Chlorella 11-Peptide Inhibits the Production of Macrophage-Induced Adhesion Molecules and Reduces Endothelin-1 Expression and Endothelial Permeability

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
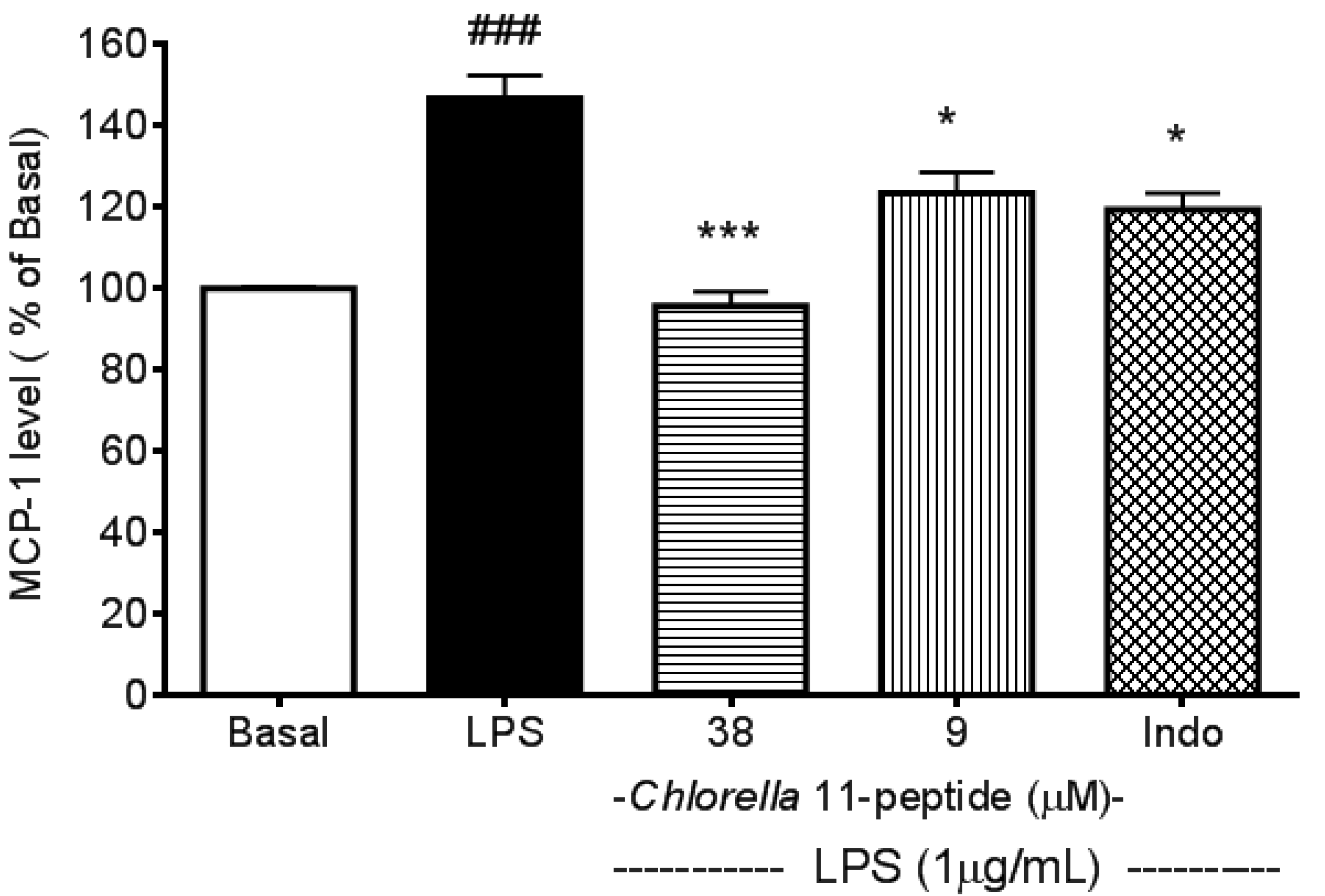
2.1. Inhibitory Effects of Chlorella 11-Peptide on LPS-Induced MCP-1 Production in RAW264.7 Macrophages
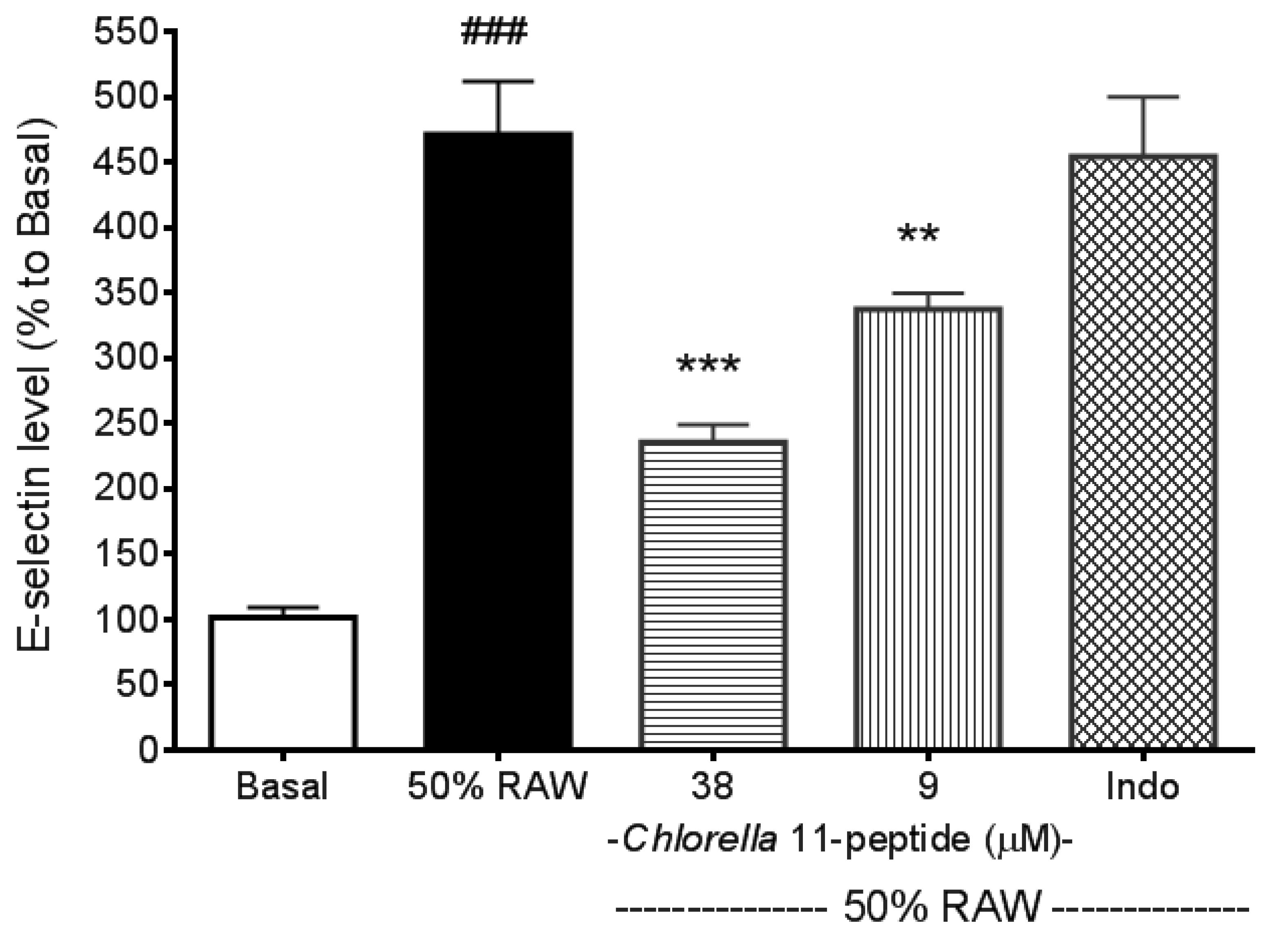
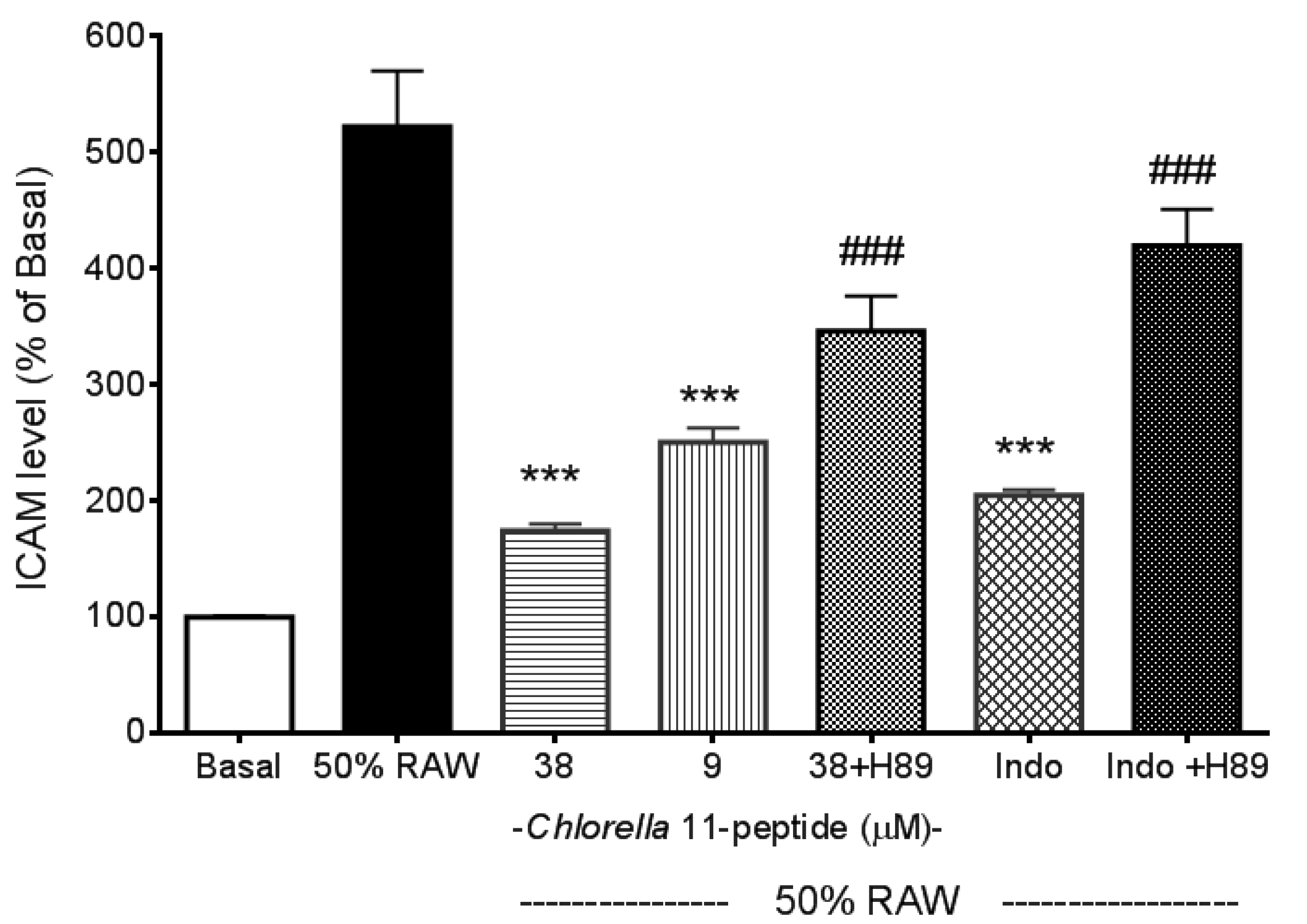
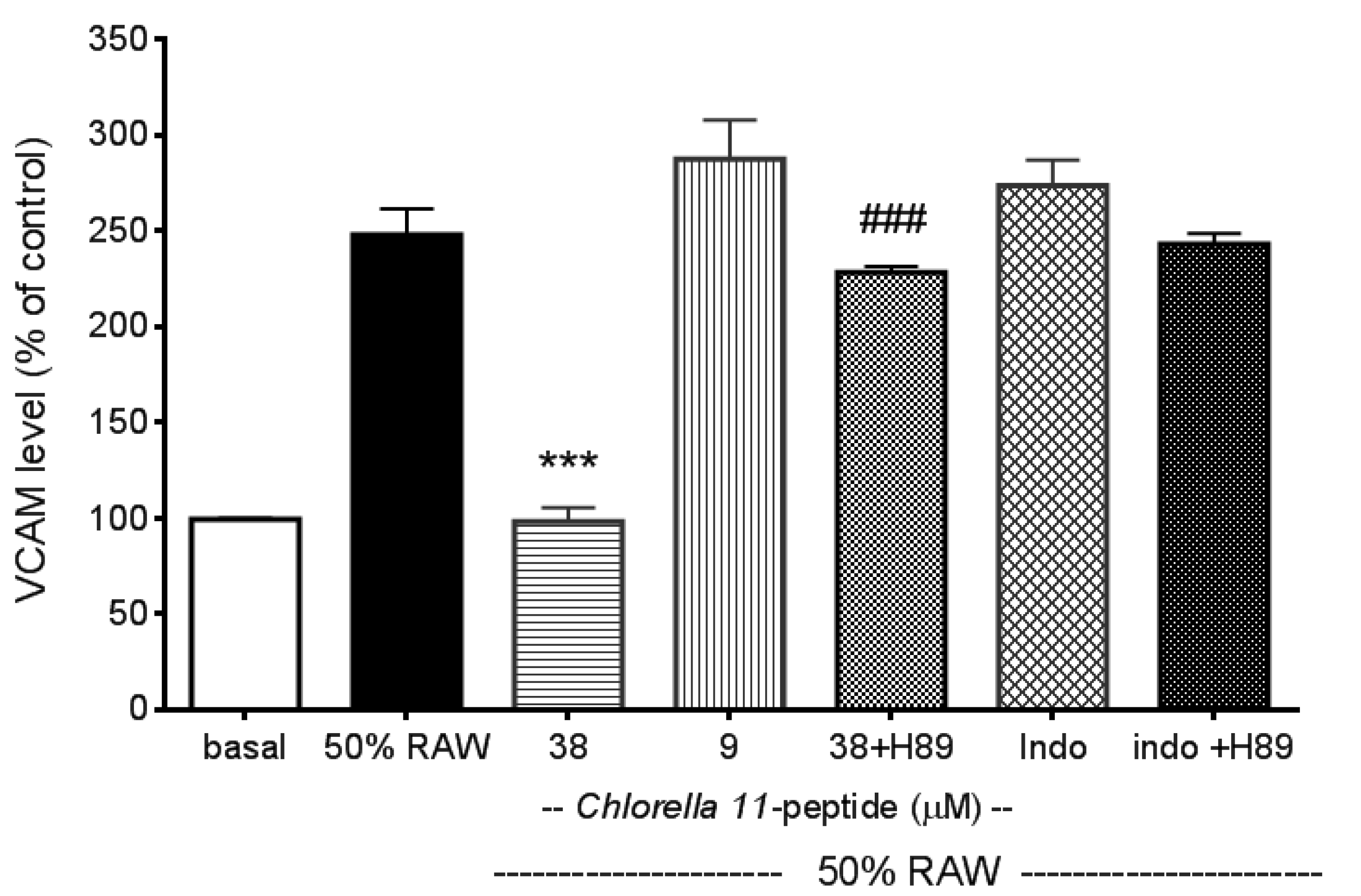
2.2. Inhibitory Effects of Chlorella 11-Peptide on E-Selectin, ICAM-1 and VCAM-1 Production Induced by 50% RAW-Conditioned Medium




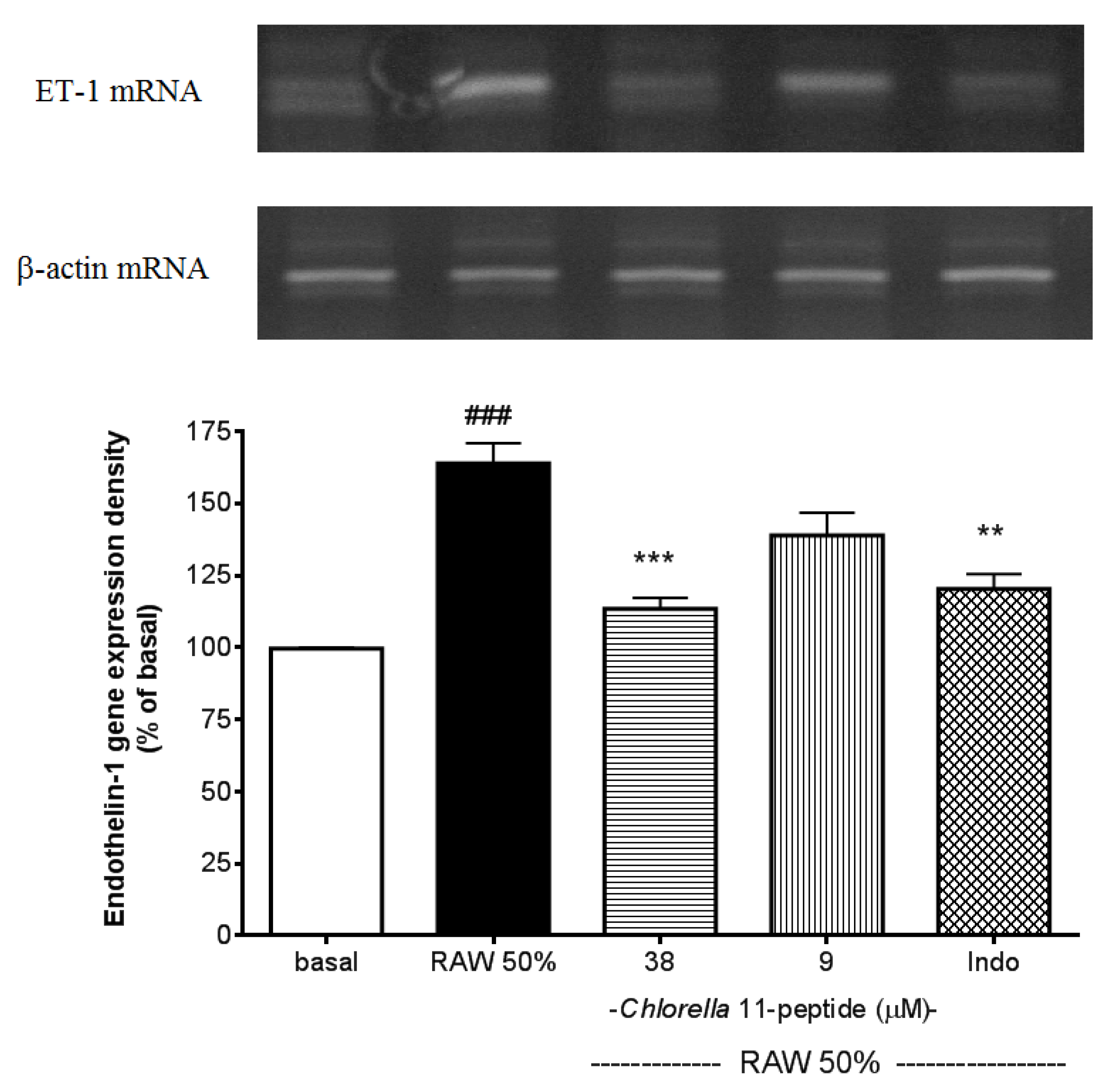
2.3. Inhibitory Effects of Chlorella 11-Peptide on Endothelin-1 Gene Expression

2.4. Inhibitory Effects of Chlorella 11-Peptide on Intercellular Permeability of Endothelia

3. Experimental Section
3.1. Materials
3.2. Chlorella-11 Peptide Preparation
3.3. RAW264.7 Macrophage Culture
3.4. MCP-1 Assay
3.5. Adhesion Molecules—E-Selectin, ICAM-1 & VCAM-1 Level Measurements in SVEC4-10 Endothelial cells
3.6. Endothelin-1 mRNA Analysis by Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
3.7. BSA Transwell Permeability Assay
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Kacimi, R.; Karliner, J.S.; Koudssi, F.; Long, C.S. Expression and regulation of adhesion molecules in cardiac cells by cytokines response to acute hypoxia. Circ. Res. 1998, 82, 576–586. [Google Scholar] [CrossRef]
- Blankenberg, S.; Barbaux, S.; Tiret, L. Adhesion molecules and atherosclerosis. Atherosclerosis 2003, 170, 191–203. [Google Scholar] [CrossRef]
- Ridker, P.M.; Buring, J.E.; Rifai, N. Soluble P-selectin and the risk of future cardiovascular events. Circulation 2001, 103, 491–495. [Google Scholar] [CrossRef]
- Nasuno, A.; Matsubara, T.; Hori, T.; Higuchi, K.; Imai, S.; Nakagawa, I.; Tsuchida, K.; Ozaki, K.; Mezaki, T.; Tanaka, T.; et al. Levels of soluble E-selectin and ICAM-1 in the coronary circulation of patients with stable coronary artery disease: Association with the severity of coronary atherosclerosis. Jpn. Heart. J. 2002, 43, 93–101. [Google Scholar] [CrossRef]
- Blake, G.J.; Ridker, P.M. Inflammatory bio-markers and cardiovascular risk prediction. J. Int. Med. 2002, 252, 283–294. [Google Scholar] [CrossRef]
- Tardif, J.C.; Grégoire, J.; Lavoie, M.A.; L’Allier, P.L. Vascular protectants for the treatment of atherosclerosis. Expert. Rev. Cardiovasc. 2003, 1, 385–392. [Google Scholar] [CrossRef]
- Wilcox, J.N.; Nelken, N.A.; Coughlin, S.R.; Gordon, D.; Schall, T.J. Local expression of inflammatory cytokines in human atherosclerotic plaques. J. Atheroscler. Thromb. 1994, 1, S10–S13. [Google Scholar]
- Gown, A.M.; Tsukada, T.; Ross, R. Human atherosclerosis: II. Immunocytochemical analysis of the cellular composition of human atherosclerotic lesions. Am. J. Pathol. 1986, 125, 191–207. [Google Scholar]
- Deo, R.; Khera, A.; McGuire, D.K.; Murphy, S.A.; Meo Neto, J.P.; Morrow, D.A.; de Lemos, J.A. Association among plasma levels of monocyte chemoattractant protein-1, traditional cardiovascular risk factors, and subclinical atherosclerosis. J. Am. Coll. Cardiol. 2004, 44, 1792–1800. [Google Scholar]
- Sheikine, Y.; Hansson, G.K. Chemokines and atherosclerosis. Ann. Med. 2004, 36, 98–118. [Google Scholar] [CrossRef]
- Hayashidani, S.; Tsutsui, H.; Shiomi, T.; Ikeuchi, M.; Matsusaka, H.; Suematsu, N.; Wen, J.; Egashira, K.; Takeshita, A. Anti-monocyte chemoattractant protein-1 gene therapy attenuates left ventricular remodeling and failure after experimental myocardial infarction. Circulation 2003, 108, 1–7. [Google Scholar] [CrossRef]
- Dewald, O.; Zymek, P.; Winkelmann, K.; Koerting, A.; Ren, G.; Abou-Khamis, T.; Michael, L.H.; Rollins, B.J.; Entman, M.L.; Frangogiannis, N.G. CCL2/Monocyte chemoattractant protein-1 regulates inflammatory responses critical to healing myocardial infarcts. Circ. Res. 2005, 96, 1–9. [Google Scholar]
- Martinovic, I.; Abegunewardene, N.; Seul, M.; Vosseler, M.; Horstick, G.; Buerke, M.; Darius, H.; Lindemann, S. Elevated monocyte chemoattractant protein-1 serum levels in patients at risk for coronary artery disease. Circ. J. 2005, 69, 1484–1489. [Google Scholar] [CrossRef]
- Inoue, S.; Egashira, K.; Ni, W.; Kitamoto, S.; Usui, M.; Otani, K.; Ishibashi, M.; Hiasa, K.; Nishida, K.; Takeshita, A. Anti-monocyte chemoattractant protein-1 gene therapy limits progression and destabilization of established atherosclerosis in apolipoprotein E-knockout mice. Circulation 2002, 106, 2700–2706. [Google Scholar] [CrossRef]
- Little, P.J.; Ivey, M.E.; Osman, N. Endothelin-1 actions on vascular smooth muscle cell functions as a target for the prevention of atherosclerosis. Curr. Vasc. Pharmacol. 2008, 6, 195–203. [Google Scholar] [CrossRef]
- Winkles, J.A.; Alberts, G.F.; Brogi, E.; Libby, P. Endothelin-1 and endothelin receptor mRNA expression in normal and atherosclerotic human arteries. Biochem. Biophys. Res. Commun. 1993, 191, 1081–1088. [Google Scholar] [CrossRef]
- Cherng, J.-Y.; Shih, M.-F. Preventing dyslipidemia by Chlorella pyrenoidosa in rats and hamsters after chronic high fat diet treatment. Life Sci. 2005, 76, 3001–3013. [Google Scholar] [CrossRef]
- Sano, T.; Tanaka, Y. Effects of dried powdered Chlorella vulgaris on experimental atherosclerosis and alimentary hypercholesterolemia in cholesterol-fed rabbit. Artery 1987, 14, 76–84. [Google Scholar]
- Cherng, J.Y.; Liu, C.C.; Shen, C.R.; Lin, H.H.; Shih, M.-F. Beneficial effects of Chlorella-11 peptide on blocking the LPS-induced macrophage activation and alleviating the thermal injury induced inflammation in rats. Int. J. Immunopathol. Pharmacol. 2010, 24, 817–826. [Google Scholar]
- Sheih, I.C.; Fang, T.J.; Wu, T.K.; Lin, P.H. Anticancer and antioxidant activities of the peptide fraction from algae protein waste. J. Agric. Food Chem. 2010, 58, 1202–1207. [Google Scholar] [CrossRef]
- Ross, R. Atherosclerosis is an inflammatory disease. Am. Heart. J. 1999, 138, S419–S420. [Google Scholar] [CrossRef]
- Rus, H.G.; Vlaicu, R.; Niculescu, F. Interleukin-6 and interleukin-8 protein and gene expression in human arterial atherosclerotic wall. Atherosclerosis 1996, 127, 263–271. [Google Scholar] [CrossRef]
- Harama, D.; Koyama, K.; Mukai, M.; Shimokawa, N.; Miyata, M.; Nakamura, Y.; Ohnuma, Y.; Ogawa, H.; Matsuoka, S.; Paton, A.W.; et al. A subcytotoxic dose of subtilase cytotoxin prevents lipopolysaccharide-induced inflammatory responses, depending on its capacity to induce the unfolded protein response. J. Immunol. 2009, 183, 1368–1374. [Google Scholar] [CrossRef]
- Krieglstein, C.F.; Granger, D.N. Adhesion molecules and their role in vascular disease. Am. J. Hypertens. 2001, 14, S44–S54. [Google Scholar] [CrossRef]
- Granger, D.N.; Vowinkel, T.; Petnehazy, T. Modulation of the inflammatory response in cardiovascular disease. Hypertension 2004, 43, 924–931. [Google Scholar] [CrossRef]
- Amberger, A.; Hala, M.; Saurwein-Teissl, M.; Metzler, B.; Grubeck-Loebenstein, B.; Xu, Q.; Wick, G. Suppressive effects of anti-inflammatory agents on human endothelial cell activation and induction of heat shock proteins. Mol. Med. 1999, 5, 117–128. [Google Scholar]
- Fotis, L.; Giannakopoulos, D.; Stamogiannou, L.; Xatzipsalti, M. Intercellular cell adhesion molecule-1 and vascular cell adhesion molecule-1 in children. Do they play a role in the progression of atherosclerosis? Hormones (Athens) 2012, 11, 140–146. [Google Scholar]
- Poston, R.N.; Johnson-Tidey, R.R. Localized adhesion of monocytes to human atherosclerotic plaques demonstrated in vitro: Implications for atherogenesis. Am. J. Pathol. 1996, 149, 73–80. [Google Scholar]
- Ghersa, P.; Hooft van Huijs duijnen, R.; Whelan, J.; Cambet, Y.; Pescini, R.; DeLamarter, J.F. Inhibition of E-selectin gene transcription through a cAMP-dependent protein kinase pathway. J. Biol. Chem. 1994, 269, 29129–29137. [Google Scholar]
- Balyasnikova, I.V.; Pelligrino, D.A.; Greenwood, J.; Adamson, P.; Dragon, S.; Raza, H.; Galea, E. Cyclic adenosine monophosphate regulates the expression of the intercellular adhesion molecule and the inducible nitric oxide synthase in brain endothelial cells. J. Cereb. Blood. Flow. Metab. 2000, 20, 688–699. [Google Scholar]
- Ono, H.; Ichiki, T.; Ohtsubo, H.; Fukuyama, K.; Imayama, I.; Iino, N.; Masuda, S.; Hashiguchi, Y.; Takeshita, A.; Sunagawa, K. cAMP-Response element-binding protein mediates tumor necrosis factor-alpha-induced vascular cell adhesion molecule-1 expression in endothelial cells. Hypertens. Res. 2006, 29, 39–47. [Google Scholar] [CrossRef]
- Goueli, S.A.; Ahmed, K. Indomethacin and inhibition of protein kinase reactions. Nature 1980, 287, 171–172. [Google Scholar] [CrossRef]
- Babaei, S.; Picard, P.; Ravandi, A.; Monge, J.C.; Lee, T.C.; Cernacek, P.; Stewart, D.J. Blockade of endothelin receptors markedly reduces atherosclerosis in LDL receptor deficient mice: Role of endothelin in macrophage foam cell formation. Cardiovasc. Res. 2000, 48, 158–167. [Google Scholar] [CrossRef]
- Dejana, E.; Orsenigo, F.; Lampugnani, M.G. The role of adherens junctions and VE-cadherin in the control of vascular permeability. J. Cell Sci. 2008, 121, 2115–2122. [Google Scholar] [CrossRef]
- Mehta, D.; Malik, A.B. Signaling mechanisms regulating endothelial permeability. Physiol. Rev. 2006, 86, 279–367. [Google Scholar] [CrossRef]
- Sun, C.; Wu, M.H.; Yuan, S.Y. Nonmuscle myosin light-chain kinase deficiency attenuates atherosclerosis in apolipoprotein E-deficient mice via reduced endothelial barrier dysfunction and monocyte migration. Circulation 2011, 124, 48–57. [Google Scholar] [CrossRef]
- O’Connell, K.A.; Edidin, M. A mouse lymphoid endothelial cell line immortalized by simian virus 40 binds lymphocytes and retains functional. J. Immunol. 1990, 144, 521–525. [Google Scholar]
- Bian, C.; Wu, Y.; Chen, P. Telmisartan increases the permeability of endothelial cells through zonula occludens-1. Biol. Pharm. Bull. 2009, 32, 416–420. [Google Scholar] [CrossRef]
Supplementary Files
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Shih, M.F.; Chen, L.C.; Cherng, J.Y. Chlorella 11-Peptide Inhibits the Production of Macrophage-Induced Adhesion Molecules and Reduces Endothelin-1 Expression and Endothelial Permeability. Mar. Drugs 2013, 11, 3861-3874. https://doi.org/10.3390/md11103861
Shih MF, Chen LC, Cherng JY. Chlorella 11-Peptide Inhibits the Production of Macrophage-Induced Adhesion Molecules and Reduces Endothelin-1 Expression and Endothelial Permeability. Marine Drugs. 2013; 11(10):3861-3874. https://doi.org/10.3390/md11103861
Chicago/Turabian StyleShih, Mei Fen, Lih Chi Chen, and Jong Yuh Cherng. 2013. "Chlorella 11-Peptide Inhibits the Production of Macrophage-Induced Adhesion Molecules and Reduces Endothelin-1 Expression and Endothelial Permeability" Marine Drugs 11, no. 10: 3861-3874. https://doi.org/10.3390/md11103861
APA StyleShih, M. F., Chen, L. C., & Cherng, J. Y. (2013). Chlorella 11-Peptide Inhibits the Production of Macrophage-Induced Adhesion Molecules and Reduces Endothelin-1 Expression and Endothelial Permeability. Marine Drugs, 11(10), 3861-3874. https://doi.org/10.3390/md11103861