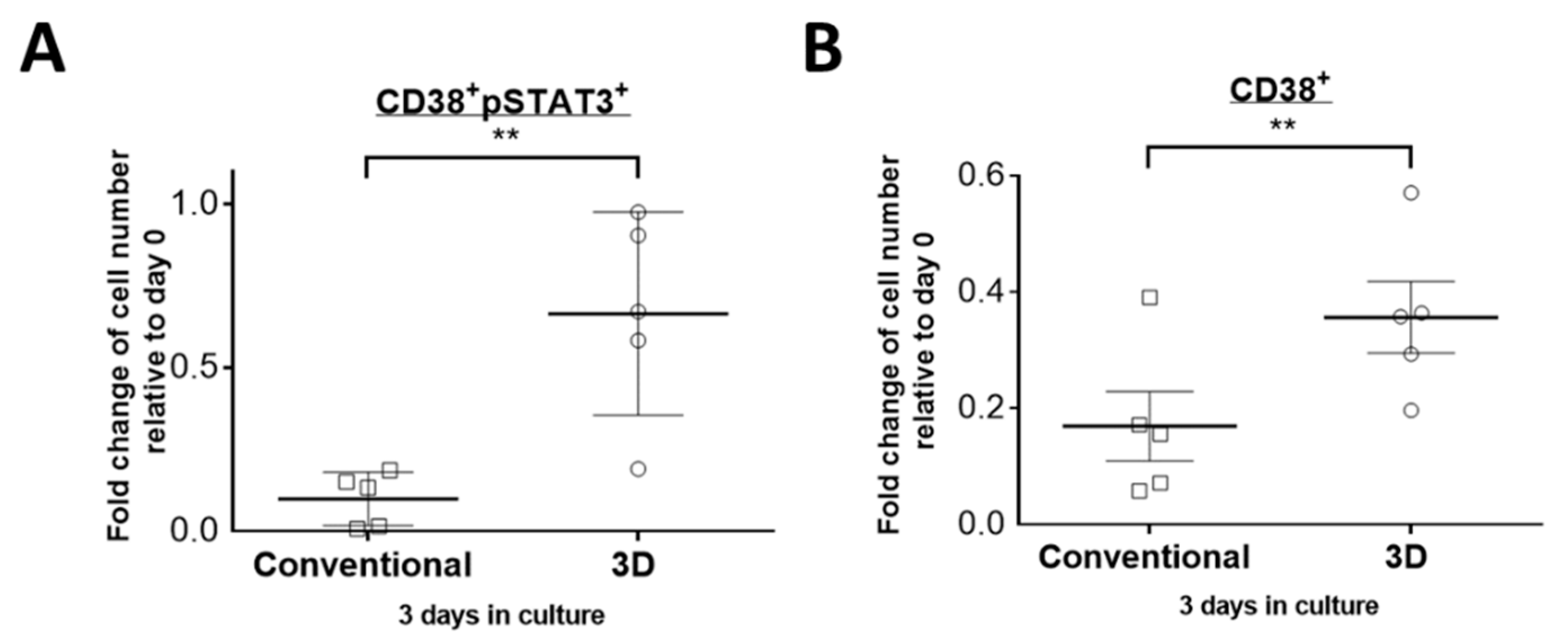
2.3. STAT3 Is More Active in PM Cells Cultured in 3D Culture
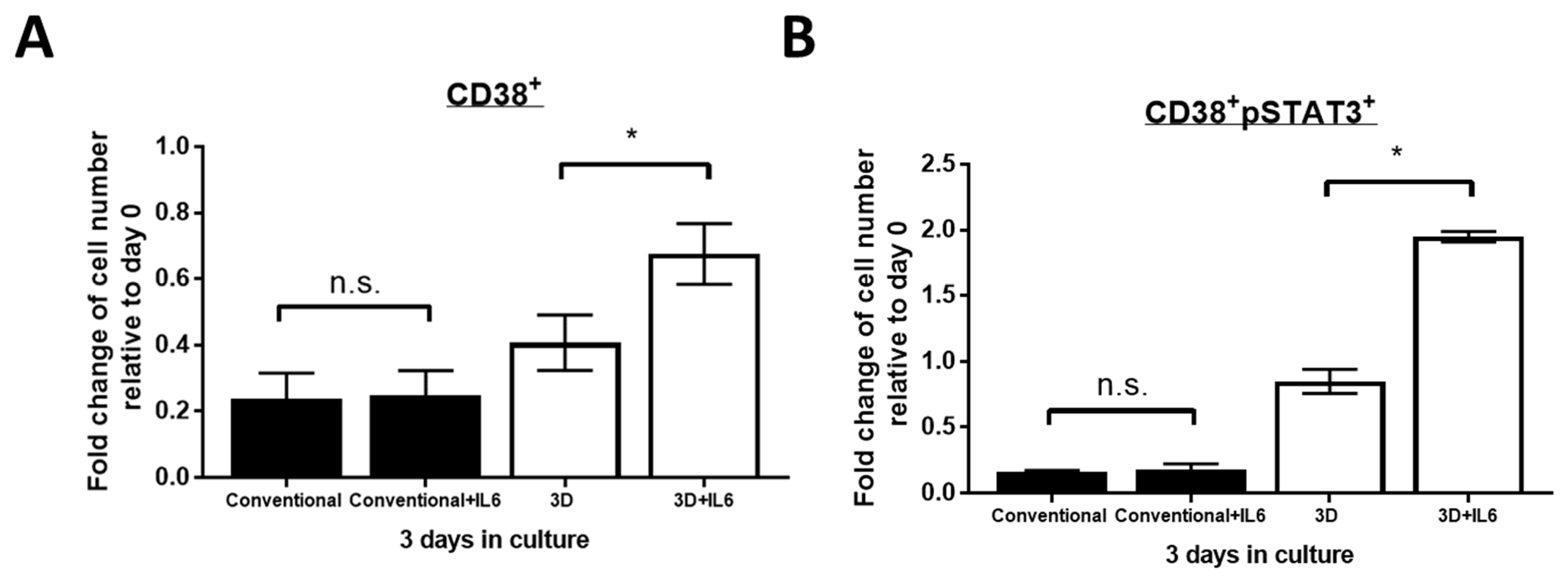
2.4. IL6 Further Improved PM Cell Viability in 3D but Not Conventional Culture
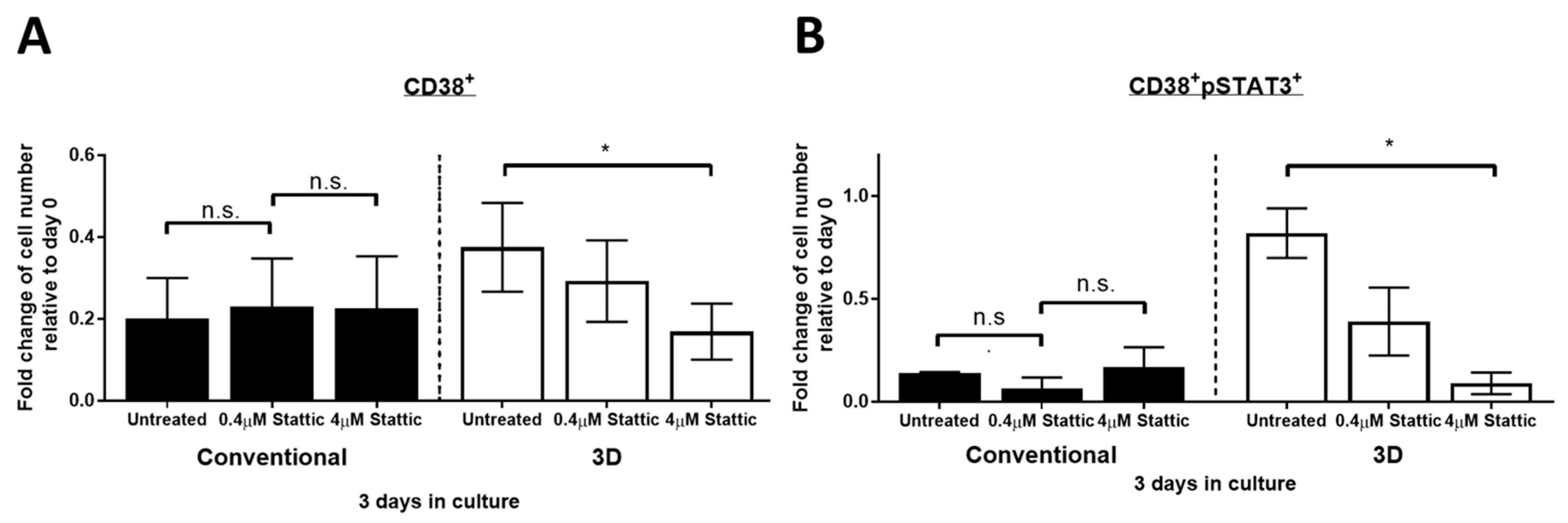
2.5. Stattic Inhibits Cell Growth of PM in 3D Culture but Not in Conventional Culture
3. Discussion
4. Materials and Methods
4.1. Patient Samples, 3D Culture and Stattic/IL6 Treatment
4.2. Flow Cytometry Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, Y.; Liu, Y.; Zheng, C.; Shen, C. Investigation of cross-contamination and misidentification of 278 widely used tumor cell lines. PLoS ONE 2017, 12, e0170384. [Google Scholar] [CrossRef]
- Matsuo, Y.; Drexler, H.G.; Kaufmann, M.; Milch, H.; MacLeod, R.A.F.; Dirks, W.G. Widespread intraspecies cross-contamination of human tumor cell lines arising at source. Int. J. Cancer 2002, 83, 555. [Google Scholar]
- Sun, Y.; Liu, Q. Deciphering the Correlation between Breast Tumor Samples and Cell Lines by Integrating Copy Number Changes and Gene Expression Prfiles. Biomed. Res. Int. 2015, 2015, 901303. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Walling, J.; Kotliarov, Y.; Center, A.; Steed, M.E.; Ahn, S.J.; Rosenblum, M.; Mikkelsen, T.; Zenklusen, J.C.; Fine, H.A. Genomic Changes and Gene Expression Profiles Reveal That Established Glioma Cell Lines Are Poorly Representative of Primary Human Gliomas. Mol. Cancer Res. 2008, 6, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gillet, J.P.; Varma, S.; Gottesman, M.M. The clinical relevance of cancer cell lines. J. Natl. Cancer Inst. 2013, 105, 452–458. [Google Scholar] [CrossRef] [Green Version]
- Marx, V. Cell-line authentication demystified. Nat. Methods 2014, 11, 483–488. [Google Scholar] [CrossRef]
- Fernando, R.C.; de Carvalho, F.; Mazzotti, D.R.; Evangelista, F.; Moisés, W.; Braga, T.; Colleoni, G.W.B. Multiple myeloma cell lines and primary tumors proteoma: Protein biosynthesis and Immune system as potential therapeutic targets. Genes Cancer 2015, 6, 462. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, G.; Munshi, N. Pathogenesis beyond the cancer clone (s) in multiple myeloma. Blood 2015, 125, 3049–3059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nijhof, I.S.; Groen, R.W.J.; Noort, W.A.; VanKessel, B.; DeJong-Korlaar, R.; Bakker, J.; Mutis, T. Preclinical evidence for the therapeutic potential of CD38-Targeted Immuno-chemotherapy in multiple Myeloma patients refractory to Lenalidomide and Bortezomib. Clin. Cancer Res. 2015, 21, 2802–2810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van derVeer, M.S.; de Weers, M.; van Kessel, B.; Bakker, J.M.; Wittebol, S.; Parren, P.W.H.I.; Lokhorst, H.M.; Mutis, T. Towards effective immunotherapy of myeloma: Enhanced elimination of myeloma cells by combination of lenalidomide with the human CD38 monoclonal antibody daratumumab. Haematologica 2011, 96, 284–290. [Google Scholar] [CrossRef] [Green Version]
- Zlei, M.; Egert, S.; Wider, D.; Ihorst, G.; Wäsch, R.; Engelhardt, M. Characterization of in vitro growth of multiple myeloma cells. Exp. Hematol. 2007, 35, 1550–1561. [Google Scholar] [CrossRef]
- Weiswald, L.B.; Bellet, D.; Dangles-Marie, V. Spherical Cancer Models in Tumor Biology. Neoplasia 2015, 17, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Kirshner, J.; Thulien, K.J.; Martin, L.D.; Debes Marun, C.; Reiman, T.; Belch, A.R.; Pilarski, L.M. A unique three-dimensional model for evaluating the impact of therapy on multiple myeloma. Blood 2008, 112, 2935–2945. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.-H.H.; Molavi, O.; Alshareef, A.; Haque, M.; Wang, Q.; Chu, M.P.; Lai, R. Constitutive activation of STAT3 in myeloma cells cultured in a three-dimensional, reconstructed bone marrow model. Cancers 2018, 10, 206. [Google Scholar] [CrossRef] [Green Version]
- Bharti, A.C.; Shishodia, S.; Reuben, J.M.; Weber, D.; Alexanian, R.; Raj-Vadhan, S.; Estrov, Z.; Talpaz, M.; Aggarwal, B.B. Nuclear factor-κB and STAT3 are constitutively active in CD138 + cells derived from multiple myeloma patients, and suppression of these transcription factors leads to apoptosis. Blood 2004, 103, 3175–3184. [Google Scholar] [CrossRef] [PubMed]
- Brown, R.; Yang, S.; Weatherburn, C.; Gibson, J.; Ho, P.J.; Suen, H.; Hart, D.; Joshua, D. Phospho-flow detection of constitutive and cytokine-induced pSTAT3/5, pAKT and pERK expression highlights novel prognostic biomarkers for patients with multiple myeloma. Leukemia 2015, 29, 483–490. [Google Scholar] [CrossRef]
- Quintanilla-Martinez, L.; Kremer, M.; Specht, K.; Calzada-Wack, J.; Nathrath, M.; Schaich, R.; Höfler, H.; Fend, F. Analysis of signal transducer and activator of transcription 3 (Stat 3) pathway in multiple myeloma: Stat 3 activation and cyclin D1 dysregulation are mutually exclusive events. Am. J. Pathol. 2003, 162, 1449–1461. [Google Scholar] [CrossRef]
- Chong, P.S.Y.; Chng, W.J.; de Mel, S. STAT3: A promising therapeutic target in multiple myeloma. Cancers 2019, 11, 731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamińska, J.; Koper, O.M.; Dymicka-Piekarska, V.; Motybel, E.; Kłoczko, J.; Ke-Mona, H. Angiogenic cytokines: IL-6, sIL-6R, TNF-α, sVCAM-1, and PDGF-AB in multiple myeloma patients depending on the stage of the disease. Edorium. J. Tumor Biol. Edorium. J. Tumor Bio. 2015, 22, 11–19. [Google Scholar]
- Holien, T.; Misund, K.; Olsen, O.E.; Baranowska, K.A.; Buene, G.; Børset, M.; Waage, A.; Sundan, A. Correction: MYC amplifications in myeloma cell lines: Correlation with MYC-inhibitor efficacy. Oncotarget 2018, 9, 36048. [Google Scholar] [CrossRef]
- Vincent, L.; Jin, D.K.; Karajannis, M.A.; Shido, K.; Hooper, A.T.; Rashbaum, W.K.; Pytowski, B.; Wu, Y.; Hicklin, D.J.; Zhu, Z.; et al. Fetal Stromal–Dependent Paracrine and Intracrine Vascular Endothelial Growth Factor-A/Vascular Endothelial Growth Factor Receptor-1 Signaling Promotes Proliferation and Motility of Human Primary Myeloma Cells. Cancer Res. 2005, 65, 3185–3192. [Google Scholar] [CrossRef] [Green Version]
- Quinn, J.; Glassford, J.; Percy, L.; Munson, P.; Marafioti, T.; Rodriguez-Justo, M.; Yong, K. APRIL promotes cell-cycle progression in primary multiple myeloma cells: Influence of D-type cyclin group and translocation status. Blood 2010, 117, 890–901. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Lee, W.Y.; Siegel, D.S.; Tolias, P.; Zilberberg, J. Patient-Specific 3D Microfluidic Tissue Model for Multiple Myeloma. Tissue Eng. Part C Methods 2013, 20, 663–670. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de la Puente, P.; Muz, B.; Gilson, R.; Azab, F.; Luderer, M.; King, J.; Achilefu, S.; Vij, R.; Azab, A.K. 3D tissue-engineered bone marrow as a novel model to study pathophysiology and drug resistance in multiple myeloma. Biomaterials 2015, 73, 70–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braham, M.V.J.; Minnema, M.C.; Aarts, T.; Sebestyen, Z.; Straetemans, T.; Vyborova, A.; Kuball, J.; Öner, F.C.; Robin, C.; Alblas, J. Cellular immunotherapy on primary multiple myeloma expanded in a 3D bone marrow niche model. OncoImmunology 2018, 7, e1434465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alexandrakis, M.G.; Passam, F.H.; Kyriakou, D.S.; Dambaki, K.; Niniraki, M.; Stathopoulos, E. Ki-67 Proliferation Index: Correlation with Prognostic Parameters and Outcome in Multiple Myeloma. Am. J. Clin. Oncol. Cancer Clin. Trials 2004, 27, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Gu, Y.; Sun, Q.; Siegel, D.S.; Tolias, P.; Yang, Z.; Lee, W.O.; Zilberberg, J. Ex Vivo Maintenance of Primary Human Multiple Myeloma Cells through the Optimization of the Osteoblastic Niche. PLoS ONE 2015, 10, e0125995. [Google Scholar] [CrossRef]
- Catlett-Falcone, R.; Landowski, T.H.; Oshiro, M.M.; Turkson, J.; Levitzki, A.; Savino, R.; Ciliberto, G.; Moscinski, L.; Fernández-Luna, J.L.; Nuñez, G.; et al. Constitutive Activation of Stat3 Signaling Confers Resistance to Apoptosis in Human U266 Myeloma Cells. Immunity 1999, 10, 105–115. [Google Scholar] [CrossRef] [Green Version]
- Abroun, S.; Ishikawa, H.; Tsuyama, N.; Liu, S.; Li, F.J.; Otsuyama, K.I.; Zheng, X.; Obata, M.; Kawano, M.M. Receptor synergy of interleukin-6 (IL-6) and insulin-like growth factor-I that highly express IL-6 receptor α myeloma cells. Blood 2004, 103, 2291–2298. [Google Scholar] [CrossRef] [Green Version]
- Jung, S.-H.; Ahn, S.-Y.; Choi, H.-W.; Shin, M.-G.; Lee, S.-S.; Yang, D.-H.; Ahn, J.-S.; Kim, Y.-K.; Kim, H.-J.; Lee, J.-J. STAT3 expression is associated with poor survival in non-elderly adult patients with newly diagnosed multiple myeloma. Blood Res. 2017, 52, 293–299. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.; Wang, Z.; Li, Z.-J.; Peng, H.; Luo, Y.; Deng, M.; Li, R.; Dai, C.; Xu, Y.; Liu, S.; et al. Icaritin suppresses multiple myeloma, by inhibiting IL-6/JAK2/STAT3. Oncotarget 2015, 6, 10460–10472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, L.; Benson, D.M.; DeAngelis, S.; Bakan, C.E.; Li, P.-K.; Li, C.; Lin, J. A small molecule, LLL12 inhibits constitutive STAT3 and IL-6-induced STAT3 signaling and exhibits potent growth suppressive activity in human multiple myeloma cells. Int. J. Cancer 2011, 130, 1459–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soleimani, A.H.; Garg, S.; Paiva, I.M.; Vakili, M.R.; Alshareef, A.; Huang, Y.-H.; Molavi, O.; Lai, R.; Lavasanifar, A. Micellar nano-carriers for the delivery of STAT3 dimerization inhibitors to melanoma. Drug Deliv. Transl. Res. 2017, 7, 571–581. [Google Scholar] [CrossRef]
- Molavi, O.; Ma, Z.; Mahmud, A.; Alshamsan, A.; Samuel, J.; Lai, R.; Kwon, G.S.; Lavasanifar, A. Polymeric micelles for the solubilization and delivery of STAT3 inhibitor cucurbitacins in solid tumors. Int. J. Pharm. 2008, 347, 118–127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parikh, M.R.; Belch, A.R.; Pilarski, L.M.; Kirshner, J. A Three-dimensional Tissue Culture Model to Study Primary Human Bone Marrow and its Malignancies. J. Vis. Exp. 2014, 85, 50947. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient # | Age | Gender | Diagnosis | Cytogenetic | % Bone Cellularity | % Plasma Cells | Anemia Present? | Light Chain Type | Notes |
|---|---|---|---|---|---|---|---|---|---|
| 1 | 64 | Male | Initial | CCND1 | 20–30 | 80 | Yes | Kappa | - |
| 2 | 78 | Male | Initial | Gain CCND1 locus | 50 | 12 | No | Kappa | Plasmacytoma in ribs |
| 3 | 52 | Female | Initial | Not detected | 60 | 80 | Yes | Kappa | Plasmacytoma in femur |
| 4 | 61 | Female | Initial | Not detected | 60–70 | 50–60 | No | Kappa | - |
| 5 | 45 | Male | Initial | Not detected | 80–90 | 90 | Yes | Lambda | Plasmacytoma in ribs |
| 6 | 63 | Female | Initial | CCND1 | 80–90 | 70 | Yes | Kappa | - |
| 7 | 66 | Female | Initial | Trisomy 17, CCND1 | 95 | 60 | No | Lambda | Plasmacytoma in pelvis |
| 8 | 55 | Female | Relapse | CCND1 | 40 | 50 | No | Lambda | - |
| 9 | 73 | Female | Initial | CCND1 and IgH, MAF | 50 | 60 | No | Lambda | - |
| 10 | 54 | Male | Initial | CCND1 | 50 | 30 | No | Lambda | - |
| 11 | 78 | Male | Initial | Not detected | 50 | 10 | No | Lambda | - |
| 12 | 66 | Female | Initial | TP53 deletion | 35 | 10–20 | Yes | Lambda | Positive for amyloidosis, rib lesion |
| 13 | 64 | Male | Initial | Unknown | 50 | 50 | Yes | Kappa | Previous bone lesion |
| 14 | 46 | Female | Initial | Unknown | 70–80 | 15 | No | Lambda | - |
| 15 | 77 | Male | Initial | Unknown | 60–70 | 70–80 | No | Kappa | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, Y.-H.; Almowaled, M.; Li, J.; Venner, C.; Sandhu, I.; Peters, A.; Lavasanifar, A.; Lai, R. Three-Dimensional Reconstructed Bone Marrow Matrix Culture Improves the Viability of Primary Myeloma Cells In-Vitro via a STAT3-Dependent Mechanism. Curr. Issues Mol. Biol. 2021, 43, 313-323. https://doi.org/10.3390/cimb43010026
Huang Y-H, Almowaled M, Li J, Venner C, Sandhu I, Peters A, Lavasanifar A, Lai R. Three-Dimensional Reconstructed Bone Marrow Matrix Culture Improves the Viability of Primary Myeloma Cells In-Vitro via a STAT3-Dependent Mechanism. Current Issues in Molecular Biology. 2021; 43(1):313-323. https://doi.org/10.3390/cimb43010026
Chicago/Turabian StyleHuang, Yung-Hsing, Meaad Almowaled, Jing Li, Christopher Venner, Irwindeep Sandhu, Anthea Peters, Afsaneh Lavasanifar, and Raymond Lai. 2021. "Three-Dimensional Reconstructed Bone Marrow Matrix Culture Improves the Viability of Primary Myeloma Cells In-Vitro via a STAT3-Dependent Mechanism" Current Issues in Molecular Biology 43, no. 1: 313-323. https://doi.org/10.3390/cimb43010026
APA StyleHuang, Y. -H., Almowaled, M., Li, J., Venner, C., Sandhu, I., Peters, A., Lavasanifar, A., & Lai, R. (2021). Three-Dimensional Reconstructed Bone Marrow Matrix Culture Improves the Viability of Primary Myeloma Cells In-Vitro via a STAT3-Dependent Mechanism. Current Issues in Molecular Biology, 43(1), 313-323. https://doi.org/10.3390/cimb43010026