Vibrio vulnificus MO6-24/O Lipopolysaccharide Stimulates Superoxide Anion, Thromboxane B2, Matrix Metalloproteinase-9, Cytokine and Chemokine Release by Rat Brain Microglia in Vitro


 ,
, 
Abstract
:1. Introduction
2. Results
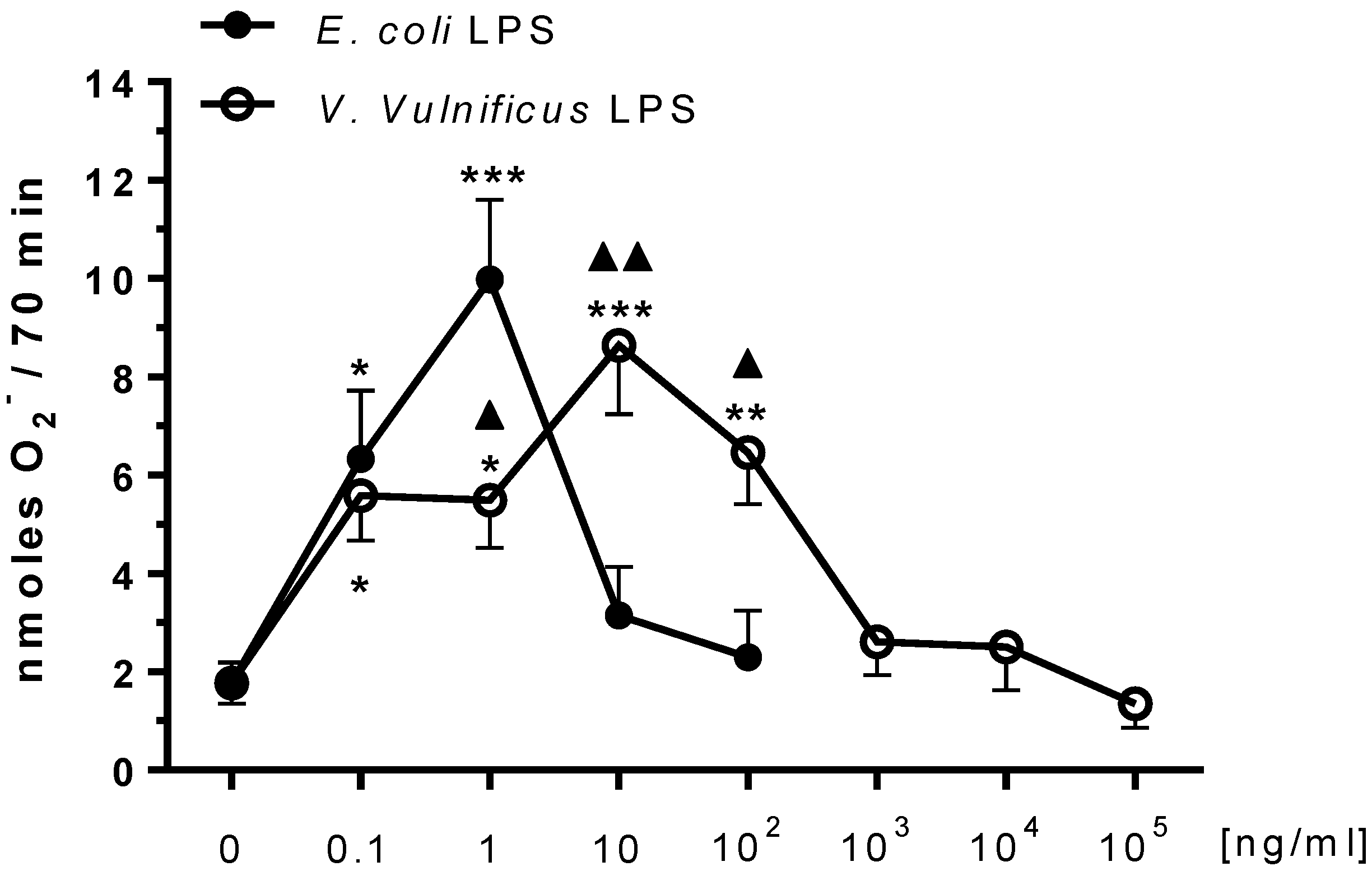
2.1. Effect of V. vulnificus LPS on Rat Brain Microglia O2− Generation

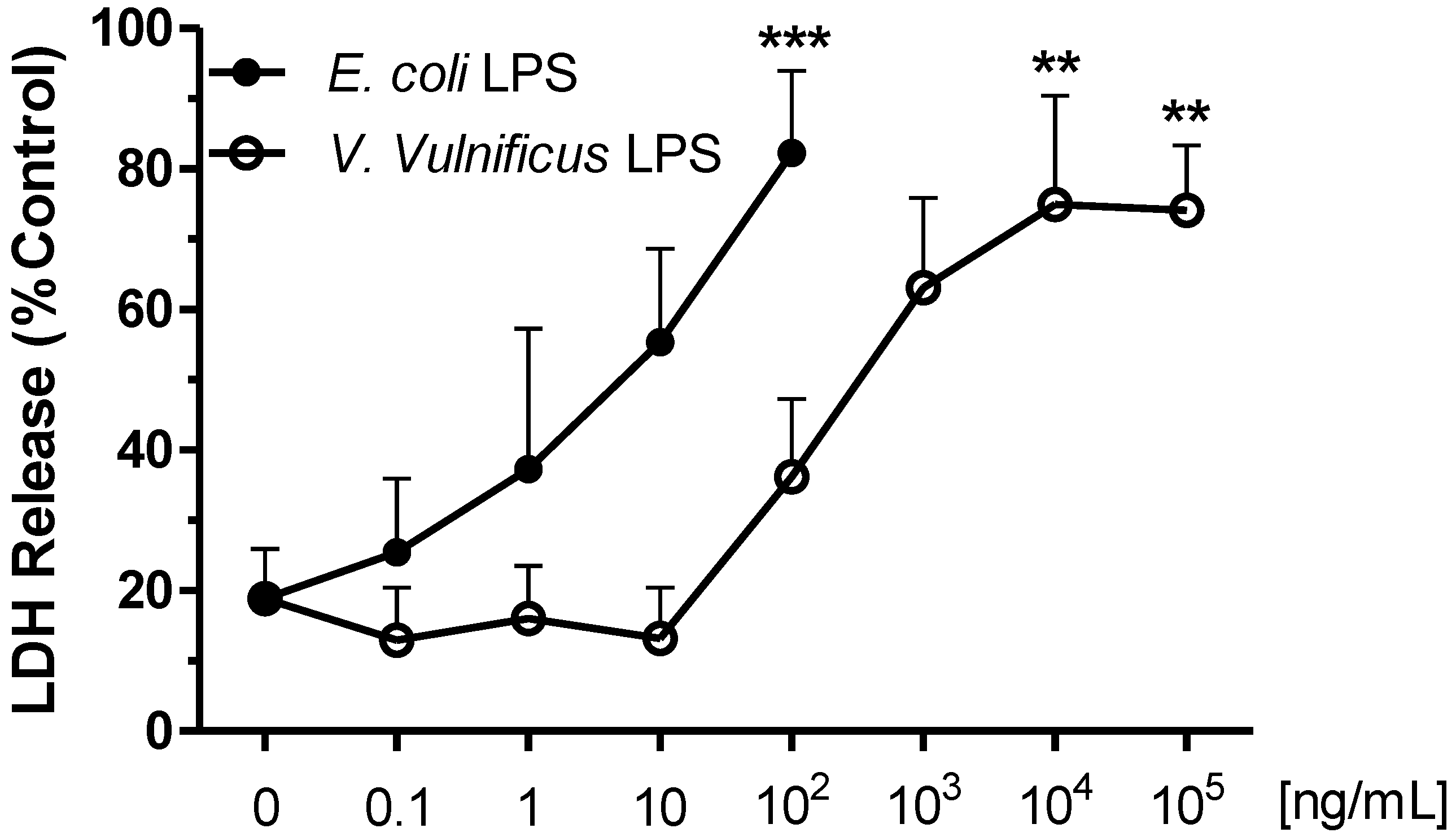
2.2. Effect of V. vulnificus LPS on Rat Brain Microglia LDH Generation

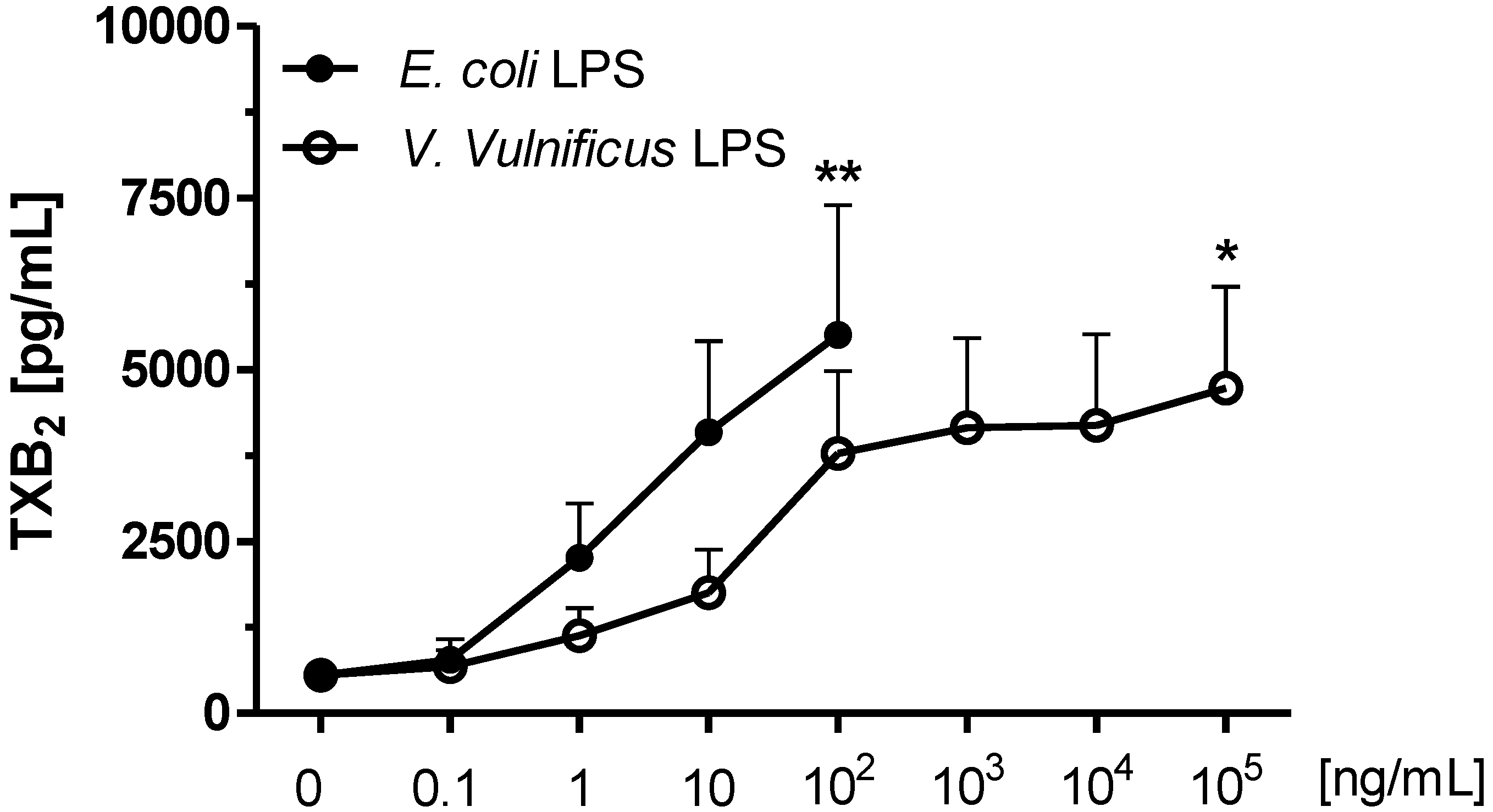
2.3. Effect of V. vulnificus LPS on Rat Brain Microglia TXB2 Generation

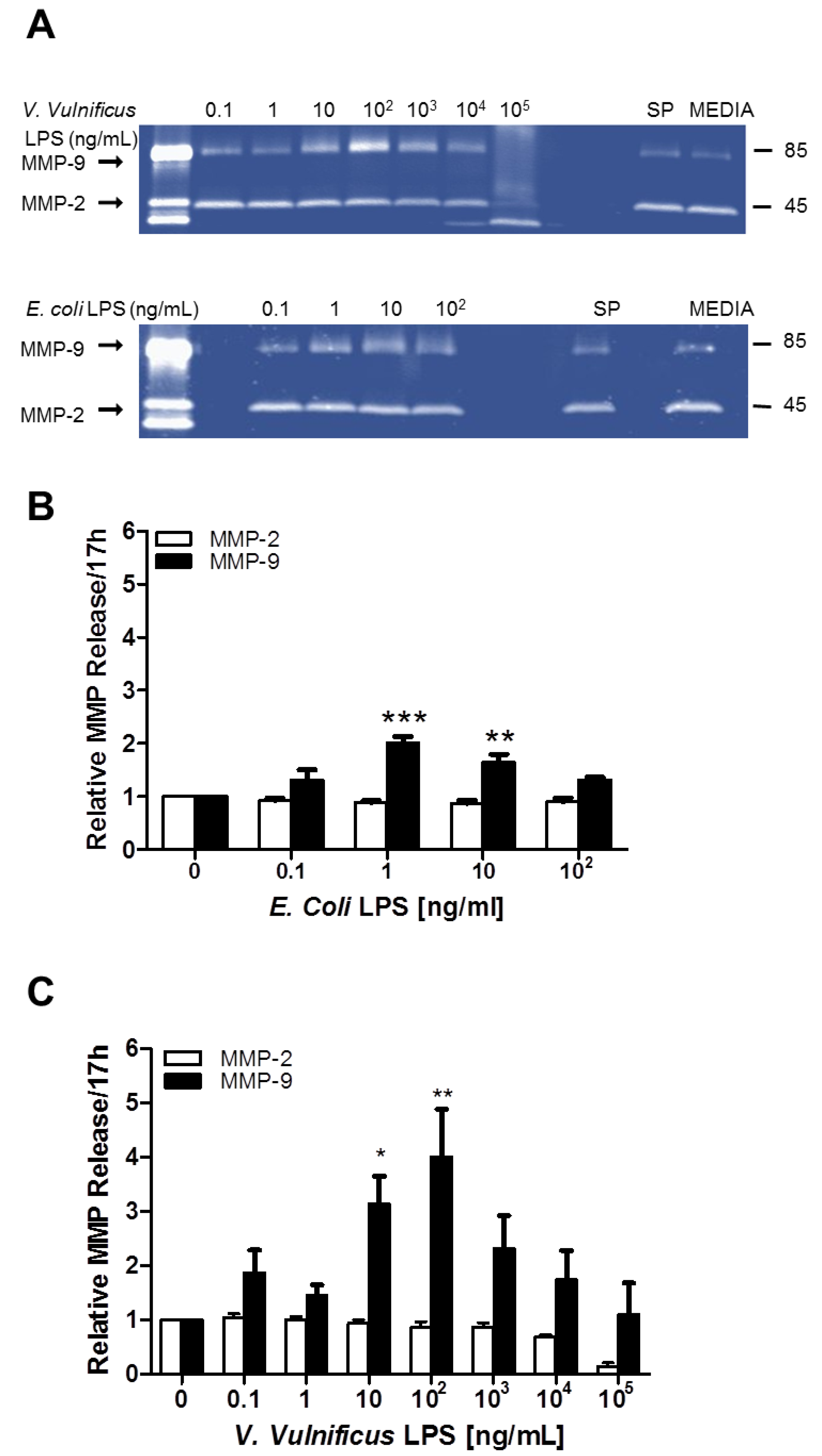
2.4. Effect of V. vulnificus LPS on Rat Brain Microglia MMP-2 and MMP-9 Generation

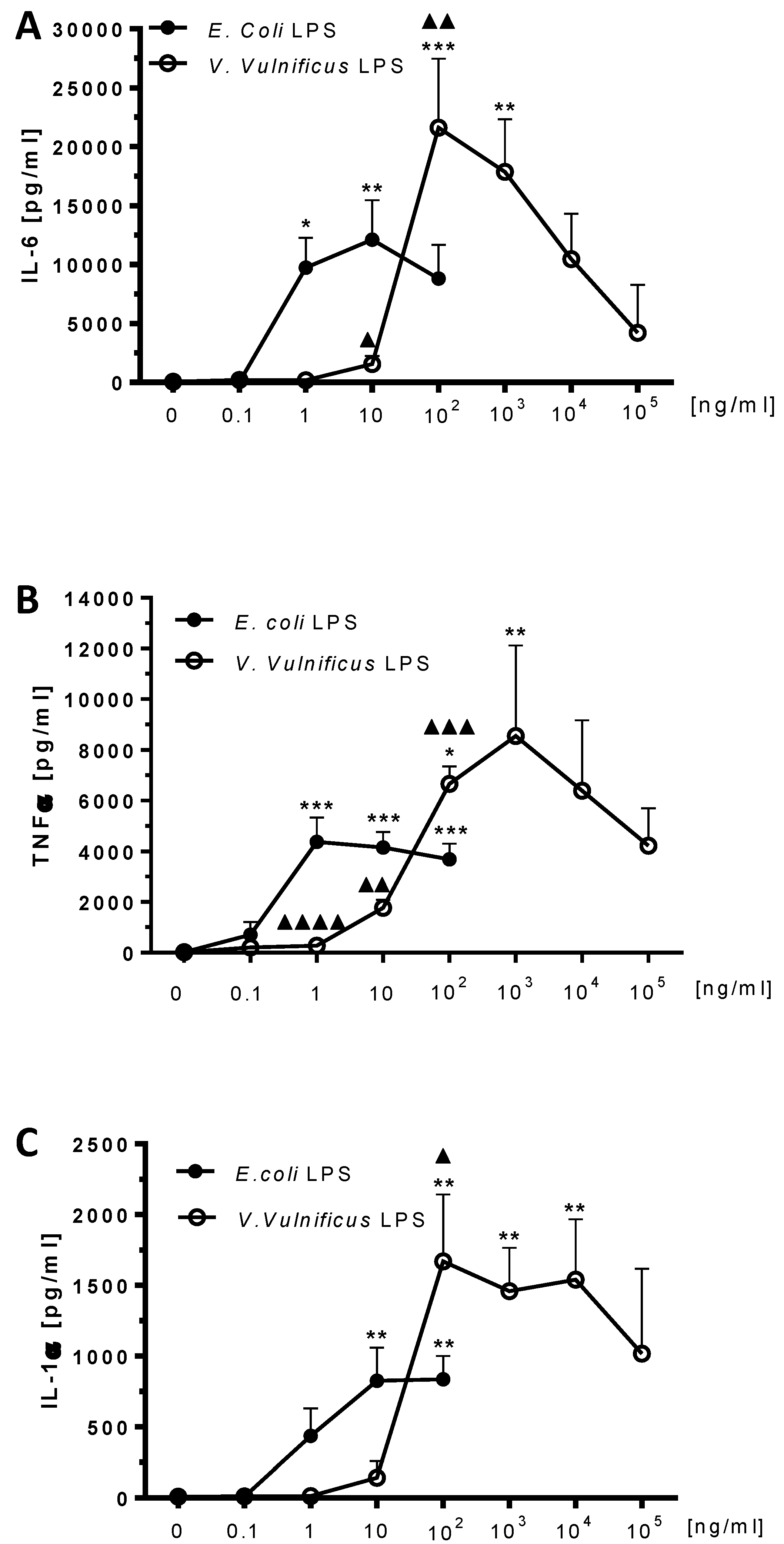
2.5. Effect of V. vulnificus LPS On Rat Brain Microglia Cytokines IL-6, TNF-α and IL-1α Release

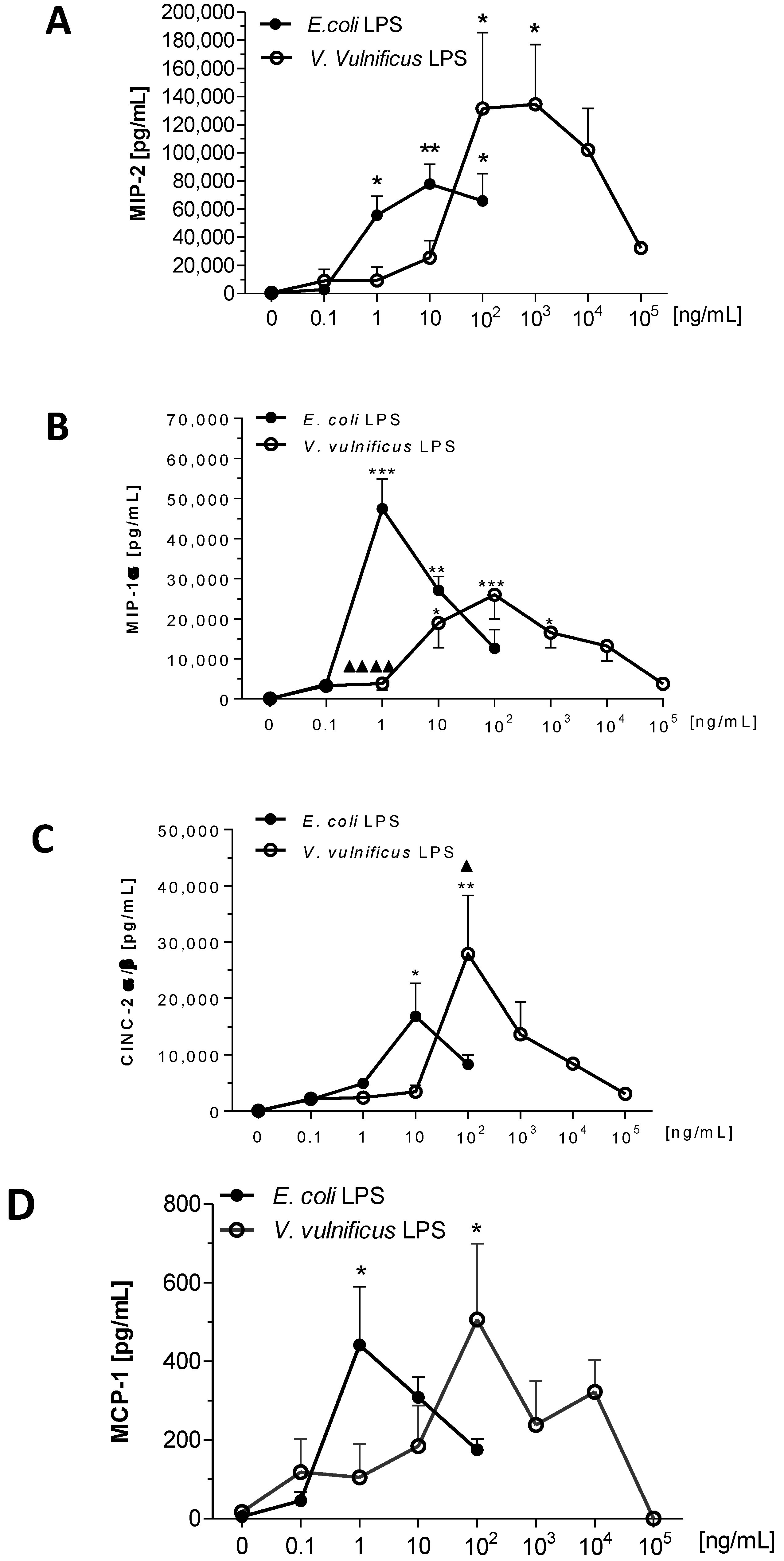
2.6. Effect of V. vulnificus LPS on Rat Brain Microglia MIP-2/CXCL2, MIP-1α/CCL3, CINC-2α/β (CXCL3) and MCP-1/CCL2 Generation

2.7. Effect of V. vulnificus LPS on Rat Brain Microglia TGF-β1 and BDNF

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TGF-β1 Release | BDNF Release | |||||||
|---|---|---|---|---|---|---|---|---|
| E. coli | V. vulnificus | E. coli | V. vulnificus | |||||
| (ng/mL) | (pg/mL) | n 1 | (pg/mL) | n 1 | (pg/mL) | n 1 | (pg/mL) | n 1 |
| 0 | 319.7 ± 50.1 | 3 | 319.7 ± 50.1 | 3 | 0 | 2 | 0 | 2 |
| 0.1 | 424.3 ± 40.3 | 3 | 386 ± 16.0 | 2 | 5.1 ± 4.4 | 2 | 1.4 ± 0.8 | 2 |
| 1 | 363.3 ± 36 | 3 | 419 ± 66.9 | 3 | 18.4 ± 14.9 | 2 | 10.2 ± 6.2 | 2 |
| 10 | 318.7 ± 73.2 | 3 | 358 ± 50.1 | 3 | 6.9 ± 3.9 | 2 | 6.5 ± 5.5 | 2 |
| 100 | 333.3 ± 58.6 | 3 | 310 ± 51.6 | 3 | 6.6 ± 3.0 | 2 | 9.1 ± 4.7 | 2 |
| 1000 | ND | 352 ± 55.5 | 3 | ND | 7.5 ± 7.5 | 2 | ||
| 10,000 | ND | 613 ± 58.7 | 3 | ND | 0 ± 0 | 2 | ||
| 100,000 | ND | 973.5 ± 264.5 * | 2 | ND | 2.05 ± 2.05 | 2 | ||
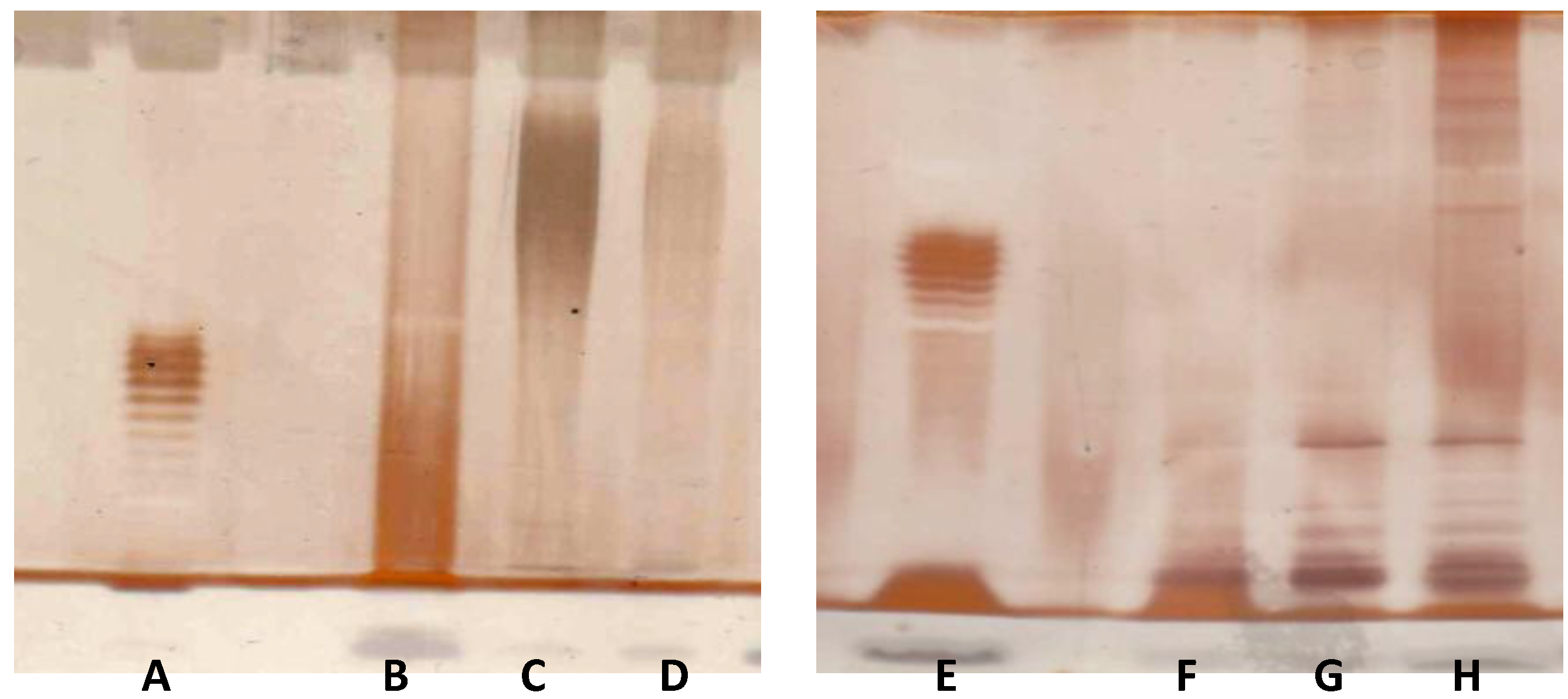
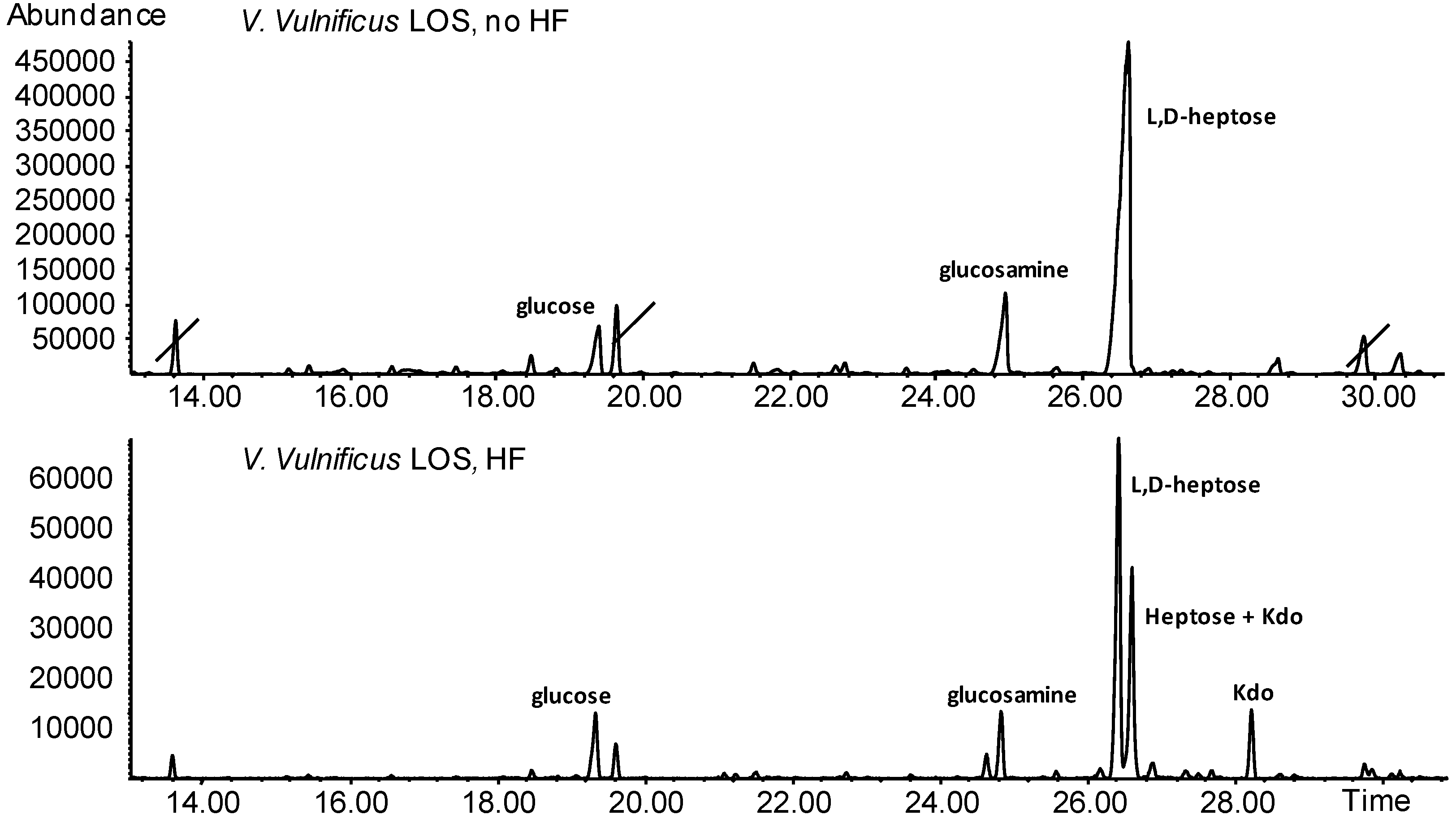
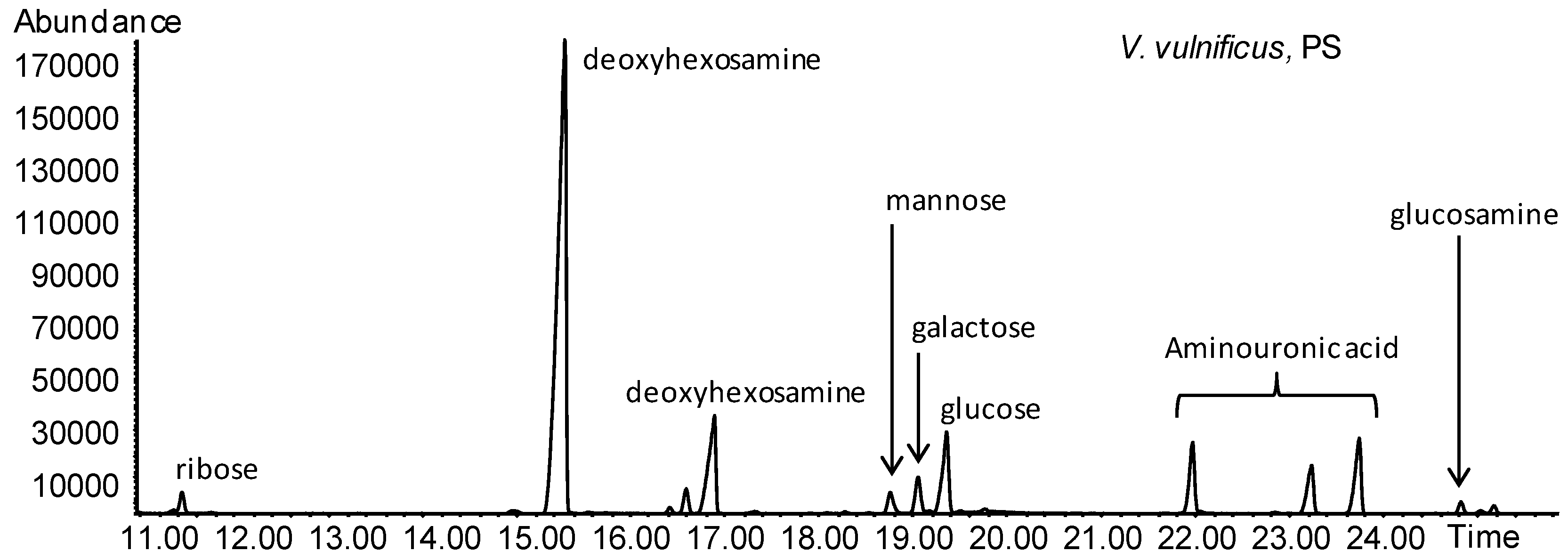
2.8. V. vulnificus LPS Isolation and Chemical Analyses



3. Discussion
4. Experimental Section
4.1. Reagents
4.2. LPS Contamination
4.3. Isolation of Rat Neonatal Microglia
4.4. Activation of Microglia with LPS (Experimental Design)
4.5. Assay for Superoxide Anion (O2−) Generation
4.6. Assay for Lactic Dehydrogenase (LDH)
4.7. Assay for Thromboxane B2 (TXB2) Generation
4.8. Gelatinase Zymography for MMP-2 and MMP-9 Analysis
4.9. Assays for Cytokines: TNF-α, IL-1α, IL-6 and TGF-β1
4.10. Assays for Chemokines: MIP-1α/CCL3, MIP-2/CXCL2, MCP-1/CCL2 and CINC-2α/β/CXCL3
4.11. Assay for the Neurotrophin Brain Derived Neurotrophic Factor (BDNF)
4.12. V. vulnificus LPS Chemical Analyses
4.13. Statistical Analysis
5. Conclusions
Abbreviations
| BDNF | brain derived neurotrophic factor |
| CINC-2α/β/CXCL3 | cytokine-induced neutrophil chemoattractant or macrophage inflammatory protein-2b or chemokine ligand 3 |
| DMEM | Dulbecco’s modified Eagle medium |
| E. coli | Escherichia coli |
| FBS | fetal bovine serum certified |
| FCC | ferricytochrome c type III |
| HBSS | Hank’s balanced salt solution |
| IL | interleukin |
| LDH | lactic dehydrogenase |
| LPS | lipopolysaccharide |
| MCP-1/CCL2 | monocyte chemotactic protein-1 or chemokine ligand 2 |
| MIP-1α/CCL3 | macrophage inflammatory protein 1α or chemokine ligand 3 |
| MIP-2/CXCL2 | macrophage inflammatory protein-2 |
| MMP-9 | matrix metalloproteinase-9 |
| O2− | superoxide anion |
| P | penicillin |
| PBS | phosphate buffered saline |
| PMA | phorbol 12-myristate 13-acetate |
| S | streptomycin |
| SEM | standard error of the mean |
| SOD | superoxide dismutase |
| TGF-β1 | transforming growth factor β1 |
| TNF-α | tumor necrosis factor |
| TXB2 | thromboxane B2 |
| V. vulnificus | Vibrio vulnificus |
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cazorla, C.; Guigon, A.; Noel, M.; Quilici, M.L.; Lacassin, F. Fatal Vibrio vulnificus infection associated with eating raw oysters, New Caledonia. Emerg. Infect. Dis. 2011, 17, 136–137. [Google Scholar] [CrossRef]
- Oliver, J.D. Wound infections caused by Vibrio vulnificus and other marine bacteria. Epidemiol. Infect. 2005, 133, 383–391. [Google Scholar] [CrossRef]
- Horseman, M.A.; Surani, S. A comprehensive review of Vibrio vulnificus: An important cause of severe sepsis and skin and soft-tissue infection. Int. J. Infect. Dis. 2011, 15, e157–e166. [Google Scholar] [CrossRef]
- Kim, S.J.; Kim, B.C.; Kim, D.C.; Kim, M.K.; Cho, K.H.; Seo, J.J.; Shin, J.H. A fatal case of Vibrio vulnificus meningoencephalitis. Clin. Microbiol. Infect. 2003, 9, 568–571. [Google Scholar] [CrossRef]
- Katz, B.Z. Vibrio vulnificus meningitis in a boy with thalassemia after eating raw oysters. Pediatrics 1988, 82, 784–786. [Google Scholar]
- Baker-Austin, C.; McArthur, J.V.; Lindell, A.H.; Wright, M.S.; Tuckfield, R.C.; Gooch, J.; Warner, L.; Oliver, J.; Stepanauskas, R. Multi-site analysis reveals widespread antibiotic resistance in the marine pathogen Vibrio vulnificus. Microb. Ecol. 2009, 57, 151–159. [Google Scholar]
- Broza, Y.Y.; Raz, N.; Lerner, L.; Danin-Poleg, Y.; Kashi, Y. Genetic diversity of the human pathogen Vibrio vulnificus: A new phylogroup. Int. J. Food Microbiol. 2012, 153, 436–443. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Lemm, E.; Hartnell, R.; Lowther, J.; Onley, R.; Amaro, C.; Oliver, J.D.; Lees, D. pilF polymorphism-based real-time PCR to distinguish Vibrio vulnificus strains of human health relevance. Food Microbiol. 2012, 30, 17–23. [Google Scholar] [CrossRef]
- Bahrani, K.; Oliver, J.D. Studies on the lipopolysaccharide of a virulent and an avirulent strain of Vibrio vulnificus. Biochem. Cell Biol. 1990, 68, 547–551. [Google Scholar] [CrossRef]
- Biosca, E.G.; Collado, R.M.; Oliver, J.D.; Amaro, C. Comparative study of biological properties and electrophoretic characteristics of lipopolysaccharide from eel-virulent and eel-A virulent Vibrio vulnificus strains. Appl. Environ. Microbiol. 1999, 65, 856–858. [Google Scholar]
- Vinogradov, E.; Wilde, C.; Anderson, E.M.; Nakhamchik, A.; Lam, J.S.; Rowe-Magnus, D.A. Structure of the lipopolysaccharide core of Vibrio vulnificus type strain 27562. Carbohydr. Res. 2009, 344, 484–490. [Google Scholar] [CrossRef]
- McPherson, V.L.; Watts, J.A.; Simpson, L.M.; Oliver, J.D. Physiological effects of the lipopolysaccharide of Vibrio vulnificus on mice and rats. Microbios 1991, 67, 141–149. [Google Scholar]
- Chatzidaki-Livanis, M.; Hubbard, M.A.; Gordon, K.; Harwood, V.J.; Wright, A.C. Genetic distinctions among clinical and environmental strains of Vibrio vulnificus. Appl. Environ. Microbiol. 2006, 72, 6136–6141. [Google Scholar] [CrossRef]
- Chae, M.R.; Park, B.H.; Kim, J.S.; Rho, H.W.; Park, J.W.; Kim, H.R. Protective effect of C-reactive protein against the lethality induced by Vibrio vulnificus lipopolysaccharide. Microbiol. Immunol. 2000, 44, 335–340. [Google Scholar] [CrossRef]
- Park, K.H.; Kim, J.S.; Lee, Y.R.; Moon, Y.J.; Hur, H.; Choi, Y.H.; Kim, C.H.; Kim, U.H.; Song, E.K.; Yoo, W.H.; et al. Low-density lipoprotein protects Vibrio vulnificus-induced lethality through blocking lipopolysaccharide action. Exp. Mol. Med. 2007, 39, 673–678. [Google Scholar] [CrossRef]
- Merkel, S.M.; Alexander, S.; Zufall, E.; Oliver, J.D.; Huet-Hudson, Y.M. Essential role for estrogen in protection against Vibrio vulnificus-induced endotoxic shock. Infect. Immun. 2001, 69, 6119–6122. [Google Scholar] [CrossRef]
- Jones, M.K.; Oliver, J.D. Vibrio vulnificus: Disease and pathogenesis. Infect. Immun. 2009, 77, 1723–1733. [Google Scholar] [CrossRef]
- Starks, A.M.; Schoeb, T.R.; Tamplin, M.L.; Parveen, S.; Doyle, T.J.; Bomeisl, P.E.; Escudero, G.M.; Gulig, P.A. Pathogenesis of infection by clinical and environmental strains of Vibrio vulnificus in iron-dextran-treated mice. Infect. Immun. 2000, 68, 5785–5793. [Google Scholar] [CrossRef]
- Powell, J.L.; Wright, A.C.; Wasserman, S.S.; Hone, D.M.; Morris, J.G., Jr. Release of tumor necrosis factor alpha in response to Vibrio vulnificus capsular polysaccharide in in vivo and in vitro models. Infect. Immun. 1997, 65, 3713–3718. [Google Scholar]
- Cunningham, C.; Wilcockson, D.C.; Campion, S.; Lunnon, K.; Perry, V.H. Central and systemic endotoxin challenges exacerbate the local inflammatory response and increase neuronal death during chronic neurodegeneration. J. Neurosci. 2005, 25, 9275–9284. [Google Scholar] [CrossRef]
- Opal, S.M. Endotoxins and other sepsis triggers. Contrib. Nephrol. 2010, 167, 14–24. [Google Scholar] [CrossRef]
- Teeling, J.L.; Perry, V.H. Systemic infection and inflammation in acute CNS injury and chronic neurodegeneration: Underlying mechanisms. Neuroscience 2009, 158, 1062–1073. [Google Scholar] [CrossRef]
- Banks, W.A.; Erickson, M.A. The blood-brain barrier and immune function and dysfunction. Neurobiol. Dis. 2010, 37, 26–32. [Google Scholar] [CrossRef]
- Hannestad, J.; Gallezot, J.D.; Schafbauer, T.; Lim, K.; Kloczynski, T.; Morris, E.D.; Carson, R.E.; Ding, Y.S.; Cosgrove, K. Endotoxin-induced systemic inflammation activates microglia: [11C]PBR28 positron emission tomography in nonhuman primates. Neuroimage 2012, 63, 232–239. [Google Scholar] [CrossRef]
- Rock, R.B.; Gekker, G.; Hu, S.; Sheng, W.S.; Cheeran, M.; Lokensgard, J.R.; Peterson, P.K. Role of microglia in central nervous system infections. Clin. Microbiol. Rev. 2004, 17, 942–964. [Google Scholar] [CrossRef]
- Mayer, A.M. Therapeutic implications of microglia activation by lipopolysaccharide and reactive oxygen species generation in septic shock and central nervous system pathologies: A review. Medicina 1998, 58, 377–385. [Google Scholar]
- Mayer, A.M.; Oh, S.; Ramsey, K.H.; Jacobson, P.B.; Glaser, K.B.; Romanic, A.M. Escherichia coli lipopolysaccharide potentiation and inhibition of rat neonatal microglia superoxide anion generation: Correlation with prior lactic dehydrogenase, nitric oxide, tumor necrosis factor-alpha, thromboxane B2, and metalloprotease release. Shock 1999, 11, 180–186. [Google Scholar] [CrossRef]
- Banati, R.B.; Gehrmann, J.; Schubert, P.; Kreutzberg, G.W. Cytotoxicity of microglia. Glia 1993, 7, 111–118. [Google Scholar] [CrossRef]
- Qin, L.; Wu, X.; Block, M.L.; Liu, Y.; Breese, G.R.; Hong, J.S.; Knapp, D.J.; Crews, F.T. Systemic LPS causes chronic neuroinflammation and progressive neurodegeneration. Glia 2007, 55, 453–462. [Google Scholar] [CrossRef]
- Perry, V.H.; Nicoll, J.A.; Holmes, C. Microglia in neurodegenerative disease. Nat. Rev. Neurol. 2010, 6, 193–201. [Google Scholar] [CrossRef]
- Halliwell, B. Reactive oxygen species and the central nervous system. J. Neurochem. 1992, 59, 1609–1623. [Google Scholar] [CrossRef]
- Mayer, A.M.S.; Hall, M.L.; Jacobson, P.B.; Powell, J.L. Differential effect of Vibrio vulnificus and Escherichia coli LPS on rat brain neonatal microglia release of thromboxane B2 and tumor necrosis factor α. FASEB J. 2007, 21. 560.9; Abstract 404. [Google Scholar]
- Frenkel, J.; Aldulescu, M.; Hall, M.; Powell, J.L.; Glaser, K.B.; Mayer, A.M.S. Effect of the marine Vibrio vulnificus lipopolysaccharide on rat brain microglia cytokine and chemokine release. Toxicologist 2009, 108, 12, Abstract 64. [Google Scholar]
- Ottenhoff, L.D.; Patel, N.; Hall, M.L.; Glaser, K.B.; Jacobson, P.; Rowley, D.; De Castro, C.; Molinaro, A.; Mayer, A.M.S. Effect of Vibrio vulnificus lipopolysaccharide (LPS) on rat brain microglia: Cytokine and chemokine expression profiling by antibody array technology. Toxicologist 2011, 120, 148, Abstract 688. [Google Scholar]
- Colton, C.A.; Wilcock, D.M. Assessing activation states in microglia. CNS Neurol. Disord. Drug Targets 2010, 9, 174–191. [Google Scholar] [CrossRef]
- Block, M.L.; Zecca, L.; Hong, J.S. Microglia-mediated neurotoxicity: Uncovering the molecular mechanisms. Nat. Rev. Neurosci. 2007, 8, 57–69. [Google Scholar] [CrossRef]
- Colton, C.A.; Gilbert, D.L. Production of superoxide anions by a CNS macrophage, the microglia. FEBS Lett. 1987, 223, 284–288. [Google Scholar] [CrossRef]
- Mayer, A.M.; Clifford, J.A.; Aldulescu, M.; Frenkel, J.A.; Holland, M.A.; Hall, M.L.; Glaser, K.B.; Berry, J. Cyanobacterial Microcystis aeruginosa lipopolysaccharide elicits release of superoxide anion, thromboxane B, cytokines, chemokines, and matrix metalloproteinase-9 by rat microglia. Toxicol. Sci. 2011, 121, 63–72. [Google Scholar] [CrossRef]
- Morgenstern, S.; Flor, R.; Kessler, G.; Klein, B. Automated determination of NAD-coupled enzymes. Determination of lactic dehydrogenase. Anal. Biochem. 1966, 13, 149–161. [Google Scholar] [CrossRef]
- Minghetti, L.; Levi, G. Induction of prostanoid biosynthesis by bacterial lipopolysaccharide and isoproterenol in rat microglial cultures. J. Neurochem. 1995, 65, 2690–2698. [Google Scholar] [CrossRef]
- Vanlaere, I.; Libert, C. Matrix metalloproteinases as drug targets in infections caused by gram-negative bacteria and in septic shock. Clin. Microbiol. Rev. 2009, 22, 224–239. [Google Scholar] [CrossRef]
- Candelario-Jalil, E.; Yang, Y.; Rosenberg, G.A. Diverse roles of matrix metalloproteinases and tissue inhibitors of metalloproteinases in neuroinflammation and cerebral ischemia. Neuroscience 2009, 158, 983–994. [Google Scholar] [CrossRef]
- Gottschall, P.E.; Yu, X.; Bing, B. Increased production of gelatinase B (matrix metalloproteinase-9) and interleukin-6 by activated rat microglia in culture. J. Neurosci. Res. 1995, 42, 335–342. [Google Scholar] [CrossRef]
- Nishimoto, N. Interleukin-6 as a therapeutic target in candidate inflammatory diseases. Clin. Pharmacol. Ther. 2010, 87, 483–487. [Google Scholar] [CrossRef]
- Maccioni, R.B.; Rojo, L.E.; Fernandez, J.A.; Kuljis, R.O. The role of neuroimmunomodulation in Alzheimer’s disease. Ann. N. Y. Acad. Sci. 2009, 1153, 240–246. [Google Scholar]
- Lee, S.C.; Liu, W.; Dickson, D.W.; Brosnan, C.F.; Berman, J.W. Cytokine production by human fetal microglia and astrocytes. Differential induction by lipopolysaccharide and IL-1 beta. J. Immunol. 1993, 150, 2659–2667. [Google Scholar]
- Hausler, K.G.; Prinz, M.; Nolte, C.; Weber, J.R.; Schumann, R.R.; Kettenmann, H.; Hanisch, U.K. Interferon-gamma differentially modulates the release of cytokines and chemokines in lipopolysaccharide- and pneumococcal cell wall-stimulated mouse microglia and macrophages. Eur. J. Neurosci. 2002, 16, 2113–2122. [Google Scholar] [CrossRef]
- Horvath, R.J.; Nutile-McMenemy, N.; Alkaitis, M.S.; Deleo, J.A. Differential migration, LPS-induced cytokine, chemokine, and NO expression in immortalized BV-2 and HAPI cell lines and primary microglial cultures. J. Neurochem. 2008, 107, 557–569. [Google Scholar] [CrossRef]
- McCoy, M.K.; Tansey, M.G. TNF signaling inhibition in the CNS: Implications for normal brain function and neurodegenerative disease. J. Neuroinflamm. 2008, 5, 5–45. [Google Scholar] [CrossRef]
- Hayashi, M.; Luo, Y.; Laning, J.; Strieter, R.M.; Dorf, M.E. Production and function of monocyte chemoattractant protein-1 and other beta-chemokines in murine glial cells. J. Neuroimmunol. 1995, 60, 143–150. [Google Scholar] [CrossRef]
- Simi, A.; Tsakiri, N.; Wang, P.; Rothwell, N.J. Interleukin-1 and inflammatory neurodegeneration. Biochem. Soc. Trans. 2007, 35, 1122–1126. [Google Scholar] [CrossRef]
- Allan, S.M.; Tyrrell, P.J.; Rothwell, N.J. Interleukin-1 and neuronal injury. Nat. Rev. Immunol. 2005, 5, 629–640. [Google Scholar] [CrossRef]
- Giulian, D.; Baker, T.J.; Shih, L.C.; Lachman, L.B. Interleukin 1 of the central nervous system is produced by ameboid microglia. J. Exp. Med. 1986, 164, 594–604. [Google Scholar] [CrossRef]
- Ubogu, E.E.; Cossoy, M.B.; Ransohoff, R.M. The expression and function of chemokines involved in CNS inflammation. Trends Pharmacol. Sci. 2006, 27, 48–55. [Google Scholar] [CrossRef]
- Esen, N.; Kielian, T. Effects of low dose GM-CSF on microglial inflammatory profiles to diverse pathogen-associated molecular patterns (PAMPs). J. Neuroinflamm. 2007, 4, 10. [Google Scholar] [CrossRef]
- Lafrance, V.; Inoue, W.; Kan, B.; Luheshi, G.N. Leptin modulates cell morphology and cytokine release in microglia. Brain Behav. Immun. 2010, 24, 358–365. [Google Scholar] [CrossRef]
- Szczucinski, A.; Losy, J. Chemokines and chemokine receptors in multiple sclerosis. Potential targets for new therapies. Acta Neurol. Scand. 2007, 115, 137–146. [Google Scholar] [CrossRef]
- Sun, D.; Hu, X.; Liu, X.; Whitaker, J.N.; Walker, W.S. Expression of chemokine genes in rat glial cells: The effect of myelin basic protein-reactive encephalitogenic T cells. J. Neurosci. Res. 1997, 48, 192–200. [Google Scholar] [CrossRef]
- Peterson, P.K.; Hu, S.; Salak-Johnson, J.; Molitor, T.W.; Chao, C.C. Differential production of and migratory response to beta chemokines by human microglia and astrocytes. J. Infect. Dis. 1997, 175, 478–481. [Google Scholar] [CrossRef]
- Bruyere, C.; Mijatovic, T.; Lonez, C.; Spiegl-Kreinecker, S.; Berger, W.; Kast, R.E.; Ruysschaert, J.M.; Kiss, R.; Lefranc, F. Temozolomide-induced modification of the CXC chemokine network in experimental gliomas. Int. J. Oncol. 2011, 38, 1453–1464. [Google Scholar]
- Janabi, N.; Hau, I.; Tardieu, M. Negative feedback between prostaglandin and alpha- and beta-chemokine synthesis in human microglial cells and astrocytes. J. Immunol. 1999, 162, 1701–1706. [Google Scholar]
- Qian, L.; Wei, S.J.; Zhang, D.; Hu, X.; Xu, Z.; Wilson, B.; El-Benna, J.; Hong, J.S.; Flood, P.M. Potent anti-inflammatory and neuroprotective effects of TGF-beta1 are mediated through the inhibition of ERK and p47phox-Ser345 phosphorylation and translocation in microglia. J. Immunol. 2008, 181, 660–668. [Google Scholar]
- Polazzi, E.; Altamira, L.E.; Eleuteri, S.; Barbaro, R.; Casadio, C.; Contestabile, A.; Monti, B. Neuroprotection of microglial conditioned medium on 6-hydroxydopamine-induced neuronal death: Role of transforming growth factor beta-2. J. Neurochem. 2009, 110, 545–556. [Google Scholar] [CrossRef]
- Walker, D.G.; Kim, S.U.; McGeer, P.L. Complement and cytokine gene expression in cultured microglial derived from postmortem human brains. J. Neurosci. Res. 1995, 40, 478–493. [Google Scholar] [CrossRef]
- Chao, C.C.; Hu, S.; Sheng, W.S.; Tsang, M.; Peterson, P.K. Tumor necrosis factor-alpha mediates the release of bioactive transforming growth factor-beta in murine microglial cell cultures. Clin. Immunol. Immunopathol. 1995, 77, 358–365. [Google Scholar] [CrossRef]
- De Castro, C.; Parrilli, M.; Holst, O.; Molinaro, A. Microbe-associated molecular patterns in innate immunity: Extraction and chemical analysis of gram-negative bacterial lipopolysaccharides. Methods Enzymol. 2010, 480, 89–115. [Google Scholar] [CrossRef]
- Ransohoff, R.M.; Perry, V.H. Microglial physiology: Unique stimuli, specialized responses. Annu. Rev. Immunol. 2009, 27, 119–145. [Google Scholar] [CrossRef]
- Holst, O.; Ulmer, A.J.; Brade, H.; Flad, H.D.; Rietschel, E.T. Biochemistry and cell biology of bacterial endotoxins. FEMS Immunol. Med. Microbiol. 1996, 16, 83–104. [Google Scholar] [CrossRef]
- Slepko, N.; Minghetti, L.; Polazzi, E.; Nicolini, A.; Levi, G. Reorientation of prostanoid production accompanies “activation” of adult microglial cells in culture. J. Neurosci. Res. 1997, 49, 292–300. [Google Scholar] [CrossRef]
- Mayer, A.M.S.; Hall, M.L.; Walker, D.L. Inhibition of LPS-primed human brain microglia superoxide and thromboxane B2 generation by the marine Manzamines. Inflamm. Res. 2004, 53, 217. [Google Scholar] [CrossRef]
- Powell, J.L.; Strauss, K.A.; Wiley, C.; Zhan, M.; Morris, J.G., Jr. Inflammatory cytokine response to Vibrio vulnificus elicited by peripheral blood mononuclear cells from chronic alcohol users is associated with biomarkers of cellular oxidative stress. Infect. Immun. 2003, 71, 4212–4216. [Google Scholar] [CrossRef]
- Sharma, S.K. Endotoxin detection and elimination in biotechnology. Biotechnol. Appl. Biochem. 1986, 8, 5–22. [Google Scholar]
- Galanos, C.; Luderitz, O.; Westphal, O. A new method for the extraction of R lipopolysaccharides. Eur. J. Biochem. 1969, 9, 245–249. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mayer, A.M.S.; Hall, M.L.; Holland, M.; De Castro, C.; Molinaro, A.; Aldulescu, M.; Frenkel, J.; Ottenhoff, L.; Rowley, D.; Powell, J. Vibrio vulnificus MO6-24/O Lipopolysaccharide Stimulates Superoxide Anion, Thromboxane B2, Matrix Metalloproteinase-9, Cytokine and Chemokine Release by Rat Brain Microglia in Vitro. Mar. Drugs 2014, 12, 1732-1756. https://doi.org/10.3390/md12041732
Mayer AMS, Hall ML, Holland M, De Castro C, Molinaro A, Aldulescu M, Frenkel J, Ottenhoff L, Rowley D, Powell J. Vibrio vulnificus MO6-24/O Lipopolysaccharide Stimulates Superoxide Anion, Thromboxane B2, Matrix Metalloproteinase-9, Cytokine and Chemokine Release by Rat Brain Microglia in Vitro. Marine Drugs. 2014; 12(4):1732-1756. https://doi.org/10.3390/md12041732
Chicago/Turabian StyleMayer, Alejandro M. S., Mary L. Hall, Michael Holland, Cristina De Castro, Antonio Molinaro, Monica Aldulescu, Jeffrey Frenkel, Lauren Ottenhoff, David Rowley, and Jan Powell. 2014. "Vibrio vulnificus MO6-24/O Lipopolysaccharide Stimulates Superoxide Anion, Thromboxane B2, Matrix Metalloproteinase-9, Cytokine and Chemokine Release by Rat Brain Microglia in Vitro" Marine Drugs 12, no. 4: 1732-1756. https://doi.org/10.3390/md12041732
APA StyleMayer, A. M. S., Hall, M. L., Holland, M., De Castro, C., Molinaro, A., Aldulescu, M., Frenkel, J., Ottenhoff, L., Rowley, D., & Powell, J. (2014). Vibrio vulnificus MO6-24/O Lipopolysaccharide Stimulates Superoxide Anion, Thromboxane B2, Matrix Metalloproteinase-9, Cytokine and Chemokine Release by Rat Brain Microglia in Vitro. Marine Drugs, 12(4), 1732-1756. https://doi.org/10.3390/md12041732