Tasco®: A Product of Ascophyllum nodosum Enhances Immune Response of Caenorhabditis elegans Against Pseudomonas aeruginosa Infection

Abstract
:1. Introduction
2. Results
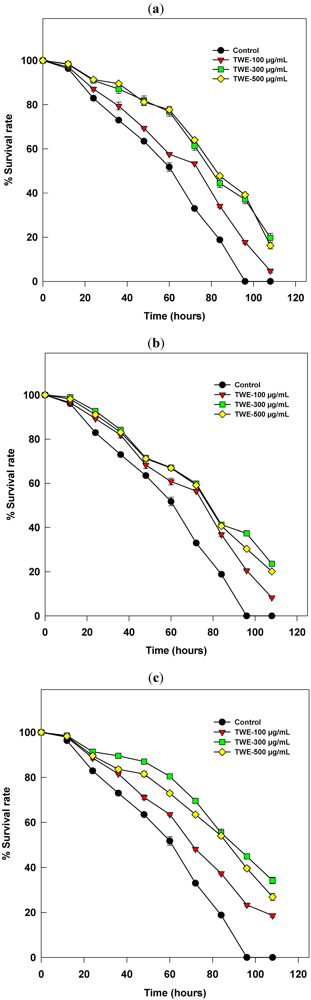
2.1. The C. elegans Killing Assay

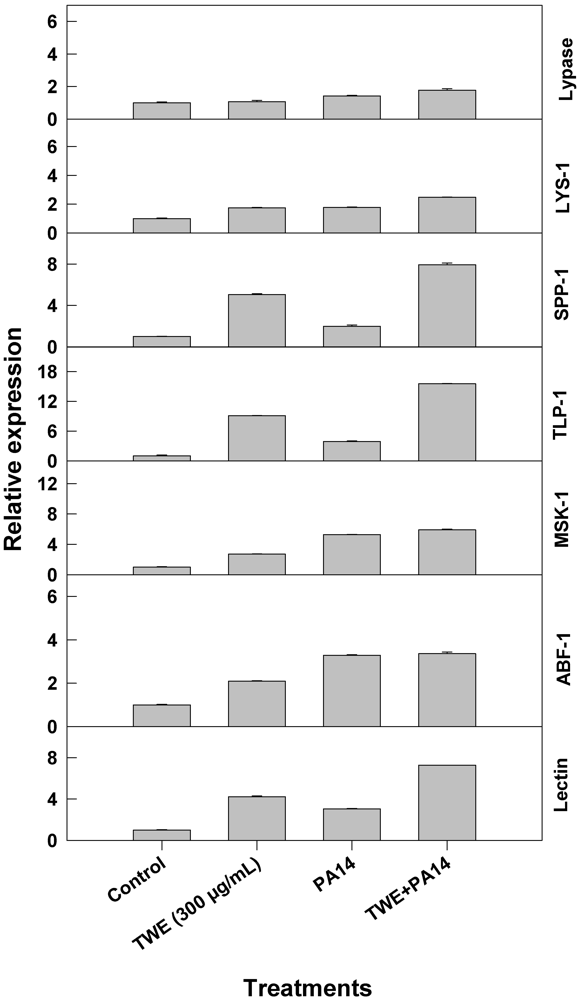
2.2. Effect of TWE-Treatment on Immune Responsive Gene Expression of C. elegans

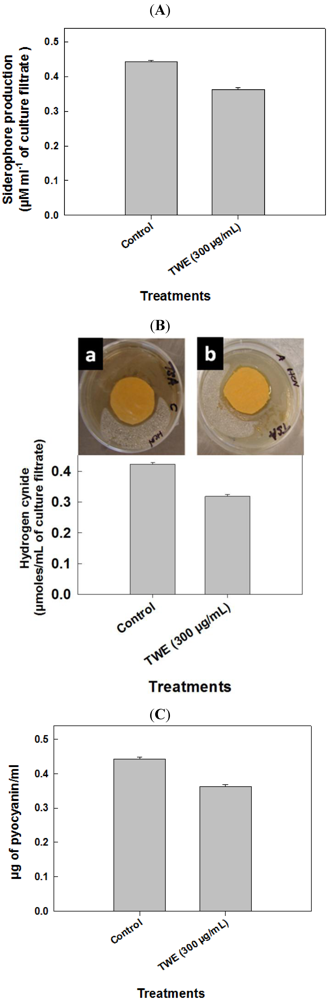
2.3. Effect of TWE-Treatment on P. aeruginosa Secreted Virulence Factors

2.4. Effect of TWE on P. aeruginosa Toxic Metabolites

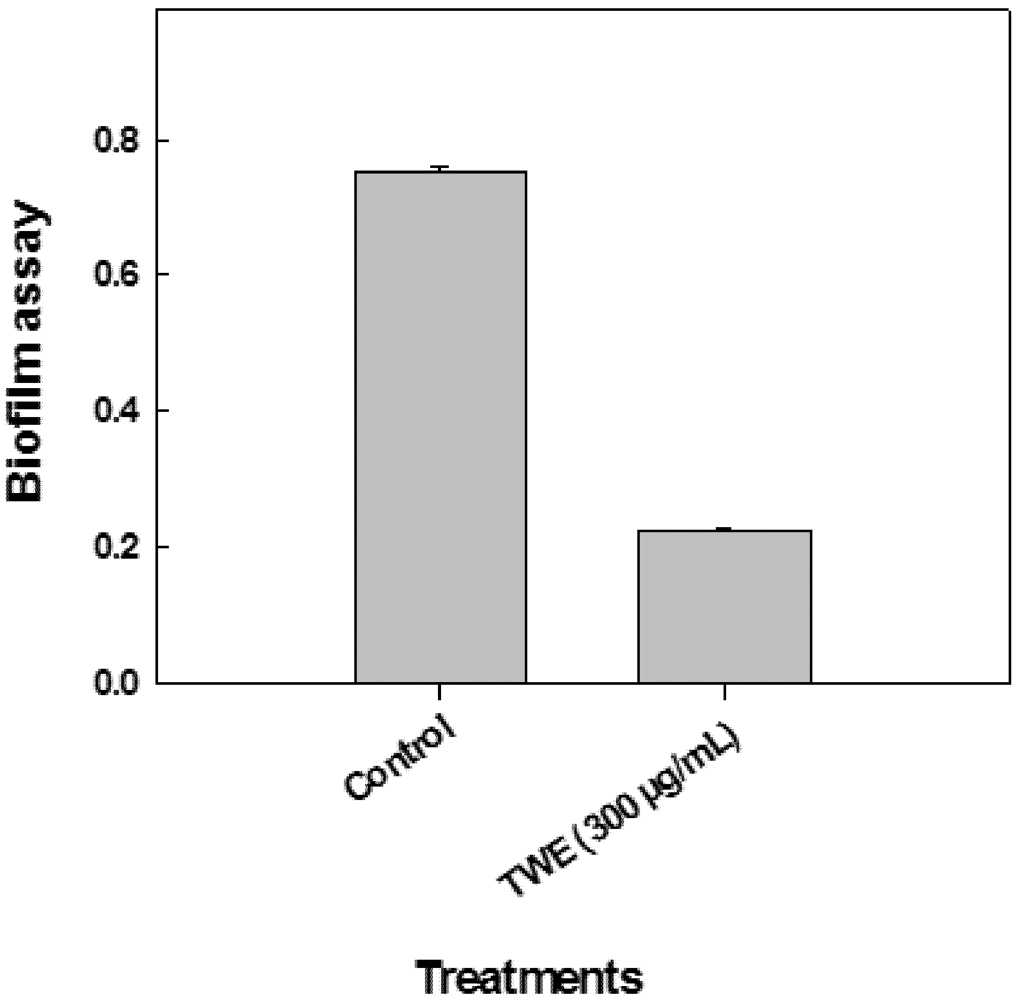
2.5. Effect of TWE on P. aeruginosa Biofilm Formation

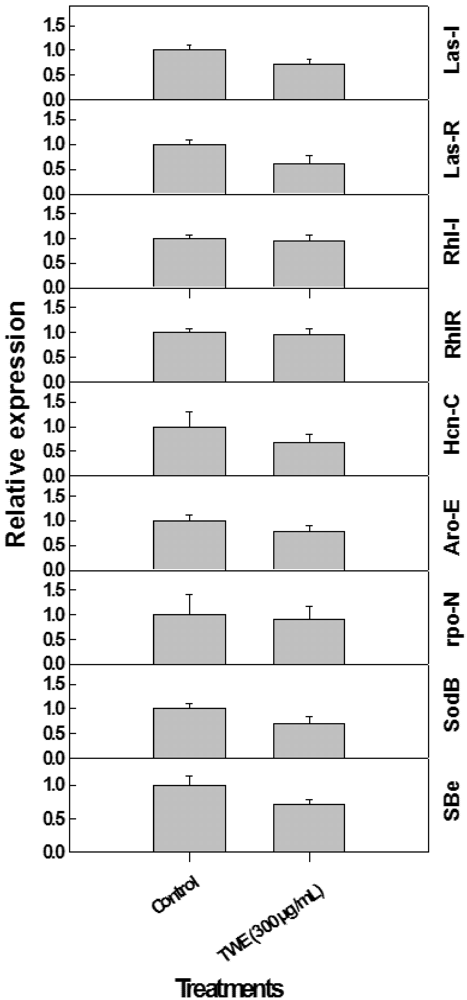
2.6. Effect of TWE on Expression of P. aeruginosa Quorum Sensing Genes

2.7. Antimicrobial Susceptibility Testing

3. Discussion
4. Experimental Section
4.1. Preparation of TWE
4.2. Bacterial Strains and Growth Conditions
4.3. C. elegans Killing Assay
4.4. Effect of TWE on Immune Response Gene Expression of C. elegans

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sl. No | Primer list | Sequence of oligonucleotides (5′→3′) |
|---|---|---|
| 1 | ZK6.7-F | CGAATTCCTCCCAAACAACT |
| ZK6.7-R | GAATAGGACGTTGTCGCAGA | |
| 2 | lys-1-F | TTCGGATCTTTCAAGAAGGC |
| lys-1-R | TGGGATTCCAACAACGTAAA | |
| 3 | spp-1-F | TGAACATCGGAACTCTTTGC |
| spp-1-R | TCAGCTCTTCCTCACACTCG | |
| 4 | F28D1.3-F | AATCTGGATGCCTCGGATAC |
| F28D1.3-R | CATCTGAGCAGTTGCAGAGC | |
| 5 | T20G5.7-F | ATGTTCTCCCTCAAGACCGT |
| T20G5.7-R | CGGAAGTGTAAACGACGAAG | |
| 6 | abf-1-F | TGCCTTCTCCTTGTTCTCCT |
| abf-1-R | ATCCTCTGCATTACCGGAAC | |
| 7 | F38A1.5-F | CTGGGCCGGTATTAATTTGT |
| F38A1.5-R | GTCTTCTTCGTCACGCACAT | |
| 8 | ama-1-F | CTGACCCAAAGAACACGGTGA |
| ama-1-R | TCCAATTCGATCCGAAGAAGC |
4.5. Effect of TWE on P. aeruginosa Protease
4.6. Effect of TWE on P. aeruginosa Alkaline Protease
4.7. Effect of TWE on P. aeruginosa Elastin
4.8. Effect of TWE on P. aeruginosa Pyocyanin
4.9. Effect of TWE on the Production of Hydrogen Cyanide in P. aeruginosa
4.10. Quantification of Hydrogen Cyanide
4.11. Quantification of Siderophore Production
4.12. RT-PCR Analysis of the Quorum Sensing and Virulence Genes
| Sl. No | Primer list | Sequence of oligonucleotides (5′→3′) |
|---|---|---|
| 1 | lasI-F | GCTCCTTGAACACTTGAGCA |
| lasI-R | GCGCGAAGAGTTCGATAAAA | |
| 2 | lasR-F | CCGCCGAATATTTCCCATA |
| lasR-R | GATATCGGTTATCTGCAACTGCT | |
| 3 | rhlI-F | GGAGCGCTATTTCGTTCG |
| rhlI-R | GTCTCGCCCTTGACCTTCT | |
| 4 | rhlR-F | TGCGTTGCATGATCGAGT |
| rhlR-R | CGGGTTGGACATCAGCAT | |
| 5 | hcnC-F | GCCTGGACAGTTGGTAGGC |
| hcnC-R | GAACAGAACCTATGACATCGTGA | |
| 6 | aroE-F | TTCTTCGAGCAGGGCAAG |
| aroE-R | CAATTCGTCCACCAGACGAT | |
| 7 | rpoN-F | ATACCTTCATGCGCAACCA |
| rpoN-R | GGCTCTGCAGGCTCTTGAT | |
| 8 | sbe-F | CTCGTTGGTCTCCTCGAGTT |
| sbe-R | CCATCTACCAGCGTGAAGG | |
| 9 | sodB-F | GTTCAAGGAAGAGTTCACCAAGA |
| sodB-R | GTCGGCCTTCTTCACCAG | |
| 10 | 16S rRNA-F | GATTAACGCTTGCACCCTTC |
| 16S rRNA-R | TAAGCACCGGCTAACTTCGT |
4.13. Biofilm Formation
4.14. Testing of Antimicrobial Susceptibility
4.15. Statistical Analysis
5. Conclusions
Acknowledgements
- Samples Availability: Available from the authors.
References
- Ugarte, R.; Craigie, J.S.; Critchley, A.T. Fucoid Flora of the Rocky Intertidal of the Canadian Maritimes: Implications for the Future with Rapid Climate Change. In Seaweeds and Their Role in Globally Changing Environments; Israel, A., Einav, R., Seckbach, J., Eds.; Springer: Berlin, Germany, 2010. [Google Scholar]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, B. Seaweed extracts as biostimulants of plant growth and development (review). J. Plant. Growth Regul. 2009, 28, 386–399. [Google Scholar]
- Sangha, J.S.; Hobson, D.; Hiltz, D.; Critchley, A.T.; Prithiviraj, B. The use of commercial seaweed extracts as a means to alleviate abiotic stress in land plants: A review. Algal Resour. 2010, 3, 153–168. [Google Scholar]
- Craigie, J.S. Seaweed extract stimuli in plant science and agriculture. J. Appl. Phycol. 2011, 23, 371–393. [Google Scholar]
- Fike, J.H.; Allen, V.G.; Schmidt, R.E.; Zhang, X.; Fontenot, J.P.; Bagley, C.P.; Ivy, R.L.; Evans, R.R.; Coelho, R.W.; Wester, D.B. Tasco-Forage: 1. Influence of a seaweed extract on antioxidant activity in tall fescue and in ruminants. J. Anim. Sci. 2001, 79, 1011–1021. [Google Scholar] [PubMed]
- Yeh, S.T.; Lee, C.S.; Chen, J.C. Administration of hot-water extract of brown seaweed Sargassum duplicatum via immersion and injection enhances the immune resistance of white shrimp Litopenaeus vannamei. Fish Shellfish Immunol. 2006, 20, 332–345. [Google Scholar]
- Kandasamy, S.; Fan, D.; Sangha, J.S.; Khan, W.; Evans, F.; Critchley, A.T.; Prithiviraj, B. Tasco®, a product of Ascophyllum nodosum, imparts thermal stress tolerance in Caenorhabditis elegans. Mar. Drugs 2011, 9, 2256–2282. [Google Scholar]
- Kim, S.K.; Wijesekara, I. Development and biological activities of marine derived bioactive peptides: A review. J. Funct. Foods 2010, 2, 1–9. [Google Scholar]
- Wijesekara, I.; Yoon, N.Y.; Kim, S.K. Phlorotannins from Ecklonia cava (Phaeophyceae): Biological activities and potential health benefits. Biofactors 2010, 36, 408–414. [Google Scholar]
- Allen, V.G.; Pond, K.R.; Saker, K.E.; Fontenot, J.P.; Bagley, C.P.; Ivy, R.L.; Evans, R.R.; Schmidt, R.E.; Fike, J.H.; Zhang, X.; et al. Tasco: Influence of a brown seaweed on antioxidants in forages and livestock—A review. J. Anim. Sci. 2001, 79, E21–E31. [Google Scholar]
- Turner, J.L.; Dritz, S.S.; Higgins, J.J.; Minton, J.E. Effects of Ascophyllum nodosum extract on growth performance and immune function of young pigs challenged with Salmonella Typhimurium. J. Anim. Sci. 2002, 80, 1947–1953. [Google Scholar]
- Dell’Antonio, G.; Quattrini, A.; Dal-Cin, E.; Fugenzi, A.; Ferrero, M.E. Relief of inflammatory pain in rats by local use of the selective P2X7 ATP receptor inhibitor, oxidized ATP. Arthritis Rheum. 2002, 46, 3378–3385. [Google Scholar]
- Vacca, D.D.; Walsh, R.A. The anti-bacterial activity of an extract obtained from Ascophyllum nodosum. J. Am. Pharm. Assoc. 1954, 43, 24–26. [Google Scholar]
- Wijesekaraa, I.; Pangestutia, R.; Kim, S.-K. Biological activities and potential health benefits of sulfated polysaccharides derived from marine algae. Carbohydr. Polym. 2011, 84, 14–21. [Google Scholar]
- Baardseth, E. Synopsis of biological data on knobbed wrack Ascophyllum nodosum (L.) LeJolis. Fish Synop. 1970, 38, 44. [Google Scholar]
- Wood, R.E.; Boat, T.F.; Doershuk, C.F. State of the art: Cystic fibrosis. Am. Rev. Respir. Dis. 1976, 113, 833–878. [Google Scholar]
- Wagner, V.E.; Filiatrault, M.J.; Picardo, K.F.; Iglewski, B.H. Pseudomonas Aeruginosa Virulence and Pathogenesis Issues. In Pseudomonas Genomics and Molecular Biology, 1st; Cornelis, P., Ed.; Caister Academic Press: Norfolk, UK, 2008; pp. 129–158. [Google Scholar]
- Lyczak, J.B.; Cannon, C.L.; Pier, G.B. Establishment of Pseudomonas aeruginosa infection: Lessons from a versatile opportunist. Microbes Infect. 2000, 2, 1051–1060. [Google Scholar]
- Wiener-Kronish, J.P.; Frank, D.; Sawa, T. Mechanisms of Lung Epithelial Cell Injury by Pseudomonas aeruginosa. In Molecular Biology of Acute Lung Injury, 1st; Wong, H.R., Shanley, T.P., Eds.; Kluwer Academic: Norwell, MA, USA, 2001; pp. 149–161. [Google Scholar]
- Stewart-Tull, D.E.S.; Armstrong, A.V. The effect of 1-hydroxylphenazine and pyocyanin from Pseudomonas aeruginosa on mammalian cell respiration. J. Med. Microbiol. 1972, 5, 67–73. [Google Scholar]
- Hassan, H.M.; Fridovich, I. Mechanism of the antibiotic action of pyocyanine. J. Bacteriol. 1980, 141, 156–163. [Google Scholar]
- Hassett, D.J.; Cohen, M.S. Bacterial adaptation to oxidative stress: Implications for pathogenesis and interaction with phagocytic cells. FASEB J. 1989, 3, 2574–2582. [Google Scholar]
- Schoental, R. The nature of the antibacterial agents present in Pseudomonas pyocyanea cultures. Br. J. Exp. Pathol. 1941, 22, 137–147. [Google Scholar]
- Baron, S.S.; Rowe, J.J. Antibiotic action of pyocyanin. Antimicrob. Agents Chemother. 1981, 20, 814–820. [Google Scholar]
- Castricp, A. Hydrogen cyanide, a secondary metabolite of Pseudomonas aeruginosa. Can. J. Microbiol. 1975, 21, 613–618. [Google Scholar]
- Davies, D.G.; Parsek, M.R.; Pearson, J.P.; Iglewski, B.H.; Costerton, J.W.; Greenberg, E.P. The involvement of cell-to-cell signals in the development of a bacterial biofilm. Science 1998, 280, 295–298. [Google Scholar]
- Fuqua, W.C.; Winans, S.C.; Greenberg, E.P. Quorum sensing in bacteria: The LuxR-LuxI family of cell density-responsive transcriptional regulators. J. Bacteriol. 1994, 176, 269–275. [Google Scholar]
- Williams, P.; Camara, M.; Hardman, A.; Swift, S.; Milton, D.; Hope, V.J.; Winzer, K.; Middleton, B.; Pritchard, D.I.; Bycroft, B.W. Quorum sensing and the population dependent control of virulence. Phil. Trans. R. Soc. Lond. Ser. B 2000, 355, 667–680. [Google Scholar]
- Tan, M.W.; Rahme, L.G.; Sternberg, J.; Tompkins, R.G.; Ausubel, F.M. Pseudomonas aeruginosa killing of Caenorhabditis elegans used to identify P. aeruginosa virulence factors. Proc. Natl. Acad. Sci. USA 1999, 96, 2408–2413. [Google Scholar]
- Aballay, A.; Ausubel, F.M. Caenorhabditis elegans as a host for the study of host-pathogen interactions. Curr. Opin. Microbiol. 2002, 5, 97–101. [Google Scholar]
- Tan, M.W.; Ausubel, F.M. Caenorhabditis elegans: A model genetic host to study Pseudomonas aeruginosa pathogenesis. Curr. Opin. Microbiol. 2000, 3, 29–34. [Google Scholar]
- Sifri, C.D.; Begun, J.; Ausubel, F.M. The worm has turned-microbial virulence modeled in Caenorhabditis elegans. Trends Microbiol. 2005, 13, 119–127. [Google Scholar]
- Kim, D.H.; Ausubel, F.M. Evolutionary perspectives on innate immunity from the study of Caenorhabditis elegans. Curr. Opin. Immunol. 2005, 17, 4–10. [Google Scholar]
- Pujol, N.; Zugasti, O.; Wong, D.; Couillault, C.; Kurz, C.L.; Schulenburg, H.; Ewbank, J.J. Anti-fungal innate immunity in C. elegans is enhanced by evolutionary diversification of antimicrobial peptides. PLoS Pathol. 2008, 4, e1000105. [Google Scholar]
- Bach, S.J.; Wangb, Y.; McAllister, T.A. Effect of feeding sun-dried seaweed (Ascophyllum nodosum) on fecal shedding of Escherichia coli O157:H7 by feedlot cattle and on growth performance of lambs. Anim. Feed Sci. Technol. 2008, 142, 17–32. [Google Scholar]
- Braden, K.W.; Blanton, J.R., Jr.; Montgomery, J.L.; van Santen, E.; Allen, V.G.; Miller, M.F. Tasco supplementation: effects on carcass characteristics, sensory attributes, and retail display shelf-life. J. Anim. Sci. 2007, 85, 754–768. [Google Scholar]
- Allen, V.G.; Pond, K.R.; Saker, K.K.; Fontenot, J.P.; Bagley, C.P.; Ivy, R.L.; Evans, R.R.; Schmidt, R.E.; Fike, J.H.; Zhang, X.; et al. Tasco: Influence of a brown seaweed on antioxidants in forages and livestock—A review. J. Anim. Sci. 2001, 79, E21–E31. [Google Scholar]
- Braden, K.W.; Blanton, J.R.; Allen, V.G.; Pond, K.R.; Miller, M.F. Ascophyllum nodosum supplementation: A pre-harvest intervention for reducing Escherichia coli O157:H7 and Salmonella spp. in feedlot steers. J. Food Prot. 2004, 67, 1824–1828. [Google Scholar] [PubMed]
- Evans, E.A.; Kawli, T.; Tan, M.W. Pseudomonas aeruginosa Suppresses Host Immunity by Activating the DAF-2 Insulin-Like Signaling Pathway in Caenorhabditis elegans. PLoS Pathog. 2008, 4, e1000175. [Google Scholar]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar]
- Reilly, P.; O’Doherty, J.; Pierce, K.; Callan, J.; O’Sullivan, J.; Sweeney, T. The effects of seaweed extract inclusion on gut morphology, selected intestinal microbiota, nutrient digestibility, volatile fatty acids concentrations and the immune status of the weaned pig. Animal 2008, 2, 1465–1473. [Google Scholar]
- Wang, J.; Zhang, Q.; Zhang, Z.; Li, Z. Antioxidant activity of sulfated polysaccharide fractions extracted from Laminaria japonica. Int. J. Biol. Macromol. 2008, 42, 127–132. [Google Scholar]
- Lin, Y.C.; Yeh, S.T.; Li, C.C.; Chen, L.L.; Cheng, A.C.; Chen, J.C. An immersion of Gracilaria tenuistipitata extract improves the immunity and survival of white shrimp Litopenaeus vannamei challenged with white spot syndrome virus. Fish Shellfish Immunol. 2011, 31, 1239–1246. [Google Scholar]
- Fujiki, K.; Matsuyama, H.; Yano, T. Effect of hot-water extracts from marine algae on resistance of carp and yellowtail against bacterial infections. Sci. Bull. Fac. Agric. 1992, 47, 137–141. [Google Scholar]
- Fujiki, K.; Matsuyama, H.; Yano, T. Protective effect of sodium alginates against bacterial infection in common carp, Cyprinus carpio L. J. Fish Dis. 1994, 17, 349–355. [Google Scholar] [CrossRef]
- Cheng, W.; Liu, C.H.; Yeh, S.T.; Chen, J.C. The immune stimulatory effect of sodium alginate on the white shrimp Litopenaeus vannamei and its resistance against Vibrio alginolyticus. Fish Shellfish Immunol. 2004, 17, 41–51. [Google Scholar]
- Cheng, W.; Liu, C.H.; Kuo, C.M.; Chen, J.C. Dietary administration of sodium alginate enhances the immune ability of white shrimp Litopenaeus vannamei and its resistance against Vibrio alginolyticus. Fish Shellfish Immunol. 2005, 18, 1–12. [Google Scholar]
- Chotigeat, W.; Tongsupa, S.; Supamataya, K.; Phongdara, A. Effect of fucoidan on disease resistance of black tiger shrimp. Aquaculure 2004, 233, 23–30. [Google Scholar]
- Cumashi, A.; Ushakova, N.A.; Preobrazhenskaya, M.E.; D’Incecco, A.; Piccoli, A.; Totani, L.; Tinari, N.; Morozevich, G.E.; Berman, A.E.; Bilan, M.I.; et al. A comparative study of the anti-inflammatory, anticoagulant, antiangiogenic and anti-adhesive activities of nine different fucoidans from brown seaweeds. Glycobiology 2007, 17, 541–552. [Google Scholar] [PubMed]
- Damonte, E.B.; Matulewicz, M.C.; Cerezo, A.S. Sulfated seaweed polysaccharides as antiviral agents. Curr. Med. Chem. 2004, 11, 2399–2419. [Google Scholar]
- Azevedo, T.C.G.; Bezerra, M.E.; Santos, M.D.; Souza, L.A.; Marques, C.T.; Benevides, N.M.; Leite, E.L. Heparinoids algal and their anticoagulant, hemorrhagic activities and platelet aggregation. Biomed. Pharmacother. 2009, 63, 477–483. [Google Scholar]
- Sulston, J.; Hodgkin, J. The Nematode Caenorhabditis elegans; Wood, W.B., Ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1988; p. 587. [Google Scholar]
- Atlas, R.M.; Parks, L.C. Handbook of Microbiological Media; CRC Press Inc.: London, UK, 1993. [Google Scholar]
- Schaber, J.A.; Carty, N.L.; McDonald, N.A.; Graham, E.D.; Cheluvappa, R.; Griswold, J.A.; Hamood, A.N. Analysis of quorum sensing-deficient clinical isolates of Pseudomonas aeruginosa. J. Med. Microbiol. 2004, 53, 841–853. [Google Scholar]
- Shao, Z.; Zhang, Y.; Powell-Coffman, J.A. Two distinct roles for EGL-9 in the regulation of HIF-1-mediated gene expression in Caenorhabditis elegans. Genetics 2009, 183, 821–829. [Google Scholar]
- Dow, J.M.; Clarke, B.R.; Milligan, D.E.; Tang, J.L.; Daniels, M.J. Extracellular proteases from Xanthomonas campestris pv. campestris, the black rot pathogen. Appl. Environ. Microbiol. 1990, 56, 2994–2998. [Google Scholar] [PubMed]
- Howe, T.R.; Iglewski, B.H. Isolation and characterization of alkaline protease-deficient mutants of Pseudomonas aeruginosa in vitro and in a mouse eye model. Infect. Immun. 1984, 43, 1058–1063. [Google Scholar]
- Rust, L.; Messing, C.R.; Iglewski, B.H. Elastase assays. Methods Enzymol. 1994, 235, 554–562. [Google Scholar]
- Dénervaud, V.; TuQuoc, P.; Blanc, D.; Favre-Bonté, S.; Krishnapillai, V.; Reimmann, C.; Haas, D.; van Delden, C. Characterization of cell-to-cell signaling-decent Pseudomonas aeruginosa strains colonizing intubated patients. J. Clin. Microbiol. 2004, 42, 554–562. [Google Scholar]
- Essar, D.W.; Eberly, L.; Hadero, A.; Crawford, I.P. Identification and characterization of genes for a second anthranilate synthase in Pseudomonas aeruginosa: Interchangeability of the two anthranilate synthases and evolutionary implications. J. Bacteriol. 1990, 172, 884–900. [Google Scholar]
- Miller, R.L.; Higgins, V.J. Association of cyanide with infection of birdfoot trefoil Stemphylium loti. Phytopathology 1970, 60, 104–110. [Google Scholar]
- Sadasivam, S.; Manickam, A. Biochemical Methods, 1st ed; New Age International Ltd. Publisher: New Delhi, India, 1996. [Google Scholar]
- Reeves, M.; Pine, L.; Neilands, J.B.; Bullows, A. Absence of siderophore activity in Legionella sp. grown in iron deficient media. J. Bacteriol. 1983, 154, 324–329. [Google Scholar] [PubMed]
- Toole, O.G.A.; Pratt, L.A.; Watnick, P.I.; Newman, D.K.; Weaver, V.B.; Kolter, R. Genetic approaches to study biofilms. Methods Enzymol. 1999, 310, 91–109. [Google Scholar]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Disc and Dilution Susceptibility Tests for Bacteria Isolated from Animals; Approved Standards, Third ed; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2008.
- COSTAT Statistics Software, version 6.4, CoHort Software: Monterey, CA, USA, 2008.
© 2012 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kandasamy, S.; Khan, W.; Evans, F.; Critchley, A.T.; Prithiviraj, B. Tasco®: A Product of Ascophyllum nodosum Enhances Immune Response of Caenorhabditis elegans Against Pseudomonas aeruginosa Infection. Mar. Drugs 2012, 10, 84-105. https://doi.org/10.3390/md10010084
Kandasamy S, Khan W, Evans F, Critchley AT, Prithiviraj B. Tasco®: A Product of Ascophyllum nodosum Enhances Immune Response of Caenorhabditis elegans Against Pseudomonas aeruginosa Infection. Marine Drugs. 2012; 10(1):84-105. https://doi.org/10.3390/md10010084
Chicago/Turabian StyleKandasamy, Saveetha, Wajahatullah Khan, Franklin Evans, Alan T. Critchley, and Balakrishnan Prithiviraj. 2012. "Tasco®: A Product of Ascophyllum nodosum Enhances Immune Response of Caenorhabditis elegans Against Pseudomonas aeruginosa Infection" Marine Drugs 10, no. 1: 84-105. https://doi.org/10.3390/md10010084
APA StyleKandasamy, S., Khan, W., Evans, F., Critchley, A. T., & Prithiviraj, B. (2012). Tasco®: A Product of Ascophyllum nodosum Enhances Immune Response of Caenorhabditis elegans Against Pseudomonas aeruginosa Infection. Marine Drugs, 10(1), 84-105. https://doi.org/10.3390/md10010084