Microbial Communities and Bioactive Compounds in Marine Sponges of the Family Irciniidae—A Review

Abstract
: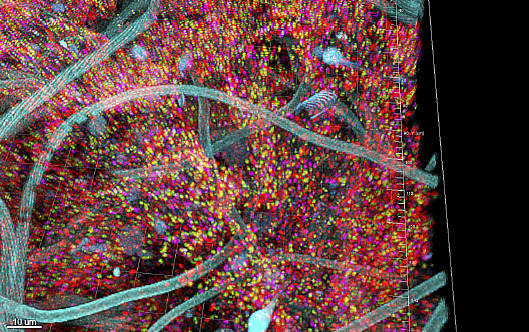
1. Introduction

{kind=link}
{kind=link}
{kind=link}
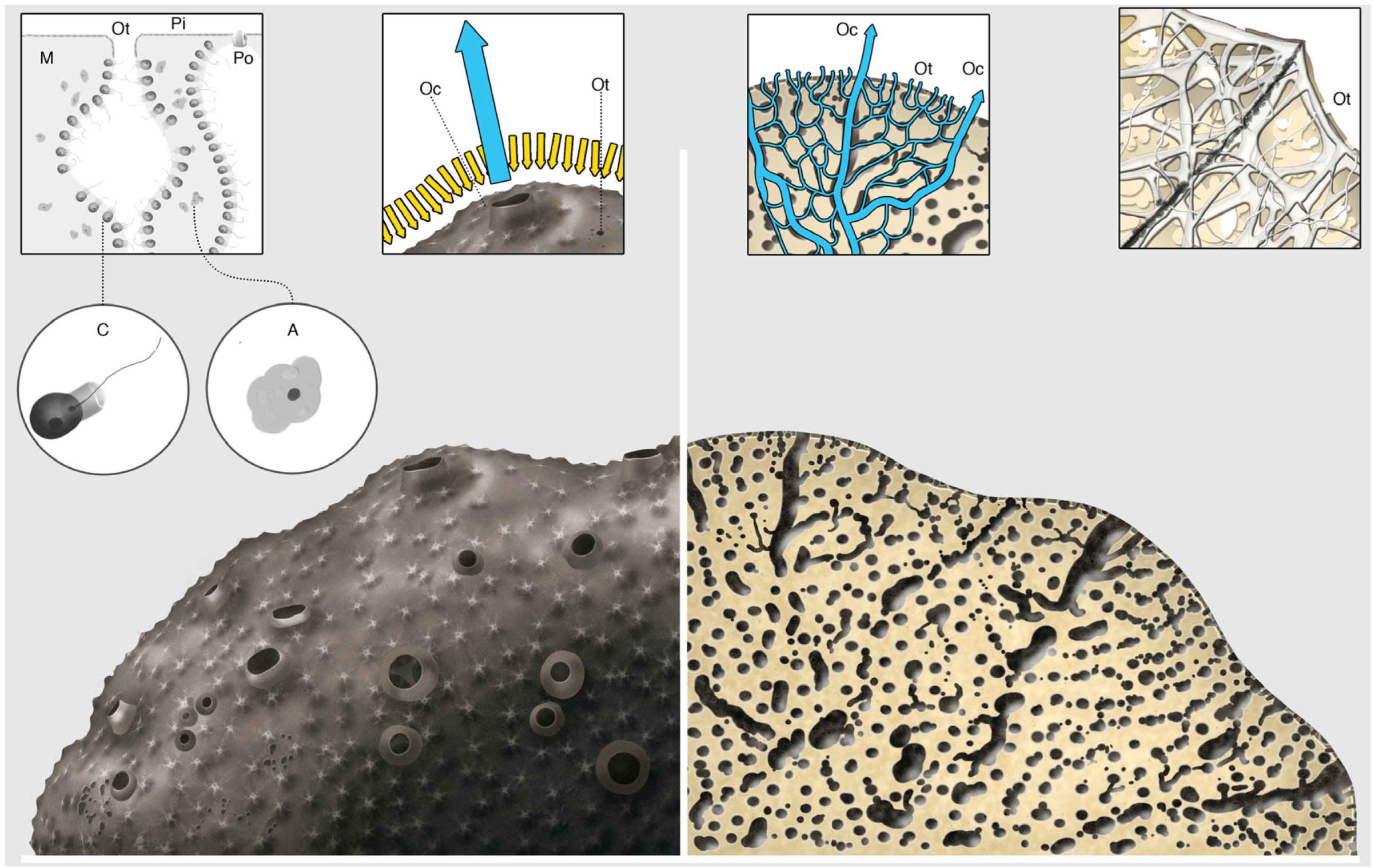{kind=link}
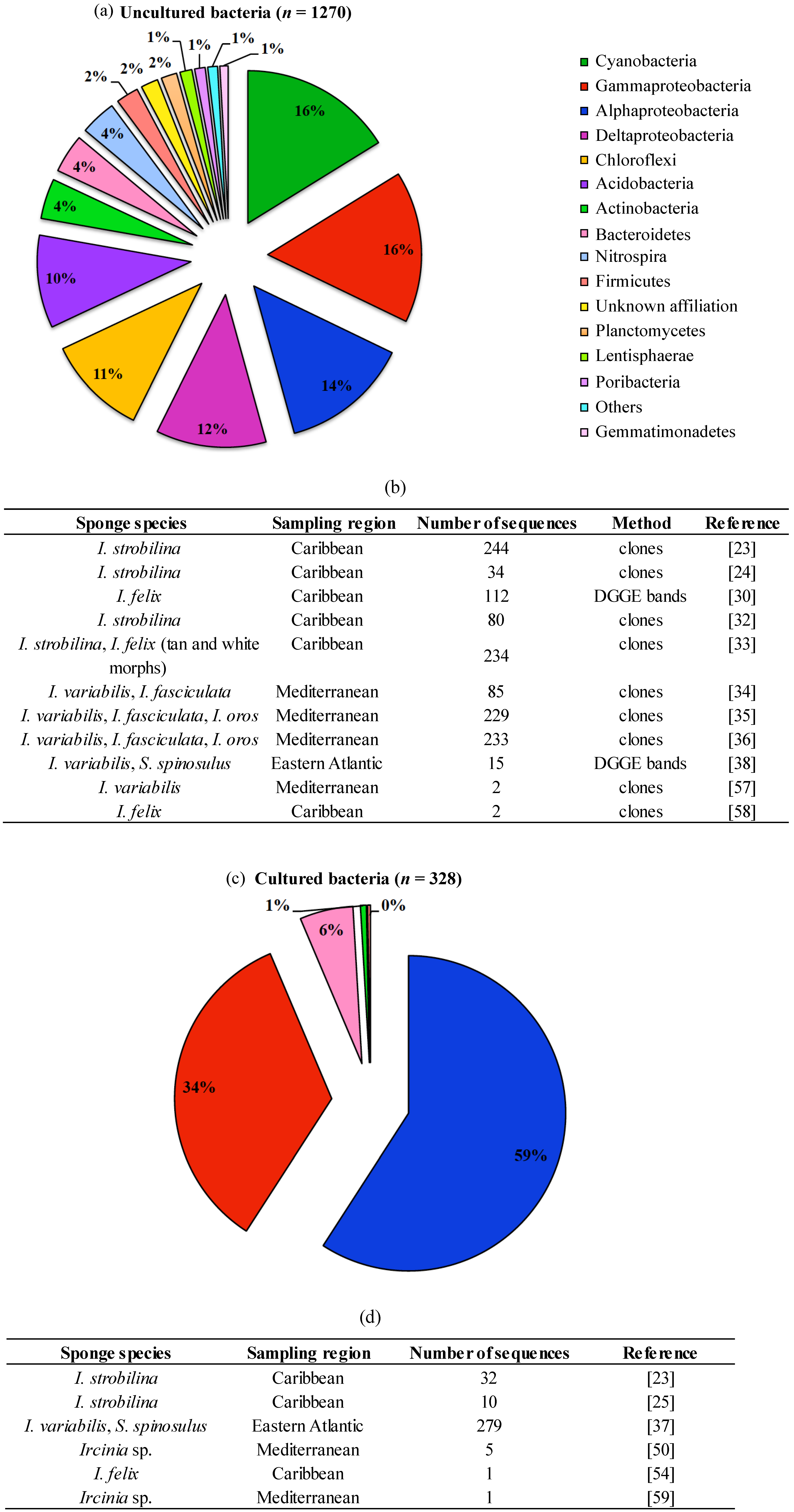{kind=link}
{kind=link}
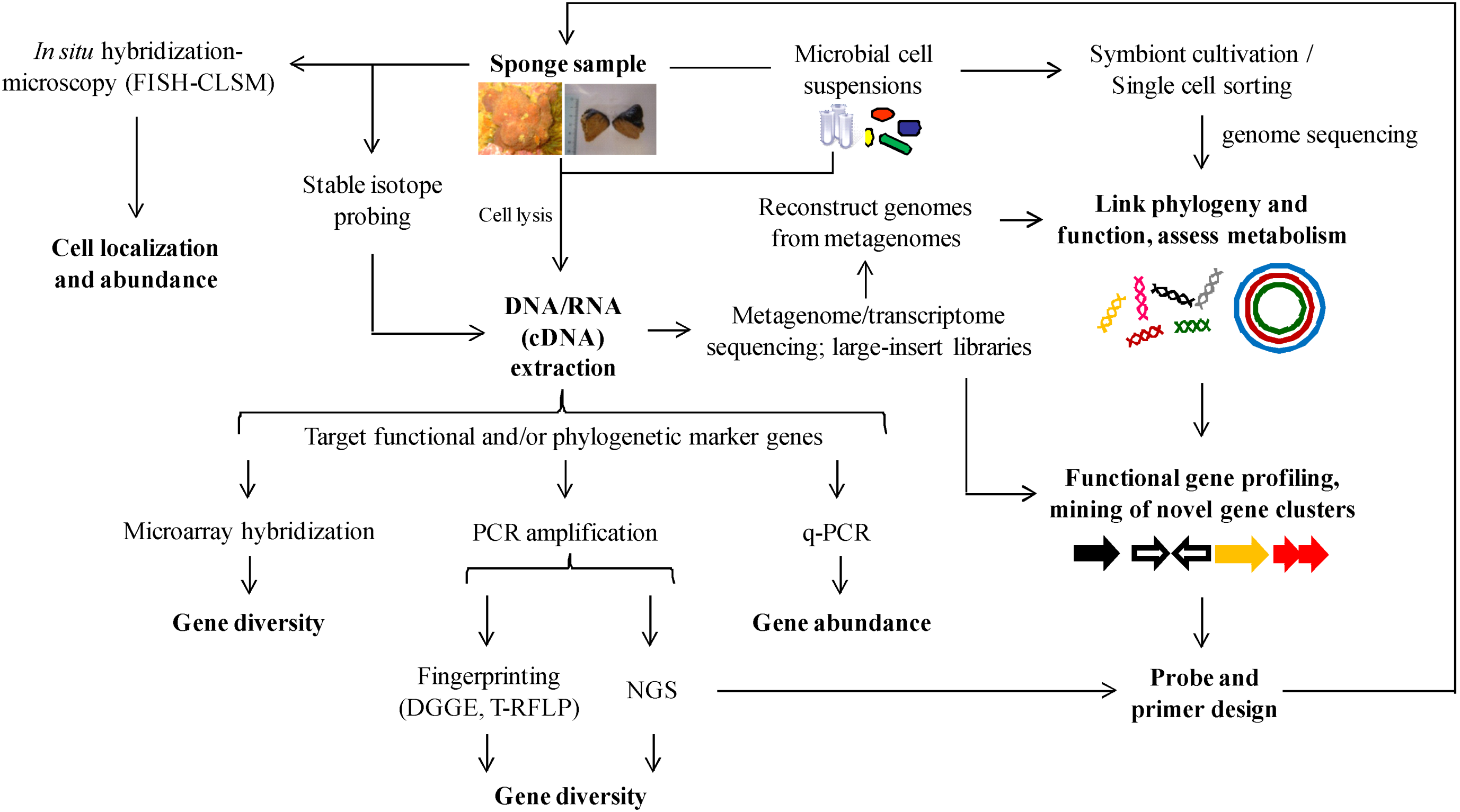{kind=link}
| Realms 1 | Provinces | Ecoregions | Number of Species 2 |
|---|---|---|---|
| Tropical Atlantic | Tropical North-western Atlantic | Eastern Caribbean | 208 |
| Tropical Atlantic | Tropical North-western Atlantic | Greater Antilles | 275 |
| Temperate Northern Atlantic | Northern European Seas | Celtic Seas | 295 |
| Temperate Northern Atlantic | Lusitanian | South European Atlantic Shelf | 231 |
| Temperate Northern Atlantic | Lusitanian | Azores, Canaries, Madeira | 458 |
| Temperate Northern Atlantic | Mediterranean Sea | Western Mediterranean | 461 |
| Western Indo-Pacific | West and South Indian Shelf | South India and Sri Lanka | 211 |
| Temperate Northern Pacific | Warn Temperate Northwest Atlantic | Central Kuroshio Current | 377 |
| Central Indo-Pacific | Tropical South-western Pacific | New Caledonia | 256 |
| Temperate Australasia | East Central Australian Shelf | Manning-Hawkesbury | 278 |
| Temperate Australasia | Southeast Australian Shelf | Bassian | 375 |

2. The Irciniidae Family as a Model Taxon in Sponge Microbiology Research
- The existence of several sympatric species within the family allows testing the host species-specific hypothesis of microbiome composition in sponges using a solid phylogenetic background;
- The wide geographical distribution of some species permits testing hypotheses of symbiont/metabolite maintenance across biogeographical gradients;
- Irciniidae species are chemically rich, a feature that might relate with microbial diversity and suggests biotechnological potential;
- Irciniidae species have been reported as high microbial abundance sponges. This suggests high metabolic activity and/or selective enrichment of their symbionts, which most likely play essential roles for host fitness and survival;
- Bacterial species from at least 15 phyla have been found to be enriched in Irciniidae hosts when compared with surrounding seawater. This distinct microbiome composition is indicative of close host-symbiont relationships;
- The order Dictyoceratida consists of four families—including Irciniidae—collectively known as “keratose” sponges because their skeleton is made of a complex matrix of protein fibres instead of mineral elements (i.e., true spicules), making these families interesting representatives of a peculiar life strategy within marine sponges.


3. Microbial Diversity and Bioactivities in the Family Irciniidae
3.1. Early Microbiology Studies
3.2. Bacteria
3.2.1. Diversity
3.2.1.1. Culture-Dependent Approaches
3.2.1.2. Cultivation-Independent Approaches


3.2.2. Bioactivities
3.2.3. Bioactive Compounds
3.3. Archaea
3.3.1. Diversity
3.3.2. Bioactivities and Bioactive Compounds
3.4. Fungi
3.4.1. Diversity
3.4.2. Bioactivities
3.4.3. Bioactive Compounds
3.5. Other Microeukaryotes
4. Vertical Transmission of Sponge Symbionts
5. Bacterial Communication and Signalling Molecules

6. Diseases Affecting Irciniids
7. Experimental Microbial Ecology
8. Metagenomics-Based Discovery of Secondary Metabolites Biosynthetic Gene Clusters
9. Cultivation of Irciniidae Species
9.1. Sponge Mariculture
9.2. Sponge Cell Culture
9.3. Sponge Larviculture
10. Concluding Remarks and Outlook
Supplementary Files
Supplementary File 1Supplementary File 2Acknowledgments
Author Contributions
Conflicts of Interest
References
- Li, C.W.; Chen, J.Y.; Hua, T.E. Precambrian sponges with cellular structures. Science 1998, 279, 879–882. [Google Scholar] [PubMed]
- Van Soest, R.W.M.; Boury-Esnault, N.; Vacelet, J.; Dohrmann, M.; Erpenbeck, D.; de Voogd, N.J.; Santodomingo, N.; Vanhoorne, B.; Kelly, M.; Hooper, J.N.A. Global diversity of sponges (Porifera). PLoS One 2012, 7, e35105. [Google Scholar] [PubMed]
- Love, G.D.; Grosjean, E.; Stalvies, C.; Fike, D.A.; Grotzinger, J.P.; Bradley, A.S.; Kelly, A.E.; Bhatia, M.; Meredith, W.; Snape, C.E.; et al. Fossil steroids record the appearance of Demospongiae during the Cryogenian period. Nature 2009, 457, 718–722. [Google Scholar]
- Vacelet, J.; Donadey, C. Electron-microscope study of association between some sponges and Bacteria. J. Exp. Mar. Biol. Ecol. 1977, 30, 301–314. [Google Scholar]
- Hentschel, U.; Piel, J.; Degnan, S.M.; Taylor, M.W. Genomic insights into the marine sponge microbiome. Nat. Rev. Microbiol. 2012, 10, 641–675. [Google Scholar] [PubMed]
- Taylor, M.W.; Radax, R.; Steger, D.; Wagner, M. Sponge-associated microorganisms: Evolution, ecology, and biotechnological potential. Microbiol. Mol. Biol. Rev. 2007, 71, 295–347. [Google Scholar] [PubMed]
- Spalding, M.D.; Fox, H.E.; Halpern, B.S.; McManus, M.A.; Molnar, J.; Allen, G.R.; Davidson, N.; Jorge, Z.A.; Lombana, A.L.; Lourie, S.A.; et al. Marine ecoregions of the world: A bioregionalization of coastal and shelf areas. Bioscience 2007, 57, 573–583. [Google Scholar]
- Webster, N.S.; Taylor, M.W. Marine sponges and their microbial symbionts: Love and other relationships. Environ. Microbiol. 2012, 14, 335–346. [Google Scholar] [PubMed]
- Thomas, T.R.A.; Kavlekar, D.P.; LokaBharathi, P.A. Marine drugs from sponge-microbe association: A review. Mar. Drugs 2010, 8, 1417–1468. [Google Scholar] [PubMed]
- Taylor, M.W.; Hill, R.T.; Hentschel, U. Meeting report: 1st International Symposium on Sponge Microbiology. Mar. Biotechnol. 2011, 13, 1057–1061. [Google Scholar] [PubMed]
- Bayer, K.; Scheuermayer, M.; Fieseler, L.; Hentschel, U. Genomic mining for novel FADH2-dependent halogenases in marine sponge-associated microbial consortia. Mar. Biotechnol. 2013, 15, 63–72. [Google Scholar] [PubMed]
- Fieseler, L.; Horn, M.; Wagner, M.; Hentschel, U. Discovery of the novel candidate phylum “Poribacteria” in marine sponges. Appl. Environ. Microbiol. 2004, 70, 3724–3732. [Google Scholar] [PubMed]
- Hentschel, U.; Hopke, J.; Horn, M.; Friedrich, A.B.; Wagner, M.; Hacker, J.; Moore, B.S. Molecular evidence for a uniform microbial community in sponges from different oceans. Appl. Environ. Microbiol. 2002, 68, 4431–4440. [Google Scholar] [PubMed]
- Schmitt, S.; Tsai, P.; Bell, J.; Fromont, J.; Ilan, M.; Lindquist, N.; Perez, T.; Rodrigo, A.; Schupp, P.J.; Vacelet, J.; et al. Assessing the complex sponge microbiota: Core, variable and species-specific bacterial communities in marine sponges. ISME J. 2012, 6, 564–576. [Google Scholar]
- Siegl, A.; Hentschel, U. PKS and NRPS gene clusters from microbial symbiont cells of marine sponges by whole genome amplification. Environ. Microbiol. Rep. 2010, 2, 507–513. [Google Scholar] [PubMed]
- Fan, L.; Liu, M.; Simister, R.; Webster, N.S.; Thomas, T. Marine microbial symbiosis heats up: The phylogenetic and functional response of a sponge holobiont to thermal stress. ISME J. 2013, 7, 991–1002. [Google Scholar] [PubMed]
- Pantile, R.; Webster, N. Strict thermal threshold identified by quantitative PCR in the sponge Rhopaloeides odorabile. Mar. Ecol. Prog. Ser. 2011, 431, 97–105. [Google Scholar]
- Simister, R.; Taylor, M.W.; Tsai, P.; Webster, N. Sponge-microbe associations survive high nutrients and temperatures. PLoS One 2012, 7, e52220. [Google Scholar]
- Webster, N.S.; Cobb, R.E.; Negri, A.P. Temperature thresholds for bacterial symbiosis with a sponge. ISME J. 2008, 2, 830–842. [Google Scholar] [PubMed]
- Webster, N.S.; Taylor, M.W.; Behnam, F.; Lucker, S.; Rattei, T.; Whalan, S.; Horn, M.; Wagner, M. Deep sequencing reveals exceptional diversity and modes of transmission for bacterial sponge symbionts. Environ. Microbiol. 2010, 12, 2070–2082. [Google Scholar] [PubMed]
- Mohamed, N.M.; Colman, A.S.; Tal, Y.; Hill, R.T. Diversity and expression of nitrogen fixation genes in bacterial symbionts of marine sponges. Environ. Microbiol. 2008, 10, 2910–2921. [Google Scholar] [PubMed]
- Mohamed, N.; Cicirelli, E.; Kan, J.; Chen, F.; Fuqua, C.; Hill, R. Diversity and quorum-sensing signal production of Proteobacteria associated with marine sponges. Environ. Microbiol. 2008, 10, 75–86. [Google Scholar] [PubMed]
- Mohamed, N.; Rao, V.; Hamann, M.; Kelly, M.; Hill, R. Monitoring bacterial diversity of the marine sponge Ircinia strobilina upon transfer into aquaculture. Appl. Environ. Microbiol. 2008, 74, 4133–4143. [Google Scholar] [PubMed]
- Mohamed, N.M.; Saito, K.; Tal, Y.; Hill, R.T. Diversity of aerobic and anaerobic ammonia-oxidizing bacteria in marine sponges. ISME J. 2010, 4, 38–48. [Google Scholar] [PubMed]
- Zan, J.D.; Fuqua, C.; Hill, R.T. Diversity and functional analysis of luxS genes in vibrios from marine sponges Mycale laxissima and Ircinia strobilina. ISME J. 2011, 5, 1505–1516. [Google Scholar] [PubMed]
- Van Soest, R.W.M.; Boury-Esnault, N.; Hooper, J.N.A.; Rützler, K.; de Voogd, N.J.; Alvarez de Glasby, B.; Hajdu, E.; Pisera, A.B.; Manconi, R.; Schoenberg, C.; et al. World Porifera Database. Available online: http://www.marinespecies.org/porifera (accessed on 20 December 2013).
- Cook, S.C.; Bergquist, P.R. Family Irciniidae Gray, 1867. In System Porifera: A Guide to the Classification of Sponges; Hooper, J.N.A., van Soest, R.W.M., Eds.; Kluwer Academic/Plenum Publishers: New York, NY, USA, 2002; pp. 1022–1027. [Google Scholar]
- Cook, S.; Bergquist, P. New species of dictyoceratid sponges from New Zealand: Genus Ircinia (Porifera: Demospongiae: Dictyoceratida). N. Z. J. Mar. Freshw. Res. 1999, 33, 545–563. [Google Scholar]
- Wörheide, G.; Dohrmann, M.; Erpenbeck, D.; Larroux, C.; Maldonado, M.; Voigt, O.; Borchiellini, C.; Lavrov, D.V. Deep phylogeny and evolution of sponges (Phylum Porifera). In Advances in Marine Biology; Becerro, M.A., Uriz, M.J., Maldonado, M., Turon, X., Eds.; Academic Press: Amsterdam, The Netherlands, 2012; pp. 1–18. [Google Scholar]
- Schmitt, S.; Weisz, J.; Lindquist, N.; Hentschel, U. Vertical transmission of a phylogenetically complex microbial consortium in the viviparous sponge Ircinia felix. Appl. Environ. Microbiol. 2007, 73, 2067–2078. [Google Scholar] [PubMed]
- Weisz, J.; Hentschel, U.; Lindquist, N.; Martens, C. Linking abundance and diversity of sponge-associated microbial communities to metabolic differences in host sponges. Mar. Biol. 2007, 152, 475–483. [Google Scholar]
- Yang, J.K.; Sun, J.; Lee, O.O.; Wong, Y.H.; Qian, P.Y. Phylogenetic diversity and community structure of sponge-associated bacteria from mangroves of the Caribbean Sea. Aquat. Microb. Ecol. 2011, 62, 231–240. [Google Scholar]
- Pita, L.; López-Legentil, S.; Erwin, P.M. Biogeography and host fidelity of bacterial communities in Ircinia spp. from the Bahamas. Microb. Ecol. 2013, 66, 437–447. [Google Scholar] [PubMed]
- Erwin, P.M.; López-Legentil, S.; González-Pech, R.; Turon, X. A specific mix of generalists: Bacterial symbionts in Mediterranean Ircinia spp. FEMS Microbiol. Ecol. 2012, 79, 619–637. [Google Scholar] [PubMed]
- Erwin, P.M.; Pita, L.; López-Legentil, S.; Turon, X. Stability of sponge-associated bacteria over large seasonal shifts in temperature and irradiance. Appl. Environ. Microbiol. 2012, 78, 7358–7368. [Google Scholar] [PubMed]
- Erwin, P.M.; López-Legentil, S.; Turon, X. Ultrastructure, molecular phylogenetics, and chlorophyll a content of novel cyanobacterial symbionts in temperate sponges. Microb. Ecol. 2012, 64, 771–783. [Google Scholar] [PubMed]
- Esteves, A.I.S.; Hardoim, C.C.P.; Xavier, J.R.; Gonçalves, J.M.S.; Costa, R. Molecular richness and biotechnological potential of bacteria cultured from Irciniidae sponges in the north-east Atlantic. FEMS Microbiol. Ecol. 2013, 85, 519–536. [Google Scholar] [PubMed]
- Hardoim, C.C.P.; Esteves, A.I.S.; Pires, F.R.; Gonçalves, J.M.S.; Cox, C.J.; Xavier, J.R.; Costa, R. Phylogenetically and spatially close marine sponges harbour divergent bacterial communities. PLoS One 2012, 7, e53029. [Google Scholar] [PubMed]
- Pita, L.; Turon, X.; López-Legentil, S.; Erwin, P.M. Host rules: Spatial stability of bacterial communities associated with marine sponges (Ircinia spp.) in the western Mediterranean Sea. FEMS Microbiol. Ecol. 2013, 86, 268–276. [Google Scholar] [PubMed]
- Usher, K.; Kuo, J.; Fromont, J.; Toze, S.; Sutton, D. Comparative morphology of five species of symbiotic and non-symbiotic coccoid Cyanobacteria. Eur. J. Phycol. 2006, 41, 179–188. [Google Scholar]
- Wilkinson, C.R. Microbial associations in sponges. III. Ultrastructure of in situ associations in coral-reef sponges. Mar. Biol. 1978, 49, 177–185. [Google Scholar]
- Webster, N.S.; Hill, R.T. The culturable microbial community of the Great Barrier Reef sponge Rhopaloeides odorabile is dominated by an alpha-proteobacterium. Mar. Biol. 2001, 138, 843–851. [Google Scholar]
- Hentschel, U.; Usher, K.M.; Taylor, M.W. Marine sponges as microbial fermenters. FEMS Microbiol. Ecol. 2006, 55, 167–177. [Google Scholar] [PubMed]
- Hardoim, C.C.P.; Costa, R.; Araújo, F.V.; Hajdu, E.; Peixoto, R.; Lins, U.; Rosado, A.S.; van Elsas, J.D. Diversity of bacteria in the marine sponge Aplysina fulva in Brazilian coastal waters. Appl. Environ. Microbiol. 2009, 75, 3331–3343. [Google Scholar] [PubMed]
- Kamke, J.; Taylor, M.W.; Schmitt, S. Activity profiles for marine sponge-associated bacteria obtained by 16S rRNA vs. 16S rRNA gene comparisons. ISME J. 2010, 4, 498–508. [Google Scholar] [PubMed]
- Sara, M. Ultrastructural aspects of symbiosis between two species of genus Aphanocapsa (Cyanophyceae) and Ircinia variabilis (Demospongiae). Mar. Biol. 1971, 11, 214–221. [Google Scholar]
- Wilkinson, C.R. Microbial associations in sponges. I. Ecology, physiology and microbial-populations of coral-reef sponges. Mar. Biol. 1978, 49, 161–167. [Google Scholar]
- Wilkinson, C.R. Microbial associations in sponges. II. Numerical-analysis of sponge and water bacterial populations. Mar. Biol. 1978, 49, 169–176. [Google Scholar]
- Hentschel, U.; Schmid, M.; Wagner, M.; Fieseler, L.; Gernert, C.; Hacker, J. Isolation and phylogenetic analysis of bacteria with antimicrobial activities from the Mediterranean sponges Aplysina aerophoba and Aplysina cavernicola. FEMS Microbiol. Ecol. 2001, 35, 305–312. [Google Scholar] [PubMed]
- Muscholl-Silberhorn, A.; Thiel, V.; Imhoff, J.F. Abundance and bioactivity of cultured sponge-associated bacteria from the Mediterranean Sea. Microb. Ecol. 2008, 55, 94–106. [Google Scholar] [PubMed]
- Thiel, V.; Imhoff, J.F. Phylogenetic identification of bacteria with antimicrobial activities isolated from Mediterranean sponges. Biomol. Eng. 2003, 20, 421–423. [Google Scholar] [PubMed]
- Enticknap, J.J.; Kelly, M.; Peraud, O.; Hill, R.T. Characterization of a culturable alphaproteobacterial symbiont common to many marine sponges and evidence for vertical transmission via sponge larvae. Appl. Environ. Microbiol. 2006, 72, 3724–3732. [Google Scholar] [PubMed]
- O’Halloran, J.A.; Barbosa, T.M.; Morrissey, J.P.; Kennedy, J.; O’Gara, F.; Dobson, A.D.W. Diversity and antimicrobial activity of Pseudovibrio spp. from Irish marine sponges. J. Appl. Microbiol. 2011, 110, 1495–1508. [Google Scholar] [PubMed]
- Tabares, P.; Pimentel-Elardo, S.M.; Schirmeister, T.; Huenig, T.; Hentschel, U. Anti-protease and immunomodulatory activities of bacteria associated with Caribbean sponges. Mar. Biotechnol. 2011, 13, 883–892. [Google Scholar] [PubMed]
- Friedrich, A.B.; Fischer, I.; Proksch, P.; Hacker, J.; Hentschel, U. Temporal variation of the microbial community associated with the Mediterranean sponge Aplysina aerophoba. FEMS Microbiol. Ecol. 2001, 38, 105–113. [Google Scholar]
- Hugenholtz, P. Exploring prokaryotic diversity in the genomic era. Genome Biol. 2002, 3. [Google Scholar] [CrossRef]
- Usher, K.; Fromont, J.; Sutton, D.; Toze, S. The biogeography and phylogeny of unicellular cyanobacterial symbionts in sponges from Australia and the Mediterranean. Microb. Ecol. 2004, 48, 167–177. [Google Scholar] [PubMed]
- Schmitt, S.; Angermeier, H.; Schiller, R.; Lindquist, N.; Hentschel, U. Molecular microbial diversity survey of sponge reproductive stages and mechanistic insights into vertical transmission of microbial symbionts. Appl. Environ. Microbiol. 2008, 74, 7694–7708. [Google Scholar] [PubMed]
- Stabili, L.; Cardone, F.; Alifano, P.; Tredici, S.M.; Piraino, S.; Corriero, G.; Gaino, E. Epidemic mortality of the sponge Ircinia variabilis (Schmidt, 1862) associated to proliferation of a Vibrio bacterium. Microb. Ecol. 2012, 64, 802–813. [Google Scholar] [PubMed]
- Lee, O.O.; Wang, Y.; Yang, J.K.; Lafi, F.F.; Al-Suwailem, A.; Qian, P.Y. Pyrosequencing reveals highly diverse and species-specific microbial communities in sponges from the Red Sea. ISME J. 2011, 5, 650–664. [Google Scholar] [PubMed]
- Hardoim, C.C.P.; Cardinale, M.; Cúcio, A.; Esteves, A.I.S.; Berg, G.; Xavier, J.R.; Cox, C.; Costa, R. Bacterial cultivation bias examined for keratose marine sponges. Unpublised work. 2014. [Google Scholar]
- Hardoim, C.C.P.; Costa, R. Temporal dynamics of prokaryotic communities in the marine sponge Sarcotragus spinosulus. Mol. Ecol. 2014, 23, 3097–3112. [Google Scholar] [PubMed]
- Fiore, C.L.; Jarett, J.K.; Olson, N.D.; Lesser, M.P. Nitrogen fixation and nitrogen transformations in marine symbioses. Trends Microbiol. 2010, 18, 455–463. [Google Scholar] [PubMed]
- Zehr, J.P.; Mellon, M.T.; Zani, S. New nitrogen-fixing microorganisms detected in oligotrophic oceans by amplification of nitrogenase (nifH) genes. Appl. Environ. Microbiol. 1998, 64, 3444–3450. [Google Scholar] [PubMed]
- Zehr, J.P.; Jenkins, B.D.; Short, S.M.; Steward, G.F. Nitrogenase gene diversity and microbial community structure: A cross-system comparison. Environ. Microbiol. 2003, 5, 539–554. [Google Scholar] [PubMed]
- Man-Aharonovich, D.; Kress, N.; Bar Zeev, E.; Berman-Frank, I.; Beja, O. Molecular ecology of nifH genes and transcripts in the eastern Mediterranean Sea. Environ. Microbiol. 2007, 9, 2354–2363. [Google Scholar] [PubMed]
- Thakur, N.; Anil, A. Antibacterial activity of the sponge Ircinia ramosa: Importance of its surface-associated bacteria. J. Chem. Ecol. 2000, 26, 57–71. [Google Scholar]
- Thakur, N.; Anil, A.; Müller, W. Culturable epibacteria of the marine sponge Ircinia fusca: Temporal variations and their possible role in the epibacterial defense of the host. Aquat. Microb. Ecol. 2004, 37, 295–304. [Google Scholar]
- Pandey, S.; Sree, A.; Dash, S.S.; Sethi, D.P.; Chowdhury, L. Diversity of marine bacteria producing beta-glucosidase inhibitors. Microb. Cell. Fact. 2013, 12. [Google Scholar] [CrossRef]
- Asano, N. Glycosidase inhibitors: Update and perspectives on practical use. Glycobiology 2003, 13, 93R–104R. [Google Scholar] [PubMed]
- De Rosa, S.; Mitova, M.; Tommonaro, G. Marine bacteria associated with sponge as source of cyclic peptides. Biomol. Eng. 2003, 20, 311–316. [Google Scholar]
- Holden, M.T.G.; Chhabra, S.R.; de Nys, R.; Stead, P.; Bainton, N.J.; Hill, P.J.; Manefield, M.; Kumar, N.; Labatte, M.; England, D.; et al. Quorum-sensing cross talk: Isolation and chemical characterization of cyclic dipeptides from Pseudomonas aeruginosa and other gram-negative bacteria. Mol. Microbiol. 1999, 33, 1254–1266. [Google Scholar] [PubMed]
- Degrassi, G.; Aguilar, C.; Bosco, M.; Zahariev, S.; Pongor, S.; Venturi, V. Plant growth-promoting Pseudomonas putida WCS358 produces and secretes four cyclic dipeptides: Cross-talk with quorum sensing bacterial sensors. Curr. Microbiol. 2002, 45, 250–254. [Google Scholar] [PubMed]
- Kalinovskaya, N.; Kuznetsova, T.; Rashkes, Y.; Milgrom, Y.; Milgrom, E.; Willis, R.; Wood, A.; Kurtz, H.; Carabedian, C.; Murphy, P.; et al. Surfactin-like structures of five cyclic depsipeptides from the marine isolate of Bacillus pumilus. Russ. Chem. Bull. 1995, 44, 951–955. [Google Scholar]
- Prokof'eva, N.; Kalinovskaya, N.; Luk’yanov, P.; Shentsova, E.; Kuznetsova, T. The membranotropic activity of cyclic acyldepsipeptides from bacterium Bacillus pumilus, associated with the marine sponge Ircinia sp. Toxicon 1999, 37, 801–813. [Google Scholar] [PubMed]
- Simister, R.L.; Deines, P.; Botte, E.S.; Webster, N.S.; Taylor, M.W. Sponge-specific clusters revisited: A comprehensive phylogeny of sponge-associated microorganisms. Environ. Microbiol. 2012, 14, 517–524. [Google Scholar] [PubMed]
- Brochier-Armanet, C.; Boussau, B.; Gribaldo, S.; Forterre, P. Mesophilic crenarchaeota: Proposal for a third archaeal phylum, the Thaumarchaeota. Nat. Rev. Microbiol. 2008, 6, 245–252. [Google Scholar] [PubMed]
- Spang, A.; Hatzenpichler, R.; Brochier-Armanet, C.; Rattei, T.; Tischler, P.; Spieck, E.; Streit, W.; Stahl, D.A.; Wagner, M.; Schleper, C. Distinct gene set in two different lineages of ammonia-oxidizing Archaea supports the phylum Thaumarchaeota. Trends Microbiol. 2010, 18, 331–340. [Google Scholar] [PubMed]
- Pester, M.; Schleper, C.; Wagner, M. The Thaumarchaeota: An emerging view of their phylogeny and ecophysiology. Curr. Opin. Microbiol. 2011, 14, 300–306. [Google Scholar] [PubMed]
- Lee, E.Y.; Lee, H.K.; Lee, Y.K.; Sim, C.J.; Lee, J.H. Diversity of symbiotic archaeal communities in marine sponges from Korea. Biomol. Eng. 2003, 20, 299–304. [Google Scholar] [PubMed]
- Webster, N.S.; Watts, J.E.M.; Hill, R.T. Detection and phylogenetic analysis of novel crenarchaeote and euryarchaeote 16S ribosomal RNA gene sequences from a Great Barrier Reef sponge. Mar. Biotechnol. 2001, 3, 600–608. [Google Scholar] [PubMed]
- Webster, N.S.; Negri, A.P.; Munro, M.; Battershill, C.N. Diverse microbial communities inhabit Antarctic sponges. Environ. Microbiol. 2004, 6, 288–300. [Google Scholar] [PubMed]
- Hoffmann, F.; Radax, R.; Woebken, D.; Holtappels, M.; Lavik, G.; Rapp, H.T.; Schlappy, M.L.; Schleper, C.; Kuypers, M.M.M. Complex nitrogen cycling in the sponge Geodia barretti. Environ. Microbiol. 2009, 11, 2228–2243. [Google Scholar] [PubMed]
- Han, M.Q.; Liu, F.; Zhang, F.L.; Li, Z.Y.; Lin, H.W. Bacterial and archaeal symbionts in the south China Sea sponge Phakellia fusca: Community structure, relative abundance, and ammonia-oxidizing populations. Mar. Biotechnol. 2012, 14, 701–713. [Google Scholar] [PubMed]
- Cardoso, J.; van Bleijswijk, J.D.L.; Witte, H.; van Duyl, F.C. Diversity and abundance of ammonia-oxidizing Archaea and Bacteria in tropical and cold-water coral reef sponges. Aquat. Microb. Ecol. 2013, 68, 215–230. [Google Scholar]
- Höller, U.; Wright, A.D.; Matthee, G.F.; Konig, G.M.; Draeger, S.; Aust, H.J.; Schulz, B. Fungi from marine sponges: Diversity, biological activity and secondary metabolites. Mycol. Res. 2000, 104, 1354–1365. [Google Scholar]
- Paz, Z.; Komon-Zelazowska, M.; Druzhinina, I.S.; Aveskamp, M.M.; Shnaiderman, A.; Aluma, Y.; Carmeli, S.; Ilan, M.; Yarden, O. Diversity and potential antifungal properties of fungi associated with a Mediterranean sponge. Fungal Divers. 2010, 42, 17–26. [Google Scholar]
- Bringmann, G.; Lang, G.; Gulder, T.; Tsuruta, H.; Muhlbacher, J.; Maksimenka, K.; Steffens, S.; Schaumann, K.; Stohr, R.; Wiese, J.; et al. The first sorbicillinoid alkaloids, the antileukemic sorbicillactones A and B, from a sponge-derived Penicillium chrysogenum strain. Tetrahedron 2005, 61, 7252–7265. [Google Scholar]
- Ereskovsky, A.V.; Gonobobleva, E.; Vishnyakov, A. Morphological evidence for vertical transmission of symbiotic bacteria in the viviparous sponge Halisarca dujardini Johnston (Porifera, Demospongiae, Halisarcida). Mar. Biol. 2005, 146, 869–875. [Google Scholar]
- Sharp, K.H.; Eam, B.; Faulkner, D.J.; Haygood, M.G. Vertical transmission of diverse microbes in the tropical sponge Corticium sp. Appl. Environ. Microbiol. 2007, 73, 622–629. [Google Scholar] [PubMed]
- Lee, O.O.; Chui, P.Y.; Wong, Y.H.; Pawlik, J.R.; Qian, P.Y. Evidence for vertical transmission of bacterial symbionts from adult to embryo in the Caribbean sponge Svenzea zeai. Appl. Environ. Microbiol. 2009, 75, 6147–6156. [Google Scholar] [PubMed]
- Lindquist, N.; Bolser, R.; Laing, K. Timing of larval release by two Caribbean demosponges. Mar. Ecol. 1997, 155, 309–313. [Google Scholar]
- Lopanik, N.; Lindquist, N.; Targett, N. Potent cytotoxins produced by a microbial symbiont protect host larvae from predation. Oecologia 2004, 139, 131–139. [Google Scholar] [PubMed]
- Lopanik, N.; Gustafson, K.R.; Lindquist, N. Structure of bryostatin 20: A symbiont-produced chemical defense for larvae of the host bryozoan, Bugula neritina. J. Nat. Prod. 2004, 67, 1412–1414. [Google Scholar] [PubMed]
- Lopanik, N.B.; Targett, N.M.; Lindquist, N. Isolation of two polyketide synthase gene fragments from the uncultured microbial symbiont of the marine bryozoan Bugula neritina. Appl. Environ. Microbiol. 2006, 72, 7941–7944. [Google Scholar] [PubMed]
- Lopanik, N.B.; Shields, J.A.; Buchholz, T.J.; Rath, C.M.; Hothersall, J.; Haygood, M.G.; Hakansson, K.; Thomas, C.M.; Sherman, D.H. In vivo and in vitro trans-acylation by BryP, the putative bryostatin pathway acyltransferase derived from an uncultured marine symbiont. Chem. Biol. 2008, 15, 1175–1186. [Google Scholar] [PubMed]
- Sudek, S.; Lopanik, N.B.; Waggoner, L.E.; Hildebrand, M.; Anderson, C.; Liu, H.B.; Patel, A.; Sherman, D.H.; Haygood, M.G. Identification of the putative bryostatin polyketide synthase gene cluster from “Candidatus Endobugula sertula”, the uncultivated microbial symbiont of the marine bryozoan Bugula neritina. J. Nat. Prod. 2007, 70, 67–74. [Google Scholar] [PubMed]
- Jaeckle, W.B. Transport and metabolism of alanine and palmitic acid by field-collected larvae of Tedania ignis (Porifera, Demospongiae)—Estimated consequences of limited label translocation. Biol. Bull. 1995, 189, 159–167. [Google Scholar]
- Bandara, H.; Lam, O.L.T.; Jin, L.J.; Samaranayake, L. Microbial chemical signaling: A current perspective. Crit. Rev. Microbiol. 2012, 38, 217–249. [Google Scholar] [PubMed]
- Kalia, V.C. Quorum sensing inhibitors: An overview. Biotechnol. Adv. 2013, 31, 224–245. [Google Scholar] [PubMed]
- Skindersoe, M.E.; Ettinger-Epstein, P.; Rasmussen, T.B.; Bjarnsholt, T.; de Nys, R.; Givskov, M. Quorum sensing antagonism from marine organisms. Mar. Biotechnol. 2008, 10, 56–63. [Google Scholar] [PubMed]
- Dobretsov, S.; Teplitski, M.; Bayer, M.; Gunasekera, S.; Proksch, P.; Paul, V.J. Inhibition of marine biofouling by bacterial quorum sensing inhibitors. Biofouling 2011, 27, 893–905. [Google Scholar] [PubMed]
- Dieckmann, R.; Graeber, I.; Kaesler, I.; Szewzyk, U.; von Dohren, H. Rapid screening and dereplication of bacterial isolates from marine sponges of the Sula Ridge by Intact-Cell-MALDI-TOF mass spectrometry (ICM-MS). Appl. Microbiol. Biotechnol. 2005, 67, 539–548. [Google Scholar] [PubMed]
- Anand, T.P.; Bhat, A.W.; Shouche, Y.S.; Roy, U.; Siddharth, J.; Sarma, S.P. Antimicrobial activity of marine bacteria associated with sponges from the waters off the coast of south east India. Microbiol. Res. 2006, 161, 252–262. [Google Scholar] [PubMed]
- Hoffmann, M.; Fischer, M.; Ottesen, A.; McCarthy, P.J.; Lopez, J.V.; Brown, E.W.; Monday, S.R. Population dynamics of Vibrio spp. associated with marine sponge microcosms. ISME J. 2010, 4, 1608–1612. [Google Scholar] [PubMed]
- Flemer, B.; Kennedy, J.; Margassery, L.M.; Morrissey, J.P.; O’Gara, F.; Dobson, A.D.W. Diversity and antimicrobial activities of microbes from two Irish marine sponges, Suberites carnosus and Leucosolenia sp. J. Appl. Microbiol. 2012, 112, 289–301. [Google Scholar] [PubMed]
- Webster, N.S. Sponge disease: A global threat? Environ. Microbiol. 2007, 9, 1363–1375. [Google Scholar]
- Gaino, E.; Pronzato, R.; Corriero, G.; Buffa, P. Mortality of commercial sponges—Incidence in two Mediterranean areas. Boll. Zool. 1992, 59, 79–85. [Google Scholar]
- Wulff, J.L. Rapid diversity and abundance decline in a Caribbean coral reef sponge community. Biol. Conserv. 2006, 127, 167–176. [Google Scholar]
- Maldonado, M.; Sanchez-Tocino, L.; Navarro, C. Recurrent disease outbreaks in corneous demosponges of the genus Ircinia: Epidemic incidence and defense mechanisms. Mar. Biol. 2010, 157, 1577–1590. [Google Scholar]
- Cebrian, E.; Uriz, M.J.; Garrabou, J.; Ballesteros, E. Sponge mass mortalities in a warming Mediterranean sea: Are Cyanobacteria-harboring species worse off? PLoS One 2011, 6, e20211. [Google Scholar] [CrossRef] [Green Version]
- di Camillo, C.G.; Bartolucci, I.; Cerrano, C.; Bavestrello, G. Sponge disease in the Adriatic Sea. Mar. Ecol. 2013, 34, 62–71. [Google Scholar]
- Pita, L.; Erwin, P.M.; Turon, X.; López-Legentil, S. Till death do us part: Stable sponge-bacteria associations under thermal and food shortage stresses. PLoS One 2013, 8, e80307. [Google Scholar] [PubMed]
- Luter, H.M.; Whalan, S.; Webster, N.S. Prevalence of tissue necrosis and brown spot lesions in a common marine sponge. Mar. Freshw. Res. 2010, 61, 484–489. [Google Scholar]
- Mohamed, N.M.; Enticknap, J.J.; Lohr, J.E.; McIntosh, S.M.; Hill, R.T. Changes in bacterial communities of the marine sponge Mycale laxissima on transfer into aquaculture. Appl. Environ. Microbiol. 2008, 74, 1209–1222. [Google Scholar] [PubMed]
- Isaacs, L.T.; Kan, J.J.; Nguyen, L.; Videau, P.; Anderson, M.A.; Wright, T.L.; Hill, R.T. Comparison of the bacterial communities of wild and captive sponge Clathria prolifera from the Chesapeake Bay. Mar. Biotechnol. 2009, 11, 758–770. [Google Scholar] [PubMed]
- Thomas, T.; Rusch, D.; DeMaere, M.Z.; Yung, P.Y.; Lewis, M.; Halpern, A.; Heidelberg, K.B.; Egan, S.; Steinberg, P.D.; Kjelleberg, S. Functional genomic signatures of sponge bacteria reveal unique and shared features of symbiosis. ISME J. 2010, 4, 1557–1567. [Google Scholar] [PubMed]
- Fan, L.; Reynolds, D.; Liu, M.; Stark, M.; Kjelleberg, S.; Webster, N.S.; Thomas, T. Functional equivalence and evolutionary convergence in complex communities of microbial sponge symbionts. Proc. Natl. Acad. Sci. USA 2012, 109, E1878–E1887. [Google Scholar] [PubMed]
- Simister, R.; Taylor, M.W.; Tsai, P.; Fan, L.; Bruxner, T.J.; Crowe, M.L.; Webster, N. Thermal stress responses in the bacterial biosphere of the Great Barrier Reef sponge Rhopaloeides odorabile. Environ. Microbiol. 2012, 14, 3232–3246. [Google Scholar] [PubMed]
- Webster, N.; Pantile, R.; Botte, E.; Abdo, D.; Andreakis, N.; Whalan, S. A complex life cycle in a warming planet: Gene expression in thermally stressed sponges. Mol. Ecol. 2013, 22, 1854–1868. [Google Scholar] [PubMed]
- Duckworth, A.R.; Peterson, B.J. Effects of seawater temperature and pH on the boring rates of the sponge Cliona celata in scallop shells. Mar. Biol. 2013, 160, 27–35. [Google Scholar]
- Staunton, J.; Weissman, K.J. Polyketide biosynthesis: A millennium review. Nat. Prod. Rep. 2001, 18, 380–416. [Google Scholar] [PubMed]
- Piel, J.; Butzke, D.; Fusetani, N.; Hui, D.Q.; Platzer, M.; Wen, G.P.; Matsunaga, S. Exploring the chemistry of uncultivated bacterial symbionts: Antitumor polyketides of the pederin family. J. Nat. Prod. 2005, 68, 472–479. [Google Scholar] [PubMed]
- Schirmer, A.; Gadkari, R.; Reeves, C.D.; Ibrahim, F.; DeLong, E.F.; Hutchinson, C.R. Metagenomic analysis reveals diverse polyketide synthase gene clusters in microorganisms associated with the marine sponge Discodermia dissoluta. Appl. Environ. Microbiol. 2005, 71, 4840–4849. [Google Scholar] [PubMed]
- Siegl, A.; Kamke, J.; Hochmuth, T.; Piel, J.; Richter, M.; Liang, C.G.; Dandekar, T.; Hentschel, U. Single-cell genomics reveals the lifestyle of Poribacteria, a candidate phylum symbiotically associated with marine sponges. ISME J. 2011, 5, 61–70. [Google Scholar] [PubMed]
- Fisch, K.; Gurgui, C.; Heycke, N.; van der Sar, S.; Anderson, S.; Webb, V.; Taudien, S.; Platzer, M.; Rubio, B.; Robinson, S.; et al. Polyketide assembly lines of uncultivated sponge symbionts from structure-based gene targeting. Nat. Chem. Biol. 2009, 5, 494–501. [Google Scholar] [PubMed]
- Hochmuth, T.; Niederkruger, H.; Gernert, C.; Siegl, A.; Taudien, S.; Platzer, M.; Crews, P.; Hentschel, U.; Piel, J. Linking chemical and microbial diversity in marine sponges: Possible role for Poribacteria as producers of methyl-branched fatty acids. Chembiochem 2010, 11, 2572–2578. [Google Scholar] [PubMed]
- Osinga, R.; Tramper, J.; Wijffels, R.H. Cultivation of marine sponges. Mar. Biotechnol. 1999, 1, 509–532. [Google Scholar] [PubMed]
- Rinkevich, B. Marine invertebrate cell cultures: New millennium trends. Mar. Biotechnol. 2005, 7, 429–439. [Google Scholar] [PubMed]
- Sipkema, D.; Osinga, R.; Schatton, W.; Mendola, D.; Tramper, J.; Wijffels, R.H. Large-scale production of pharmaceuticals by marine sponges: Sea, cell, or synthesis? Biotechnol. Bioeng. 2005, 90, 201–222. [Google Scholar] [CrossRef]
- Wilkinson, C.R.; Vacelet, J. Transplantation of marine sponges to different conditions of light and current. J. Exp. Mar. Biol. Ecol. 1979, 37, 91–104. [Google Scholar]
- Duckworth, A.; Battershill, C.; Bergquist, P. Influence of explant procedures and environmental factors on culture success of three sponges. Aquaculture 1997, 156, 251–267. [Google Scholar]
- Van Treeck, P.; Eisinger, M.; Müller, J.; Paster, M.; Schuhmacher, H. Mariculture trials with Mediterranean sponge species—The exploitation of an old natural resource with sustainable and novel methods. Aquaculture 2003, 218, 439–455. [Google Scholar]
- De Voogd, N. An assessment of sponge mariculture potential in the Spermonde Archipelago, Indonesia. J. Mar. Biol. Assoc. UK 2007, 396, 1777–1784. [Google Scholar]
- De Rosa, S.; de Caro, S.; Tommonaro, G.; Slantchev, K.; Stefanov, K.; Popov, S. Development in a primary cell culture of the marine sponge Ircinia muscarum and analysis of the polar compounds. Mar. Biotechnol. 2001, 3, 281–286. [Google Scholar] [PubMed]
- De Rosa, S.; Tommonaro, G.; Slantchev, K.; Stefanov, K.; Popov, S. Lipophylic metabolites from the marine sponge Ircinia muscarum and its cell cultures. Mar. Biol. 2002, 140, 465–470. [Google Scholar]
- De Rosa, S.; de Caro, S.; Iodice, C.; Tommonaro, G.; Stefanov, K.; Popov, S. Development in primary cell culture of demosponges. J. Biotechnol. 2003, 100, 119–125. [Google Scholar] [PubMed]
- De Caralt, S.; Uriz, M.J.; Wijffels, R.H. Cell culture from sponges: Pluripotency and immortality. Trends Biotechnol. 2007, 25, 467–471. [Google Scholar] [PubMed]
- Piel, J.; Hui, D.Q.; Wen, G.P.; Butzke, D.; Platzer, M.; Fusetani, N.; Matsunaga, S. Antitumor polyketide biosynthesis by an uncultivated bacterial symbiont of the marine sponge Theonella swinhoei. Proc. Natl. Acad. Sci. USA 2004, 101, 16222–16227. [Google Scholar] [PubMed]
- Liu, M.; Fan, L.; Zhong, L.; Kjelleberg, S.; Thomas, T. Metaproteogenomic analysis of a community of sponge symbionts. ISME J. 2012, 6, 1515–1525. [Google Scholar] [PubMed]
- Kamke, J.; Sczyrba, A.; Ivanova, N.; Schwientek, P.; Rinke, C.; Mavromatis, K.; Woyke, T.; Hentschel, U. Single-cell genomics reveals complex carbohydrate degradation patterns in poribacterial symbionts of marine sponges. ISME J. 2013, 7, 2287–2300. [Google Scholar] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hardoim, C.C.P.; Costa, R. Microbial Communities and Bioactive Compounds in Marine Sponges of the Family Irciniidae—A Review. Mar. Drugs 2014, 12, 5089-5122. https://doi.org/10.3390/md12105089
Hardoim CCP, Costa R. Microbial Communities and Bioactive Compounds in Marine Sponges of the Family Irciniidae—A Review. Marine Drugs. 2014; 12(10):5089-5122. https://doi.org/10.3390/md12105089
Chicago/Turabian StyleHardoim, Cristiane C. P., and Rodrigo Costa. 2014. "Microbial Communities and Bioactive Compounds in Marine Sponges of the Family Irciniidae—A Review" Marine Drugs 12, no. 10: 5089-5122. https://doi.org/10.3390/md12105089
APA StyleHardoim, C. C. P., & Costa, R. (2014). Microbial Communities and Bioactive Compounds in Marine Sponges of the Family Irciniidae—A Review. Marine Drugs, 12(10), 5089-5122. https://doi.org/10.3390/md12105089