Diamond Squid (Thysanoteuthis rhombus)-Derived Chondroitin Sulfate Stimulates Bone Healing within a Rat Calvarial Defect


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
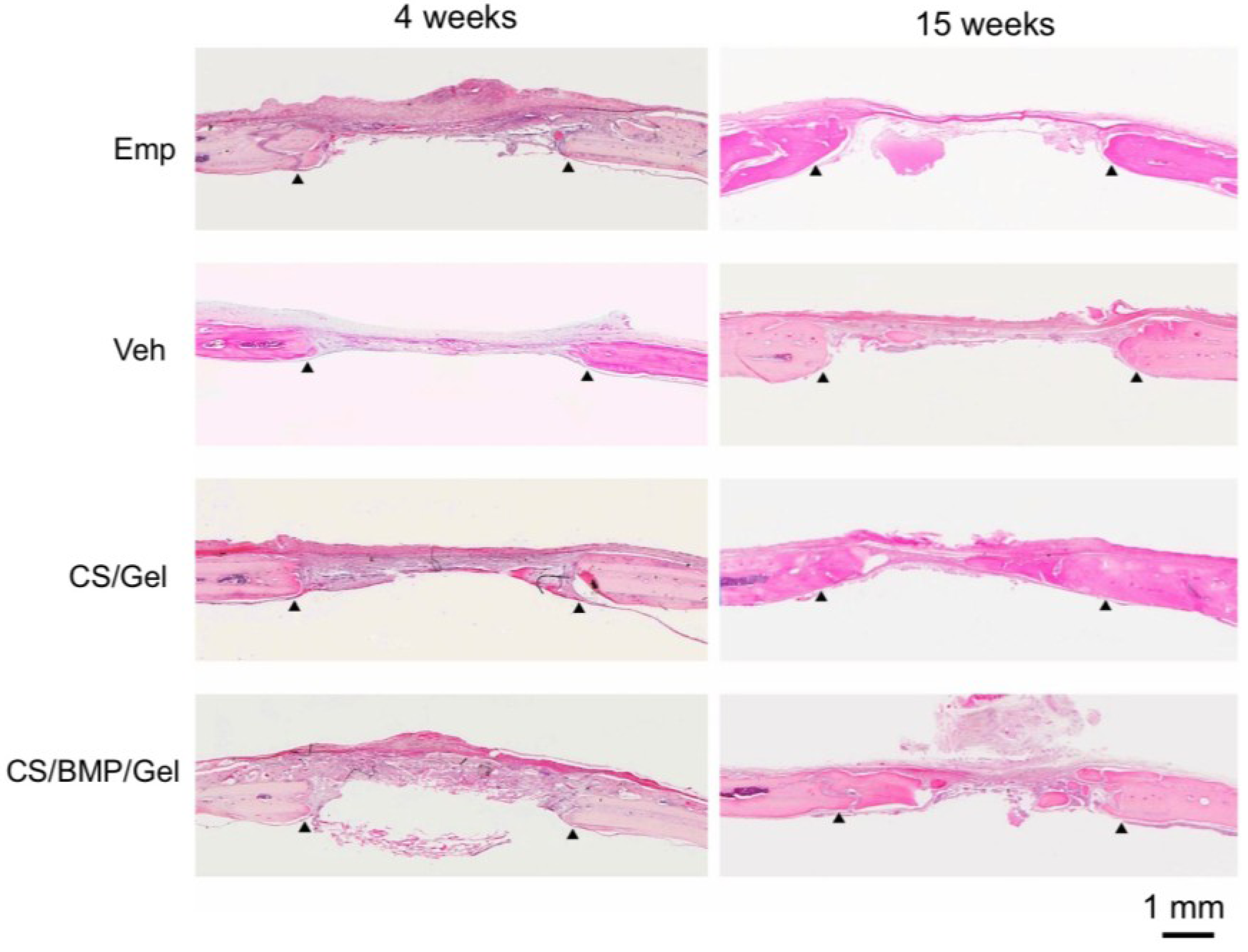
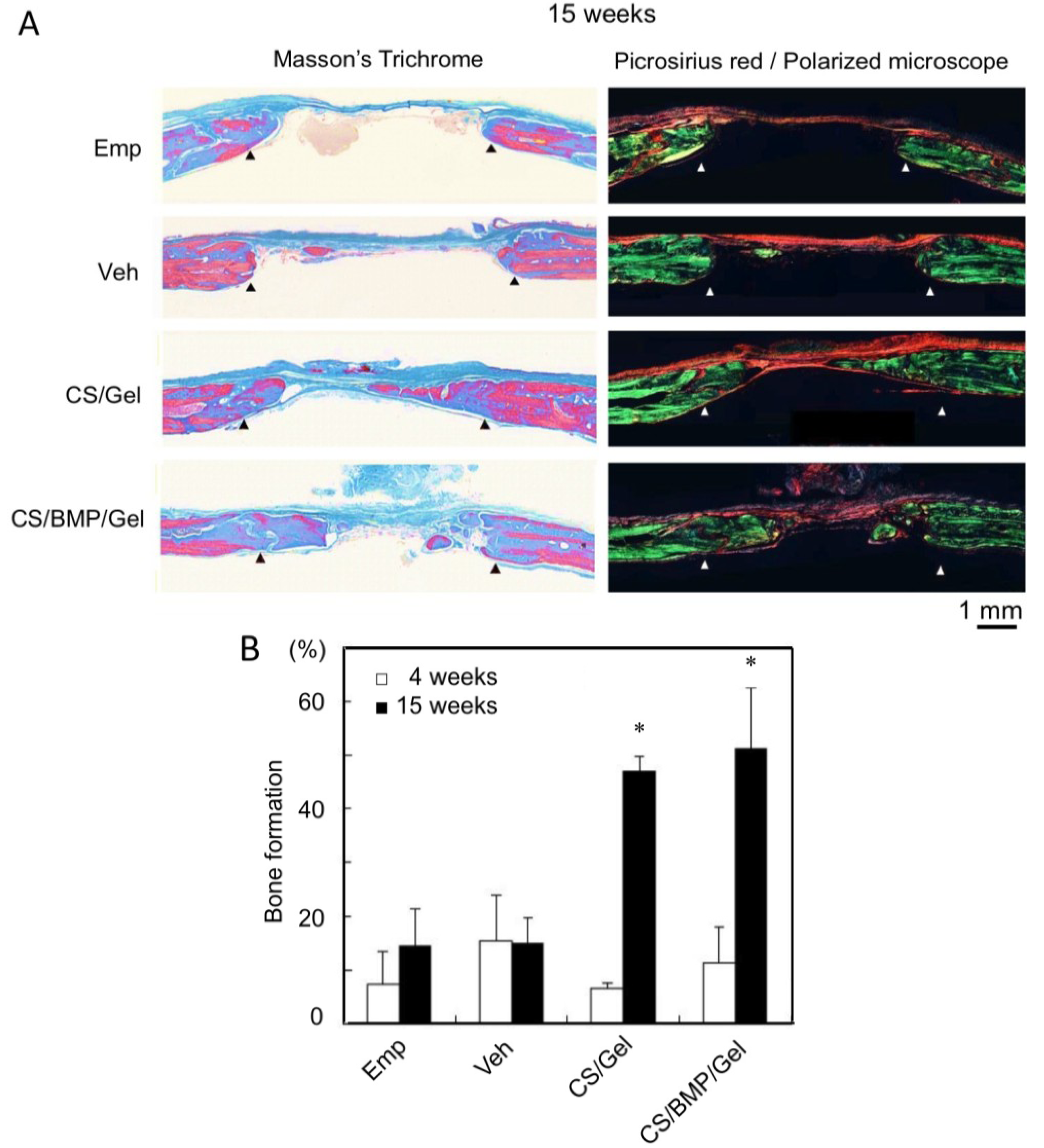
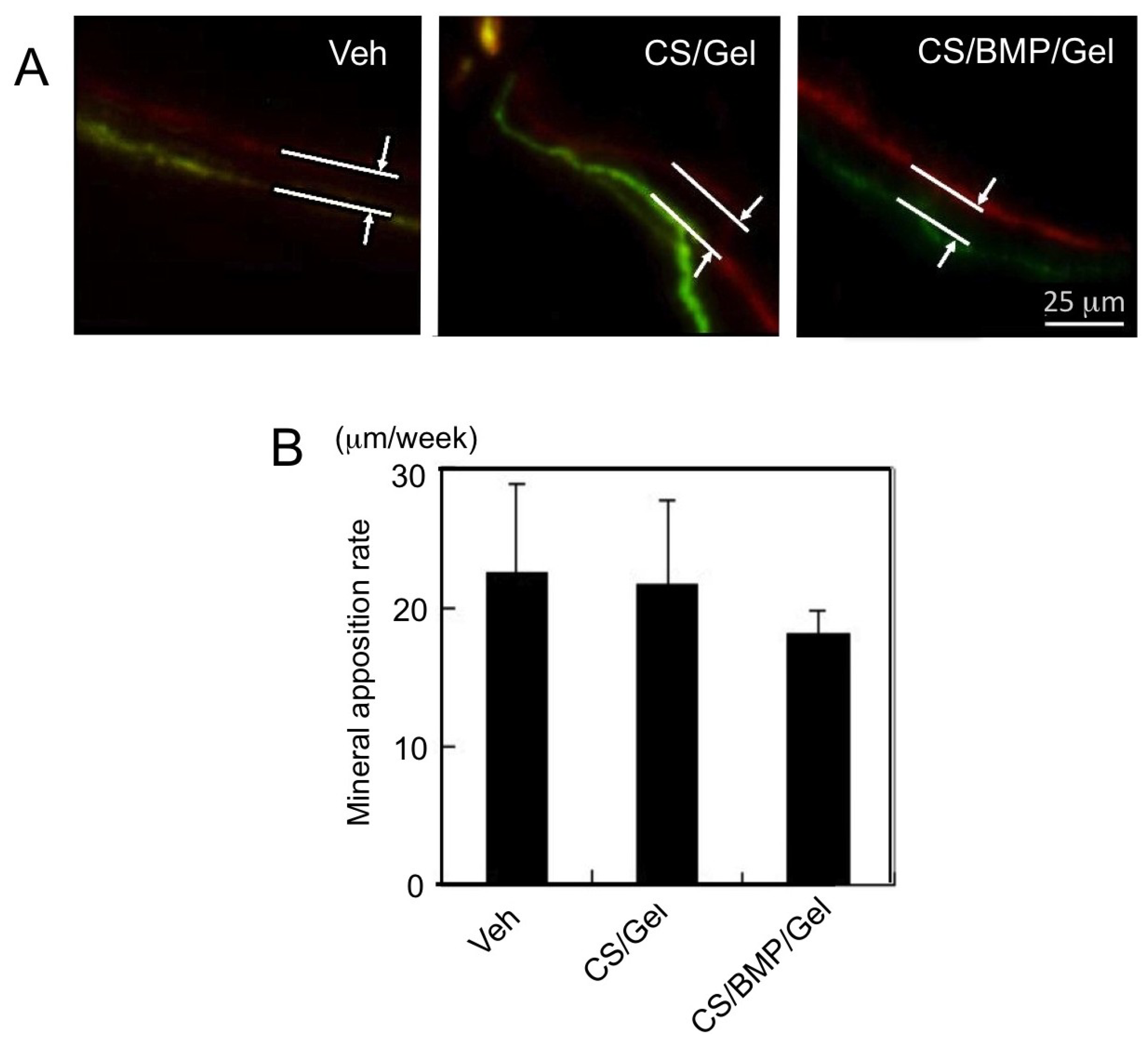
2. Results




3. Discussion
4. Experimental Section
4.1. Experimental Design and Surgical Procedure
4.2. Mineralization Label by Fluorochrome
4.3. Histological and Histomorphometrical Analyses
4.4. Statistical Analysis
5. Conclusions
Acknowledgements
Conflicts of Interest
References
- Bellows, C.G.; Aubin, J.E. Determination of numbers of osteoprogenitors present in isolated fetal rat calvaria cells in vitro. Dev. Biol. 1989, 133, 8–13. [Google Scholar] [CrossRef]
- Stein, G.S.; Lian, J.B. Molecular mechanisms mediating proliferation/differentiation interrelationships during progressive development of the osteoblast phenotype. Endocr. Rev. 1993, 14, 424–442. [Google Scholar]
- Hoshi, K.; Ejiri, S.; Ozawa, H. Localizational alterations of calcium, phosphorus, and calcification-related organics such as proteoglycans and alkaline phosphatase during bone calcification. J. Bone Miner. Res. 2001, 16, 289–298. [Google Scholar] [CrossRef]
- Boskey, A.L.; Spevak, L.; Doty, S.B.; Rosenberg, L. Effects of bone CS-proteoglycans, DS-decorin, and DS-biglycan on hydroxyapatite formation in a gelatin gel. Calcif. Tissue Int. 1997, 61, 298–305. [Google Scholar] [CrossRef]
- Silverman, L.; Boskey, A.L. Diffusion systems for evaluation of biomineralization. Calcif. Tissue Int. 2004, 75, 494–501. [Google Scholar] [CrossRef]
- Ameye, L.; Young, M.F. Mice deficient in small leucine-rich proteoglycans: Novel in vivo models for osteoporosis, osteoarthritis, Ehlers-Danlos syndrome, muscular dystrophy, and corneal diseases. Glycobiology 2002, 12, 107R–116R. [Google Scholar] [CrossRef]
- Wadhwa, S.; Bi, Y.; Ortiz, A.T.; Embree, M.C.; Kilts, T.; Iozzo, R.; Opperman, L.A.; Young, M.F. Impaired posterior frontal sutural fusion in the biglycan/decorin double deficient mice. Bone 2007, 40, 861–866. [Google Scholar] [CrossRef]
- Iozzo, R.V.; Murdoch, A.D. Proteoglycans of the extracellular environment: Clues from the gene and protein side offer novel perspectives in molecular diversity and function. FASEB J. 1996, 10, 598–614. [Google Scholar]
- Seldin, D.C.; Seno, N.; Austen, K.F.; Stevens, R.L. Analysis of polysulfated chondroitin disaccharides by high-performance liquid chromatography. Anal. Biochem. 1984, 141, 291–300. [Google Scholar] [CrossRef]
- Miyazaki, T.; Miyauchi, S.; Tawada, A.; Anada, T.; Matsuzaka, S.; Suzuki, O. Oversulfated chondroitin sulfate-E binds to BMP-4 and enhances osteoblast differentiation. J. Cell Physiol. 2008, 217, 769–777. [Google Scholar] [CrossRef]
- Miyazaki, T.; Miyauchi, S.; Tawada, A.; Anada, T.; Suzuki, O. Effect of chondroitin sulfate-E on the osteoclastic differentiation of RAW264 cells. Dent. Mater. J. 2010, 29, 403–410. [Google Scholar] [CrossRef]
- Stadlinger, B.; Pilling, E.; Mai, R.; Bierbaum, S.; Berhardt, R.; Scharnweber, D.; Eckelt, U. Effect of biological implant surface coatings on boneformation, applying collagen, proteoglycans, glycosaminoglycans and growth factors. J. Mater. Sci. Mater. Med. 2008, 19, 1043–1049. [Google Scholar] [CrossRef]
- Urist, M.R.; Nilsson, O.; Rasmussen, J.; Hirota, W.; Lovell, T.; Schmalzreid, T.; Finerman, G.A. Bone regeneration under the influence of bone morphogenetic protein (BMP) beta tricalcium phosphate (TCP) composite in skull trephine defects in dogs. Clin. Orthop. Relat. Res. 1987, 214, 295–304. [Google Scholar]
- Lindholm, T.C.; Londholm, T.S.; Alitalo, I.; Urist, M.R. Bovine bone morphogenetic protein (bBMP) induced repair of skull trephine defects in sheep. Clin. Orthop. Relat. Res. 1988, 227, 265–268. [Google Scholar]
- Sawyer, A.A.; Song, S.J.; Susanto, E.; Chuan, P.; Lam, C.X.; Woodruff, M.A.; Hutmacher, D.W.; Cool, S.M. The stimulation of healing within a rat calvarial defect by mPCL-TCP/collagen scaffolds loaded with rhBMP-2. Biomaterials 2009, 30, 2479–2488. [Google Scholar] [CrossRef]
- Lu, H.; Kawazoe, N.; Kitajima, T.; Myokan, Y.; Tomita, M.; Umezawa, A.; Chen, G.; Ito, Y. Spatial immobilization of bone morphogenetic protein-4 in a collagen-PLGA hybrid scaffold for enhanced osteoinductivity. Biomaterials 2012, 33, 6014–6146. [Google Scholar]
- Lee, Y.J.; Lee, J.H.; Cho, H.J.; Kim, H.K.; Yoon, T.R.; Shin, H. Electrospun fibers immobilized with bone forming peptide-1 derived from BMP-7 for guided bone regeneration. Biomaterials 2013, 34, 5059–5069. [Google Scholar] [CrossRef]
- Ling, L.; Dombowski, C.; Foon, K.M.; Haupt, L.M.; Stein, G.S.; Nurcombe, V.; van Wijnen, A.J.; Simon, S.M. Synergism between Wnt3a and heparin enhance osteogenesis via a phosphoinositide 3-kinase/Akt/RUNX2 pathway. J. Biol. Chem. 2010, 285, 26233–26244. [Google Scholar] [CrossRef]
- Marie, P.J. Fibroblast growth factor signaling controlling osteoblast differentiation. Gene 2003, 316, 23–32. [Google Scholar] [CrossRef]
- Zhang, X.; Ibrahimi, O.A.; Olsen, S.K.; Umemori, H.; Mohammado, M. Receptor specificity of the fibroblast growth factor family: The complete mammalian FGF family. J. Biol. Chem. 2006, 281, 15694–15700. [Google Scholar] [CrossRef]
- Zhao, B.; Katagiri, T.; Toyoda, H.; Takada, T.; Yanai, T.; Fukuda, T.; Chung, U.I.; Koike, T.; Takaoka, K.; Kamijo, R. Heparin potentiates the in vivo ectopic bone formation induced by bone morphogenetic protein-2. J. Biol. Chem. 2006, 281, 23246–23253. [Google Scholar] [CrossRef]
- Tamura, J.; Arima, K.; Imazu, A.; Tsutsumishita, N.; Fujita, H.; Yamane, M.; Matsumi, Y. Sulfation patterns and the amounts of chondroitin sulfate in the diamond squid, Thysanoteuthis rhombus. Biosci. Biotechnol. Biochem. 2009, 73, 1387–1391. [Google Scholar] [CrossRef]
- Junqueira, L.C.; Bignolas, G.; Brentani, R.R. Picrosirius staining plus polarization microscopy, a specific method for collagen detection in tissue sections. Histochem. J. 1979, 11, 447–455. [Google Scholar] [CrossRef]
- Whittaker, P.; Kloner, R.A.; Boughner, D.R.; Pickering, J.G. Quantitative assessment of myocardial collagen with picrosirius red staining and circularly polarized light. Basic. Res. Cardiol. 1994, 89, 397–410. [Google Scholar] [CrossRef]
- Choi, Y.J.; Lee, J.Y.; Park, J.H.; Park, J.B.; Suh, J.S.; Choi, Y.S.; Lee, S.J.; Chung, C.P.; Park, Y.J. The identification of a heparin binding domain peptide from bone morphogenetic protein-4 and its role on osteogenesis. Biomaterials 2010, 31, 7226–7238. [Google Scholar] [CrossRef]
- Çakır-Özkan, N.; Eyibilen, A.; Ozkan, F.; Gulbahar, M.Y.; Kabak, Y.B. Immunohistochemical analysis of reconstructed sheep mandibles: transport distraction osteogenesis versus autogenous bone grafting. J. Oral Maxillofac. Surg. 2011, 69, 1248–1254. [Google Scholar] [CrossRef]
- Wang, Z.; Clark, C.C.; Brighton, C.T. Up-regulation of bone morphogenetic proteins in cultured murine bone cells with use of specific electric fields. J. Bone Joint Surg. Am. 2006, 88, 1053–1065. [Google Scholar] [CrossRef]
- Suzawa, M.; Takeuchi, Y.; Fukumoto, S.; Kato, S.; Ueno, N.; Miyazono, K.; Matsumoto, T.; Fujita, T. Extracellular matrix-associated bone morphogenetic proteins are essential for differentiation of murine osteoblastic cells in vitro. Endocrinology 1999, 140, 2125–2133. [Google Scholar] [CrossRef]
- Stadlinger, B.; Pilling, E.; Huhle, M.; Mai, R.; Bierbaum, S.; Scharnweber, D.; Kuhlisch, E.; Loukoa, R.; Eckelt, U. Evaluation of osseointeguration of dental implants coated with collagen, chondroitin sulphate and BMP-4: An animal study. Int. J. Oral Maxillofac. Surg. 2008, 37, 54–59. [Google Scholar] [CrossRef]
- Ueno, T.; Kagawa, T.; Kanou, M.; Fujii, T.; Fukunaga, J.; Mizukawa, N.; Sugahara, T.; Yamamoto, T. Immunohistochemical observations of cellular differentiation and proliferation in endochondral bone formation from grafted periosteum: expression and localization of BMP-2 and -4 in the grafted periosteum. J. Craniomaxillofac. Surg. 2003, 31, 356–361. [Google Scholar]
- Sojo, K.; Sawaki, Y.; Hattori, H.; Mizutani, H.; Ueda, M. Immunohistochemical study of vascular endothelial growth factor (VEGF) and bone morphogenetic protein-2, -4 (BMP-2, -4) on lengthened rat femurs. J. Craniomaxillofac. Surg. 2005, 33, 238–245. [Google Scholar] [CrossRef]
- McCullough, K.A.; Waits, C.A.; Garimella, R.; Tague, S.E.; Sipe, J.B.; Anderson, H.C. Immunohistochemical localization of bone morphogenetic proteins (BMPs) 2, 4, 6, and 7 during induced heterotopic bone formation. J. Orthop. Res. 2007, 25, 465–472. [Google Scholar] [CrossRef]
- Takeuchi, H.; Nagayama, M.; Imaizumi, Y.; Tsukahara, T.; Nakazawa, J.; Kusaka, Y.; Ohtomo, K. Immunohistochemical analysis of osteoconductivity of beta-tricalcium phosphate and carbonate apatite applied in femoral and parietal bone defects in rats. Dent. Mater. J. 2009, 28, 595–601. [Google Scholar] [CrossRef]
- Zhang, L. Glycosaminoglycan (GAG) biosynthesis and GAG-binding proteins. Prog. Mol. Biol. Transl. Sci. 2010, 93, 1–17. [Google Scholar] [CrossRef]
- Guimond, S.; Maccarana, M.; Olwin, B.B.; Lindahl, U.; Rapraeger, A.C. Activating and inhibitory heparin sequences for FGF-2 (basic FGF). Distinct requirements for FGF-1, FGF-2, and FGF-4. J. Biol. Chem. 1993, 268, 23906–23914. [Google Scholar]
- Ishihara, M.; Shaklee, P.N.; Yang, Z.; Liang, W.; Wei, Z.; Stack, R.J.; Holme, K. Structural features in heparin which modulate specific biological activities mediated by basic fibroblast growth factor. Glycobiology 1994, 4, 451–458. [Google Scholar] [CrossRef]
- Jackson, R.A.; Murali, S.; van Wijnen, A.J.; Stein, G.S.; Nurcombe, V.; Cool, S.M. Heparan sulfate regulates the anabolic activity of MC3T3-E1 preosteoblast cells by induction of Runx2. J. Cell. Physiol. 2007, 210, 38–50. [Google Scholar] [CrossRef]
- Jiao, X.; Billings, P.C.; O’Connell, M.P.; Kaplan, F.S.; Shore, E.M.; Glaser, D.L. Heparan sulfate proteoglycans (HSPGs) modulate BMP2 osteogenic bioactivity in C2C12 cells. J. Biol. Chem. 2007, 282, 1080–1086. [Google Scholar]
- Kram, V.; Zcharia, E.; Yacoby-Zeevi, O.; Metzger, S.; Chajek-Shaul, T.; Gabet, Y.; Muller, R.; Vlodavsky, I.; Bab, I. Heparanase is expressed in osteoblastic cells and stimulates bone formation and bone mass. J. Cell. Physiol. 2006, 207, 784–792. [Google Scholar] [CrossRef]
- Chen, X.D.; Fisher, L.W.; Robey, P.G.; Young, M.F. The small leucine-rich proteoglycan biglycan modulates BMP-4-induced osteoblast differentiation. FASEB J. 2004, 18, 948–958. [Google Scholar] [CrossRef]
- Parisuthiman, D.; Mochida, Y.; Duarte, W.R.; Yamauchi, M. Biglycan modulates osteoblast differentiation and matrix mineralization. J. Bone Miner. Res. 2005, 20, 1878–1886. [Google Scholar] [CrossRef]
- Mochida, Y.; Parisuthiman, D.; Pornprasertsuk-Damrongsri, S.; Atsawasuwan, P.; Sricholpech, M.; Boskey, A.L.; Yamauchi, M. Decorin modulates collagen matrix assembly and mineralization. Matrix Biol. 2009, 28, 44–52. [Google Scholar] [CrossRef]
- Mochida, Y.; Parisuthiman, D.; Yamauchi, M. Biglycan is a positive modulator of BMP-2 induced osteoblast differentiation. Adv. Exp. Med Biol. 2006, 585, 101–113. [Google Scholar]
- Office of Science and Health Reports, Revised; DHEW Publication No. (NIH) 85-23; DDR/NIH: Bethesda, MD, USA, 1985.
- Takahashi, Y.; Yamamoto, M.; Tabata, Y. Osteogenic differentiation of mesenchymal stem cells in biodegradable sponges composed of gelatin and β-tricalcium phosphate. Biomaterials 2005, 26, 3587–3596. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hosaka, Y.Z.; Iwai, Y.; Tamura, J.-i.; Uehara, M. Diamond Squid (Thysanoteuthis rhombus)-Derived Chondroitin Sulfate Stimulates Bone Healing within a Rat Calvarial Defect. Mar. Drugs 2013, 11, 5024-5035. https://doi.org/10.3390/md11125024
Hosaka YZ, Iwai Y, Tamura J-i, Uehara M. Diamond Squid (Thysanoteuthis rhombus)-Derived Chondroitin Sulfate Stimulates Bone Healing within a Rat Calvarial Defect. Marine Drugs. 2013; 11(12):5024-5035. https://doi.org/10.3390/md11125024
Chicago/Turabian StyleHosaka, Yoshinao Z., Yuji Iwai, Jun-ichi Tamura, and Masato Uehara. 2013. "Diamond Squid (Thysanoteuthis rhombus)-Derived Chondroitin Sulfate Stimulates Bone Healing within a Rat Calvarial Defect" Marine Drugs 11, no. 12: 5024-5035. https://doi.org/10.3390/md11125024
APA StyleHosaka, Y. Z., Iwai, Y., Tamura, J. -i., & Uehara, M. (2013). Diamond Squid (Thysanoteuthis rhombus)-Derived Chondroitin Sulfate Stimulates Bone Healing within a Rat Calvarial Defect. Marine Drugs, 11(12), 5024-5035. https://doi.org/10.3390/md11125024