Identification of Four New agr Quorum Sensing-Interfering Cyclodepsipeptides from a Marine Photobacterium

 ,
,
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Isolation and Structure Elucidation of Ngercheumicins F–I


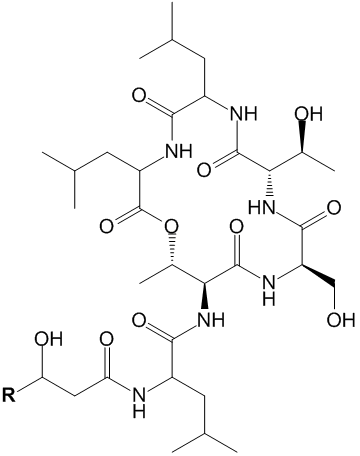{kind=link}
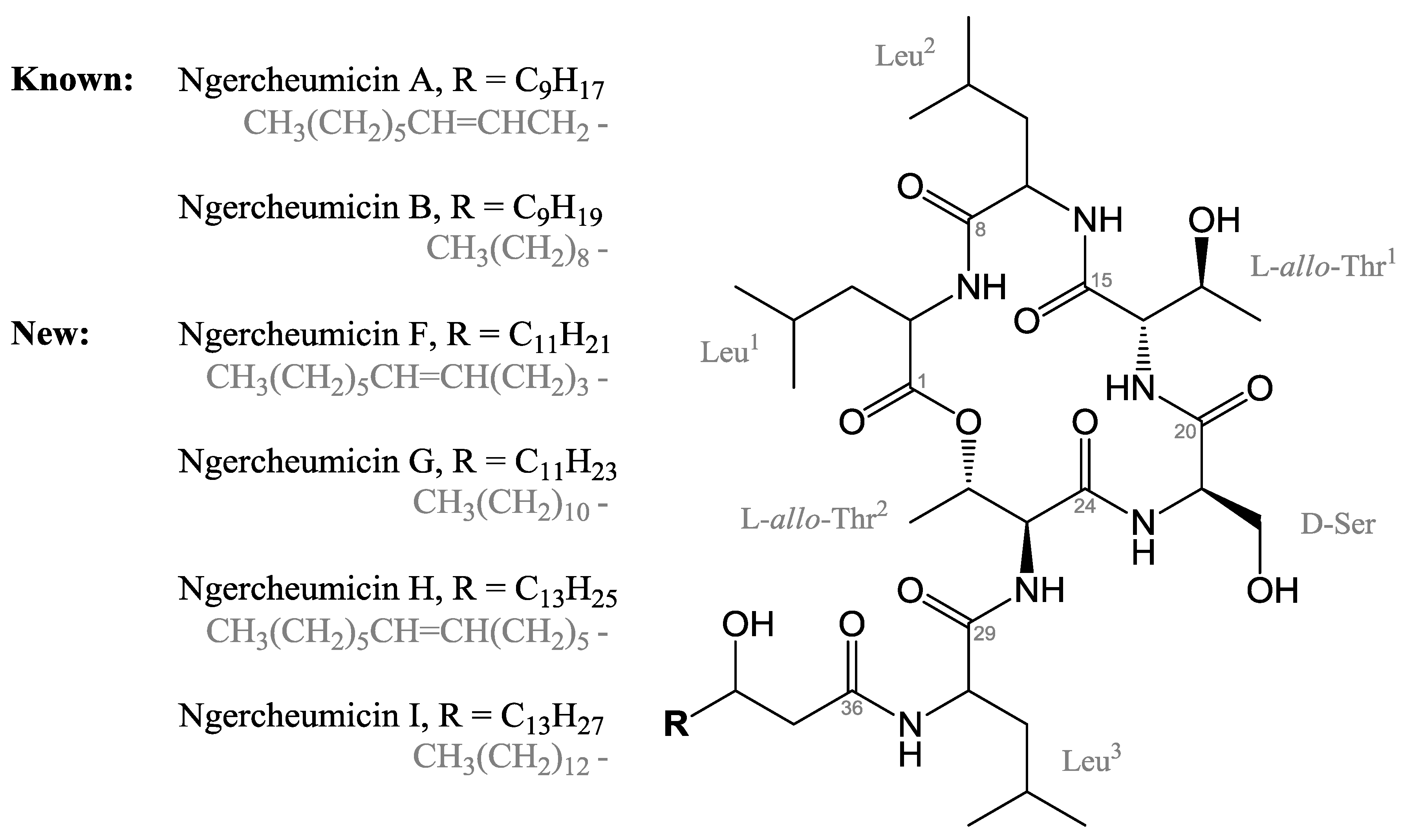{kind=link}
{kind=link}
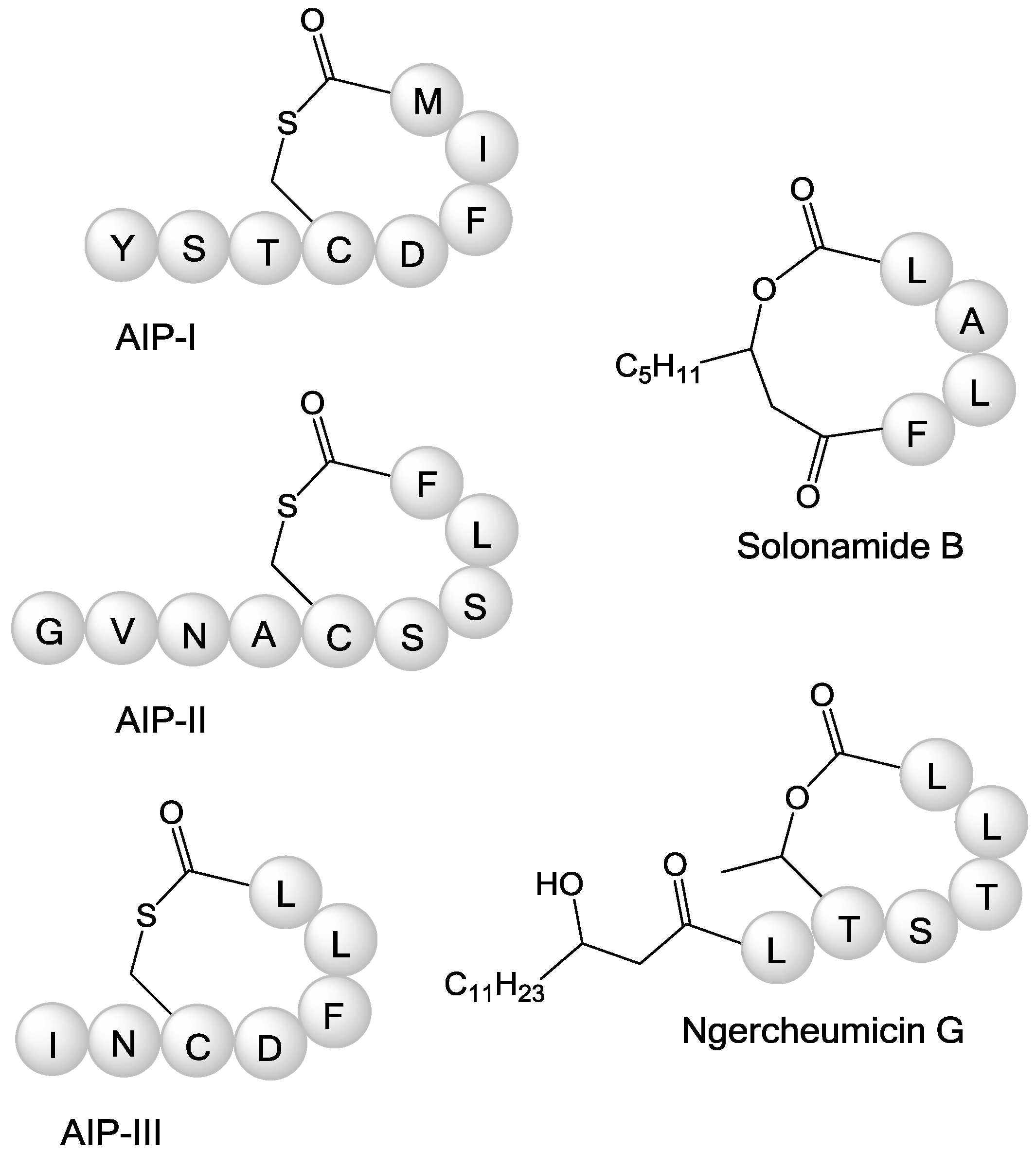{kind=link}
| Ngercheumicin F | Ngercheumicin G | Ngercheumicin H | Ngercheumicin I | |||||
|---|---|---|---|---|---|---|---|---|
| Position, Type | δH (ppm) | δC (ppm) | δH (ppm) | δC (ppm) | δH (ppm) | δC (ppm) | δH (ppm) | δC (ppm) |
| Leu1 | ||||||||
| 1—CO | − | 170.8 | − | 170.8 | − | 170.8 | − | 170.8 |
| 2—CHα | 4.37 | 51.1 | 4.37 | 51.1 | 4.37 | 51.1 | 4.37 | 51.1 |
| 3—CHβ | 1.62, 1.55 | 40.4 | 1.61, 1.55 | 40.4 | 1.62, 1.56 | 40.4 | 1.61, 1.56 | 40.4 |
| 4—CHγ | 1.49 | 24.5 | 1.49 | 24.5 | 1.50 | 24.5 | 1.49 | 24.5 |
| 5—CHδ,1 | 0.88 | 22.3 | 0.88 | 22.3 | 0.89 | 22.3 | 0.88 | 22.3 |
| 6—CHδ,2 | 0.85 | 22.5 | 0.85 | 22.5 | 0.85 | 22.5 | 0.85 | 22.5 |
| 7—NH | 7.73 | − | 7.73 | − | 7.73 | − | 7.73 | − |
| Leu2 | ||||||||
| 8—CO | − | 171.0 | − | 171.0 | − | 171.0 | − | 171.0 |
| 9—CHα | 4.26 | 50.4 | 4.26 | 50.4 | 4.25 | 50.4 | 4.26 | 50.4 |
| 10—CHβ | 1.49 | 38.8 | 1.49 | 38.8 | 1.49 | 38.8 | 1.49 | 38.8 |
| 11—CHγ | 1.53 | 24.2 | 1.53 | 24.2 | 1.54 | 24.1 | 1.54 | 24.1 |
| 12—CHδ,1 | 0.87 | 22.9 | 0.87 | 22.9 | 0.87 | 22.9 | 0.87 | 22.9 |
| 13—CHδ,2 | 0.80 | 21.4 | 0.80 | 21.4 | 0.80 | 21.4 | 0.80 | 21.4 |
| 14—NH | 8.12 | − | 8.11 | − | 8.11 | − | 8.11 | − |
| l-allo-Thr1 | ||||||||
| 15—CO | − | 170.7 | − | 170.7 | − | 170.7 | − | 170.7 |
| 16—CHα | 3.86 | 59.9 | 3.86 | 59.9 | 3.86 | 59.9 | 3.86 | 59.9 |
| 17—CHβ | 3.88 | 65.1 | 3.88 | 65.1 | 3.88 | 65.1 | 3.87 | 65.1 |
| 17—OH | 4.70 | − | 4.69 | − | 4.69 | − | 4.69 | − |
| 18—CHγ | 1.03 | 20.0 | 1.03 | 20.1 | 1.03 | 20.1 | 1.03 | 20.1 |
| 19—NH | 8.09 | − | 8.09 | − | 8.09 | − | 8.09 | − |
| d-Ser | ||||||||
| 20—CO | − | 170.0 | − | 170.0 | − | 170.0 | − | 170.0 |
| 21—CHα | 4.38 | 54.5 | 4.38 | 54.6 | 4.38 | 54.5 | 4.38 | 54.5 |
| 22—CHβ | 3.54 | 61.6 | 3.54 | 61.5 | 3.54 | 61.5 | 3.54 | 61.5 |
| 22—OH | 4.86 | − | 4.86 | − | 4.86 | − | 4.86 | − |
| 23—NH | 7.53 | − | 7.53 | − | 7.53 | − | 7.53 | − |
| l-allo-Thr2 | ||||||||
| 24—CO | − | 168.4 | − | 168.4 | − | 168.4 | − | 168.4 |
| 25—CHα | 4.40 | 56.2 | 4.39 | 56.2 | 4.39 | 56.2 | 4.39 | 56.2 |
| 26—CHβ | 5.35 | 69.6 | 5.34 | 69.6 | 5.34 | 69.6 | 5.35 | 69.6 |
| 27—CHγ | 1.10 | 17.0 | 1.10 | 17.0 | 1.10 | 16.9 | 1.10 | 17.0 |
| 28—NH | 8.40 | 8.39 | 8.39 | 8.39 | ||||
| Leu3 | ||||||||
| 29—CO | − | 173.1 | − | 173.2 | − | 173.1 | − | 173.2 |
| 30—CHα | 4.37 | 51.7 | 4.37 | 51.8 | 4.37 | 51.8 | 4.37 | 51.8 |
| 31—CHβ | 1.48 | 40.3 | 1.48 | 40.2 | 1.48 | 40.2 | 1.48 | 40.2 |
| 32—CHγ | 1.62 | 24.2 | 1.62 | 24.2 | 1.62 | 24.2 | 1.63 | 24.2 |
| 33—CHδ,1 | 0.92 | 22.6 | 0.92 | 22.6 | 0.91 | 22.6 | 0.92 | 22.6 |
| 34—CHδ,2 | 0.87 | 21.9 | 0.87 | 21.9 | 0.87 | 21.9 | 0.87 | 21.9 |
| 35—NH | 8.20 | − | 8.20 | − | 8.20 | − | 8.20 | − |
| Fatty acid | ||||||||
| 36—CO | − | 171.8 | − | 171.9 | − | 171.9 | − | 171.9 |
| 37 | ~2.24 | 43.3 | 2.24, 2.22 | 43.4 | 2.25, 2.22 | 43.4 | 2.25, 2.22 | 43.4 |
| 38 | 3.78 | 67.4 | 3.77 | 67.5 | 3.78 | 67.5 | 3.78 | 67.5 |
| 38—OH | 4.62 | − | 4.60 | − | 4.60 | − | 4.59 | − |
| 39 | 1.34, 1.30 | 36.2 | 1.31 | 36.6 | 1.32 | 36.6 | 1.32 | 36.6 |
| 40 | 1.42, 1.28 | 25.1 | 1.34, 1.22 | 24.8 | 1.36, 1.23 | 24.8 | 1.35, 1.22 | 24.8 |
| 41 | 1.96 | 26.6 | 1.23 | ~29 | ~1.2 | ~29 | 1.23 | 29.1 |
| 42 | 5.31 | 129.6 | ~1.2 | ~29 | 1.28 | ~29 | ~1.2 | ~29 |
| 43 | 5.31 | 129.6 | ~1.2 | ~29 | 1.97 | 26.6 | ~1.2 | ~29 |
| 44 | 1.96 | 26.5 | ~1.2 | ~29 | 5.31 | 129.6 | ~1.2 | ~29 |
| 45 | 1.28 | 29.0 | ~1.2 | ~29 | 5.31 | 129.6 | ~1.2 | ~29 |
| 46 | 1.24 | 28.3 | 1.21 | 29.1 | 1.97 | 26.6 | ~1.2 | ~29 |
| 47 | 1.22 | 31.1 | 1.22 | 31.3 | 1.28 | ~29 | ~1.2 | ~29 |
| 48 | 1.25 | 22.1 | 1.25 | 22.1 | 1.25 | 28.2 | ~1.2 | ~29 |
| 49 | 0.84 | 13.9 | 0.84 | 13.9 | 1.22 | 31.1 | 1.22 | 31.3 |
| 50 | 1.25 | 22.0 | 1.25 | 22.1 | ||||
| 51 | 0.84 | 13.9 | 0.84 | 13.9 | ||||
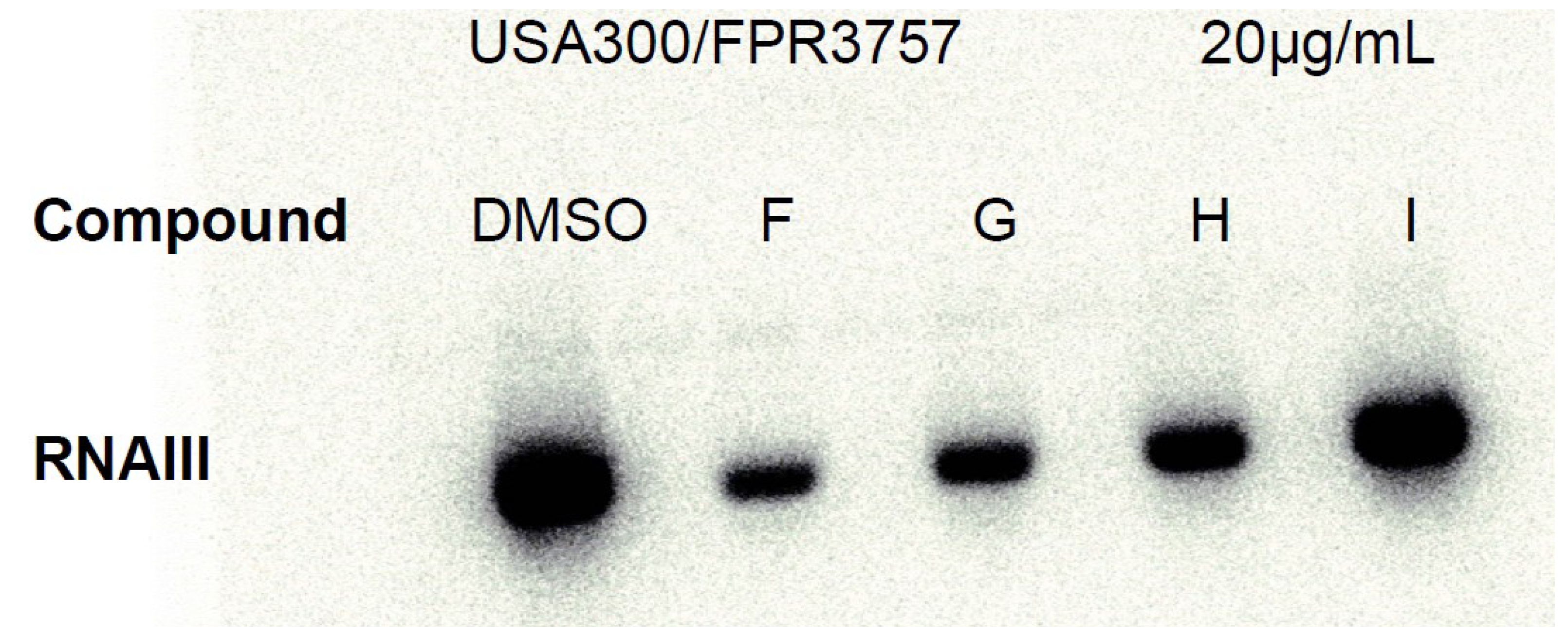
2.2. Ngercheumicins Interfere with agr


3. Experimental Section
3.1. Isolation and Identification of Strain s2753
3.2. Isolation and Structure Elucidation of Four New Ngercheumicins
3.3. Antivirulence Activity Testing and Northern Blotting
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Jensen, P.R.; Fenical, W. Marine bacterial diversity as a resource for novel microbial products. J. Ind. Microbiol. 1996, 17, 346–351. [Google Scholar]
- Gerwick, W.H.; Moore, B.S. Lessons from the Past and Charting the Future of Marine Natural Products Drug Discovery and Chemical Biology. Chem. Biol. 2012, 19, 85–98. [Google Scholar] [CrossRef]
- Gram, L.; Melchiorsen, J.; Bruhn, J.B. Antibacterial Activity of Marine Culturable Bacteria Collected from a Global Sampling of Ocean Surface Waters and Surface Swabs of Marine Organisms. Mar. Biotechnol. 2010, 12, 439–451. [Google Scholar] [CrossRef]
- Wietz, M.; Mansson, M.; Gotfredsen, C.H.; Larsen, T.O.; Gram, L. Antibacterial Compounds from Marine Vibrionaceae Isolated on a Global Expedition. Mar. Drugs 2010, 8, 2946–2960. [Google Scholar] [CrossRef]
- Mansson, M.; Nielsen, A.; Kjærulff, L.; Gotfredsen, C.H.; Wietz, M.; Ingmer, H.; Gram, L.; Larsen, T.O. Inhibition of Virulence Gene Expression in Staphylococcus aureus by Novel Depsipeptides from a Marine Photobacterium. Mar. Drugs 2011, 9, 2537–2552. [Google Scholar] [CrossRef]
- Rasko, D.A.; Sperandio, V. Anti-virulence strategies to combat bacteria-mediated disease. Nat. Rev. Drug Discov. 2010, 9, 117–128. [Google Scholar] [CrossRef]
- Novick, R.; Geisinger, E. Quorum Sensing in Staphylococci. Annu. Rev. Genet. 2008, 42, 541–564. [Google Scholar] [CrossRef]
- Adachi, K.; Kawabata, Y.; Kasai, H.; Katsuta, M.; Shizuri, Y. (Marine Biotechnol. Inst. Co. Ltd.). New Antibiotic. Jpn. Pat. Appl. JP 2007230911 A, 13 September 2007.
- Muir, T.W. Turning Virulence on and off in Staphylococci. J. Pept. Sci. 2003, 9, 612–619. [Google Scholar] [CrossRef]
- Lyon, G.J.; Mayville, P.; Muir, T.W.; Novick, R.P. Rational design of a global inhibitor of the virulence response in Staphylococcus aureus, based in part on localization of the site of inhibition to the receptor-histidine kinase, AgrC. Proc. Natl. Acad. Sci. USA 2000, 97, 13330–13335. [Google Scholar] [CrossRef]
- Mayville, P.; Ji, G.; Beavis, R.; Yang, H.; Goger, M.; Novick, R.P.; Muir, T.W. Structure-activity analysis of synthetic autoinducing thiolactone peptides from Staphylococcus aureus responsible for virulence. Proc. Natl. Acad. Sci. USA 1999, 96, 1218–1223. [Google Scholar] [CrossRef]
- Wright, J.S., III; Lyon, G.J.; George, E.A.; Muir, T.W.; Novick, R.P. Hydrophobic interactions drive ligand-receptor recognition for activation and inhibition of staphylococcal quorum sensing. Proc. Natl. Acad. Sci. USA 2004, 101, 16168–16173. [Google Scholar] [CrossRef]
- Gunstone, F.D.; Pollard, M.R.; Scrimgeour, C.M.; Vedanayagam, H.S. 13C Nuclear magnetic resonance studies of olefinic fatty acids and esters. Chem. Phys. Lipids 1977, 18, 115–129. [Google Scholar] [CrossRef]
- Fujii, K.; Ikao, Y.; Oka, H.; Suzuki, M.; Harada, K. A Nonempirical Method Using LC/MS for Determination of the Absolute Configuration of Constituent Amino Acids in a Peptide: Combination of Marfey’s Method with Mass Spectrometry and Its Practical Applications. Anal. Chem. 1997, 69, 5146–5151. [Google Scholar] [CrossRef]
- Bonnard, I.; Manzanares, I.; Rinehart, K.L. Stereochemistry of Kahalalide F. J. Nat. Prod. 2003, 66, 1466–1470. [Google Scholar] [CrossRef]
- Harada, K.; Fujii, K.; Mayumi, T.; Hibino, Y.; Suzuki, M. A Method Using LC/MS for Determination of Absolute Configuration of Constituent Amino Acids in Peptide—Advanced Marfey’s Method. Tetrahedron Lett. 1995, 36, 1515–1518. [Google Scholar] [CrossRef]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2013, 30, 237–323. [Google Scholar] [CrossRef]
- Molhoek, E.M.; van Dijk, A.; Veldhuizen, E.J.A.; Haagsman, H.P.; Bikker, F.J. Improved proteolytic stability of chicken cathelicidin-2 derived peptides by d-amino acid substitutions and cyclization. Peptides 2011, 32, 875–880. [Google Scholar] [CrossRef]
- Nielsen, A.; Nielsen, K.F.; Frees, D.; Larsen, T.O.; Ingmer, H. Method for Screening Compounds That Influence Virulence Gene Expression in Staphylococcus aureus. Antimicrob. Agents Chemother. 2010, 54, 509–512. [Google Scholar] [CrossRef]
- Ji, G.; Beavis, R.; Novick, R.P. Bacterial Interference Caused by Autoinducing Peptide Variants. Science 1997, 276, 2027–2030. [Google Scholar] [CrossRef]
- Thoendel, M.; Kavanaugh, J.S.; Flack, C.E.; Horswill, A.R. Peptide Signalling in the Staphylococci. Chem. Rev. 2011, 111, 117–151. [Google Scholar] [CrossRef]
- Williams, P. Quorum sensing, communication and cross-kingdom signalling in the bacterial world. Microbiology 2007, 153, 3923–3938. [Google Scholar] [CrossRef]
Supplementary Files
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Kjaerulff, L.; Nielsen, A.; Mansson, M.; Gram, L.; Larsen, T.O.; Ingmer, H.; Gotfredsen, C.H. Identification of Four New agr Quorum Sensing-Interfering Cyclodepsipeptides from a Marine Photobacterium. Mar. Drugs 2013, 11, 5051-5062. https://doi.org/10.3390/md11125051
Kjaerulff L, Nielsen A, Mansson M, Gram L, Larsen TO, Ingmer H, Gotfredsen CH. Identification of Four New agr Quorum Sensing-Interfering Cyclodepsipeptides from a Marine Photobacterium. Marine Drugs. 2013; 11(12):5051-5062. https://doi.org/10.3390/md11125051
Chicago/Turabian StyleKjaerulff, Louise, Anita Nielsen, Maria Mansson, Lone Gram, Thomas O. Larsen, Hanne Ingmer, and Charlotte H. Gotfredsen. 2013. "Identification of Four New agr Quorum Sensing-Interfering Cyclodepsipeptides from a Marine Photobacterium" Marine Drugs 11, no. 12: 5051-5062. https://doi.org/10.3390/md11125051
APA StyleKjaerulff, L., Nielsen, A., Mansson, M., Gram, L., Larsen, T. O., Ingmer, H., & Gotfredsen, C. H. (2013). Identification of Four New agr Quorum Sensing-Interfering Cyclodepsipeptides from a Marine Photobacterium. Marine Drugs, 11(12), 5051-5062. https://doi.org/10.3390/md11125051