
Seaweed Polysaccharides (Laminarin and Fucoidan) as Functional Ingredients in Pork Meat: An Evaluation of Anti-Oxidative Potential, Thermal Stability and Bioaccessibility

Abstract
:
1. Introduction
2. Results and Discussion
2.1. Free Radical Scavenging Activity of Seaweed Polysaccharides (L, F and L/F)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Incubate | Time, h | ||
|---|---|---|---|
| 1 | 4 | 20 | |
| L1 * | 1.09 ± 0.92 ab | 1.39 ± 0.96 ab | 1.64 ± 1.30 ab |
| L10 | 1.55 ± 1.21 b | 2.72 ± 1.77 b | 3.16 ± 2.72 b |
| F1 | 66.13 ± 0.32 c | 76.48 ± 0.30 c | 90.68 ± 0.55 c |
| L/F1 | 35.43 ± 2.04 d | 47.35 ± 1.79 d | 69.51 ± 1.37 d |
| L/F3 | 56.18 ± 1.01 e | 68.40 ± 0.89 e | 78.41 ± 0.21 e |
| Trolox | 95.89 ± 0.08 f | 95.92 ± 0.14 f | 95.76 ± 0.48 f |
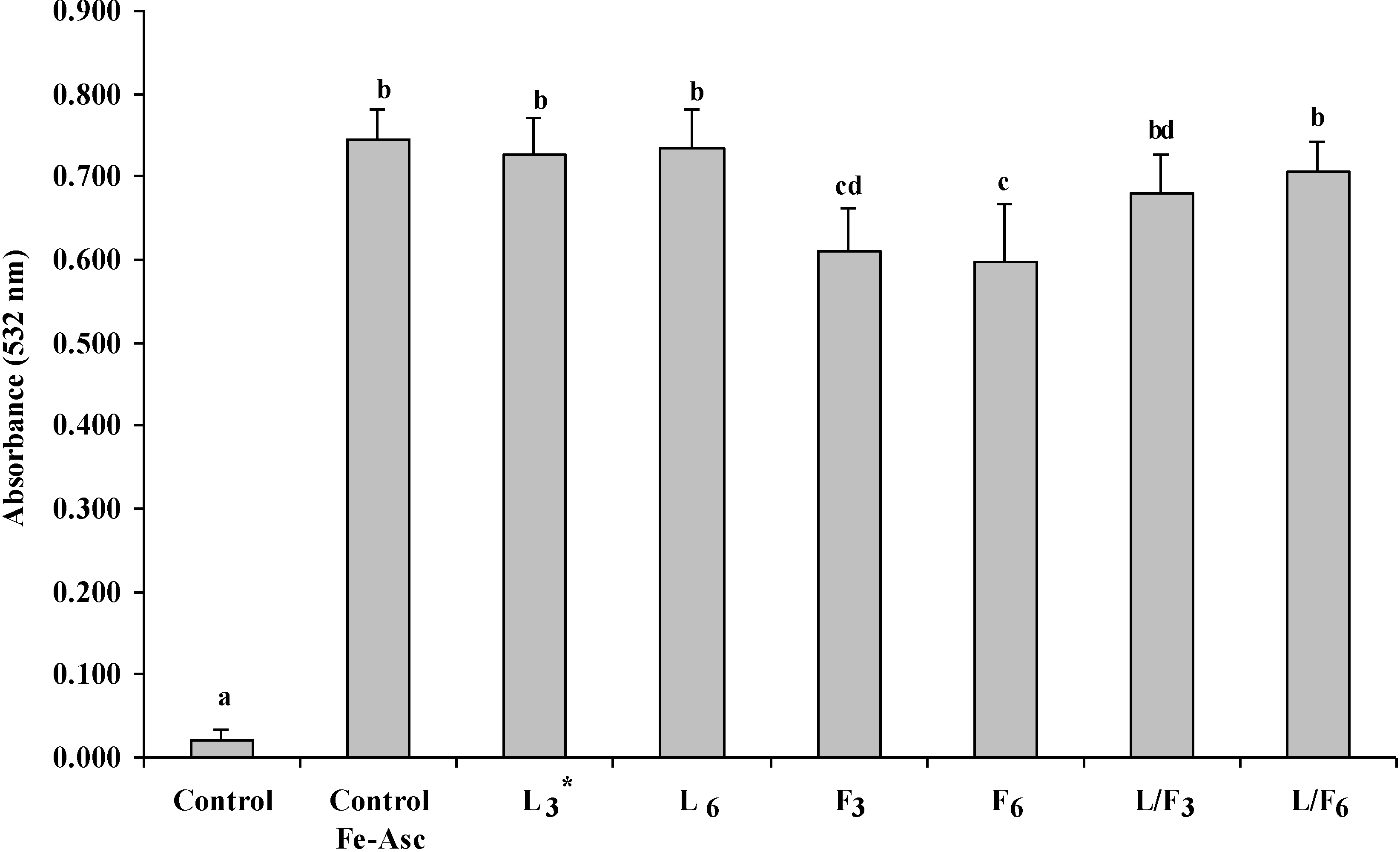
2.2. Effect of Seaweed Polysaccharides on Lipid Oxidation in Pork Muscle Model Systems

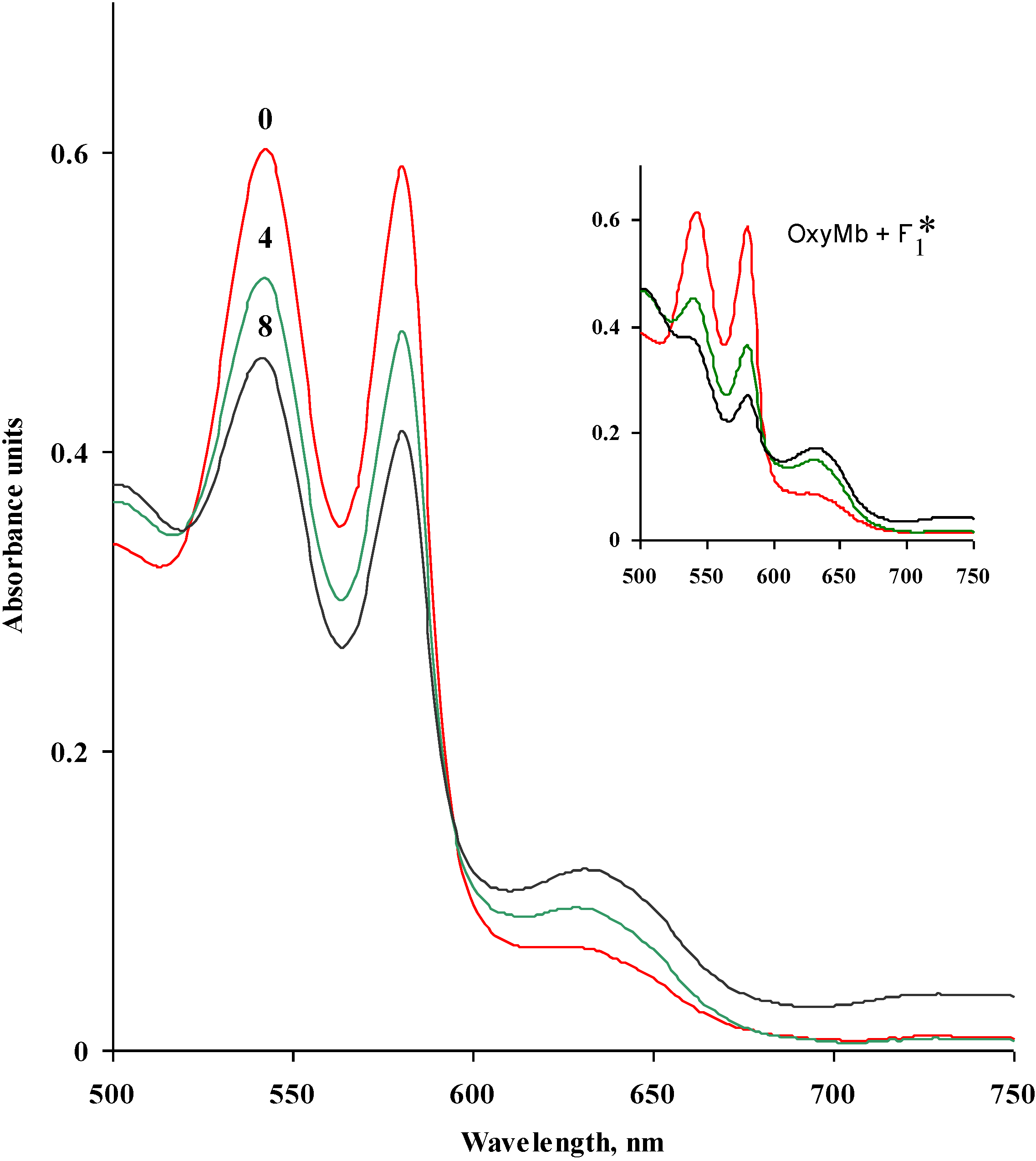
2.3. Effect of Seaweed Polysaccharides on Oxymyoglobin Oxidation
| Incubate | time, d | ||
|---|---|---|---|
| 0 | 4 | 8 | |
| Control | 76.53 ± 2.28 a | 59.92 ± 2.30 a | 54.46 ± 2.02 a |
| L0.1 * | 76.57 ± 2.31 a | 59.68 ± 2.14 ab | 54.00 ± 2.50 a |
| L1 | 76.59 ± 2.73 a | 58.11 ± 3.12 abc | 52.51 ± 2.75 a |
| F0.1 | 74.73 ± 2.54 ac | 53.44 ± 2.44 bd | 45.42 ± 2.56 b |
| F1 | 67.55 ± 2.50 b | 32.95 ± 2.00 e | 21.71 ± 1.34 c |
| L/F0.1 | 74.93 ± 2.06 ad | 52.91 ± 2.44 cd | 44.95 ± 2.82 b |
| L/F1 | 69.03 ± 2.78 bcd | 39.01 ± 1.90 e | 28.78 ± 2.25 d |

2.4. Effect of Cooking on the DPPH Free Radical Scavenging Activity of Seaweed Polysaccharides in Pork Meat
 ), fresh; (
), fresh; (  ), cooked.
), fresh; ( ), cooked.
), cooked.
), fresh; ( ), cooked.
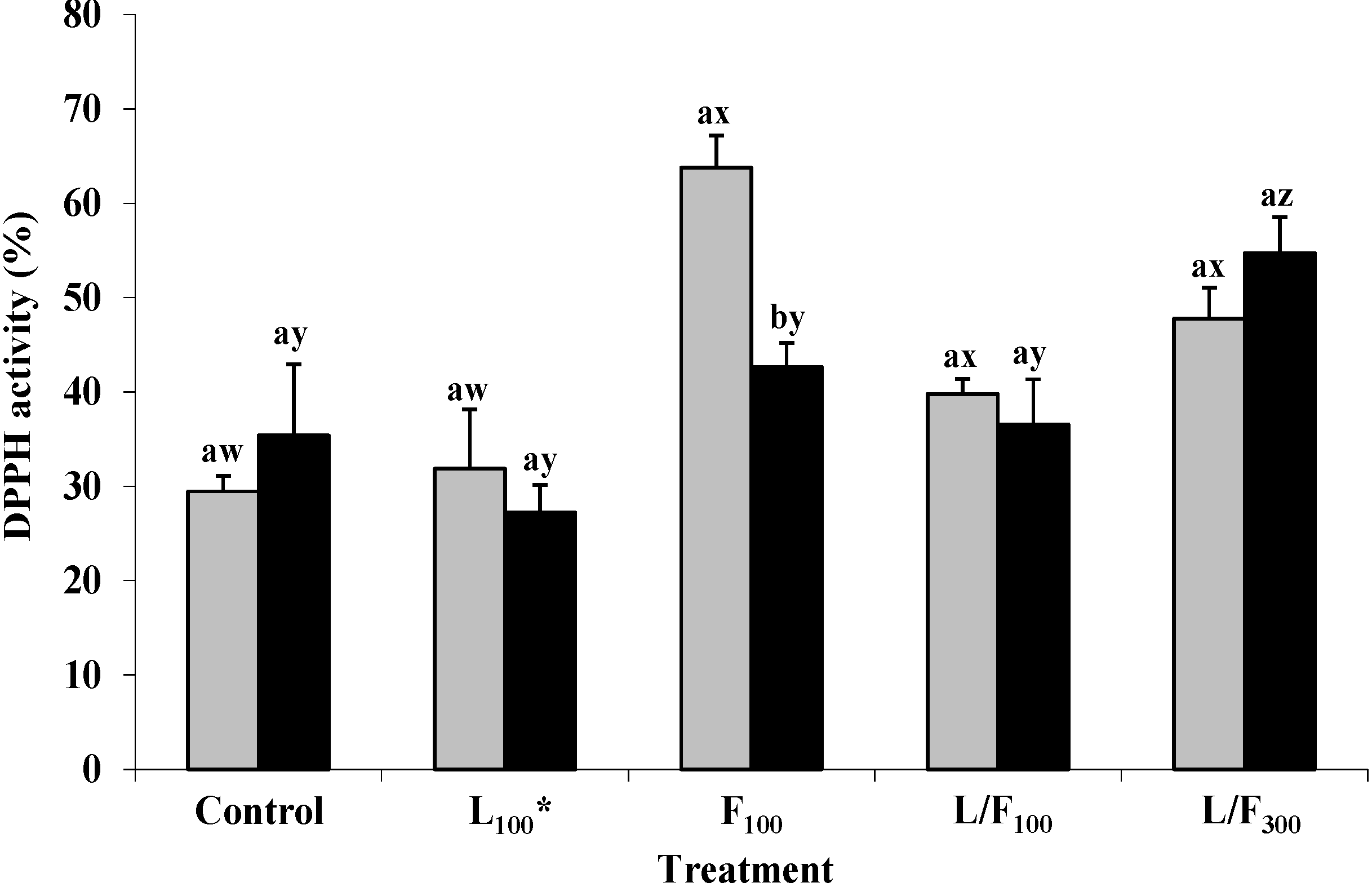
2.5. DPPH Free Radical Scavenging Activity of Seaweed Polysaccharides in Pork Meat Following in Vitro Digestion

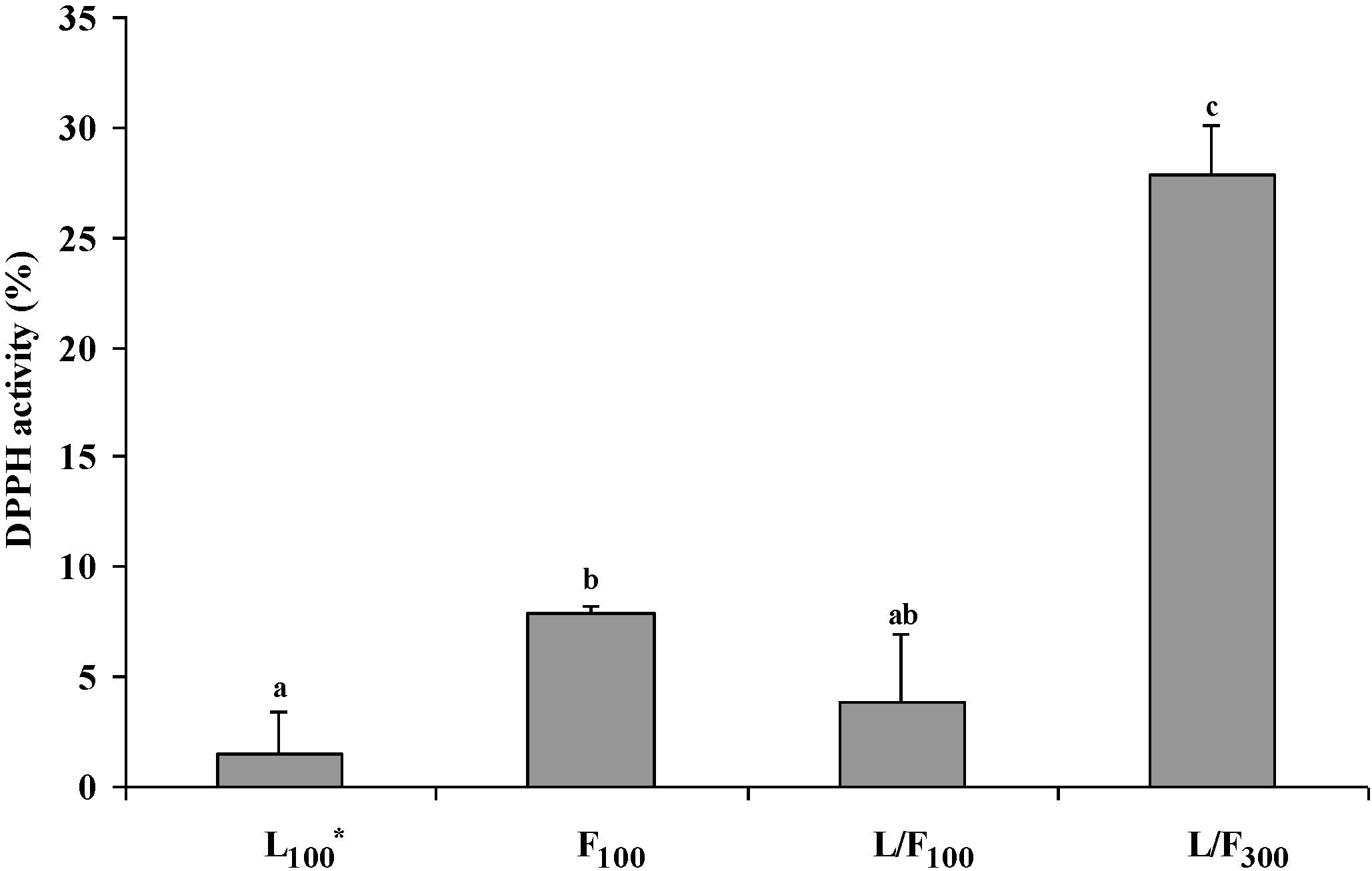
2.6. Bioaccessibility of Seaweed Polysaccharides in Pork Meat after Incubation with Caco-2 Cells
3. Experimental Section
3.1. Reagents
3.2. Measurement of the DPPH Free Radical Scavenging Activities of Seaweed Polysaccharides (L, F and L/F)
3.3. The Effect of Seaweed Polysaccharides on Lipid Oxidation in Pork Muscle Homogenates
Measurement of Lipid Oxidation in Pork Muscle Homogenates
3.4. The Effect of Seaweed Polysaccharides on Oxymyoglobin Oxidation
3.4.1. Preparation of Commercial Oxymyoglobin
3.4.2. Effect of Seaweed Polysaccharides on Oxymyoglobin Oxidation
3.5. Effect of Cooking on DPPH Free Radical Scavenging Activity of Seaweed Polysaccharides in Pork Meat
3.6. Effect of in Vitro Digestion on the DPPH Free Radical Scavenging Activity of Seaweed Polysaccharides in Cooked Pork Meat
3.7. Bioaccessibility and Theoretical Cellular Uptake of the Aqueous Fraction of Digested Minced LTL
3.8. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Ngo, D.H.; Wijesekara, I.; Vo, T.S.; van Ta, Q.; Kim, S.K. Marine food-derived functional ingredients as potential antioxidants in the food industry: An overview. Food Res. Int. 2011, 44, 523–529. [Google Scholar] [CrossRef]
- Devillé, C.; Damas, J.; Forget, P.; Dandrifosse, G.; Peulen, O. Laminarin in the dietary fibre concept. J. Sci. Food Agric. 2004, 84, 1030–1038. [Google Scholar] [CrossRef]
- Choi, J.I.; Kim, H.J.; Lee, J.W. Structural feature and antioxidant activity of low molecular weight laminarin degraded by gamma irradiation. Food Chem. 2011, 129, 520–523. [Google Scholar] [CrossRef]
- Anastyuk, S.D.; Shevchenko, N.M.; Dmitrenok, P.S.; Zvyagintseva, T.N. Structural similarities of fucoidans from brown algae Silvetia babingtonii and Fucus evanescens, determined by tandem MALDI-TOF mass spectrometry. Carbohydr. Res. 2012, 358, 78–81. [Google Scholar] [CrossRef] [PubMed]
- Ustyuzhanina, N.E.; Bilan, M.I.; Ushakova, N.A.; Usov, A.I.; Kiselevskiy, M.V.; Nifantiev, N.E. Fucoidans: Pro- or antiangiogenic agents? Glycobiology 2014, 24, 1265–1274. [Google Scholar] [CrossRef] [PubMed]
- Mak, W.; Hamid, N.; Liu, T.; Lu, J.; White, W.L. Fucoidan from New Zealand Undaria pinnatifida: Monthly variations and determination of antioxidant activities of fucoidan. Carbohydr. Polym. 2013, 95, 606–614. [Google Scholar] [CrossRef] [PubMed]
- Kadam, S.U.; Tiwari, B.K.; O’Donnell, C.P. Extraction, structure and biofunctional activities of laminarin from brown algae. Int. J. Food Sci. Technol. 2014. [Google Scholar] [CrossRef]
- Gómez-Ordóñez, E.; Jiménez-Escrig, A.; Rupérez, P. Bioactivity of sulfated polysaccharides from the edible red seaweed Mastocarpus stellatus. Bioact. Carbohydr. Diet. Fibre 2014, 3, 29–40. [Google Scholar] [CrossRef]
- Koleva, I.I.; van Beek, T.A.; Linssen, J.P.H.; de Groot, A.; Evstatieva, L.N. Screening of plant extracts for antioxidant activity: A comparative study on three testing methods. Phytochem. Anal. 2002, 13, 8–17. [Google Scholar] [CrossRef] [PubMed]
- MacDonald-Wicks, L.K.; Wood, L.G.; Garg, M.L. Methodology for the determination of biological antioxidant capacity in vitro: A review. J. Sci. Food Agric. 2006, 86, 2046–2056. [Google Scholar] [CrossRef]
- Machová, E.; Bystrický, S. Antioxidant capacities of mannans and glucans are related to their susceptibility of free radical degradation. Int. J. Biol. Macromol. 2013, 61, 308–311. [Google Scholar] [CrossRef] [PubMed]
- O’Doherty, J.V.; Dillon, S.; Figat, S.; Callan, J.J.; Sweeney, T. The effects of lactose inclusion and seaweed extract derived from Laminaria spp. on performance, digestibility of diet components and microbial populations in newly weaned pigs. Anim. Feed Sci. Technol. 2010, 157, 173–180. [Google Scholar] [CrossRef]
- Moroney, N.C.; O’Grady, M.N.; Robertson, R.C.; Stanton, C.; O’Doherty, J.V.; Kerry, J.P. Influence of level and duration of feeding polysaccharide (laminarin and fucoidan) extracts from brown seaweed (Laminaria digitata) on quality indices of fresh pork. Meat Sci. 2015, 99, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Moroney, N.C.; O’Grady, M.N.; O’Doherty, J.V.; Kerry, J.P. Addition of seaweed (Laminaria digitata) extracts containing laminarin and fucoidan to porcine diets: Influence on the quality and shelf-life of fresh pork. Meat Sci. 2012, 92, 423–429. [Google Scholar] [CrossRef] [PubMed]
- Moroney, N.C.; O’Grady, M.N.; O’Doherty, J.V.; Kerry, J.P. Effect of a brown seaweed (Laminaria digitata) extract containing laminarin and fucoidan on the quality and shelf-life of fresh and cooked minced pork patties. Meat Sci. 2013, 94, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Elleuch, M.; Bedigian, D.; Roiseux, O.; Besbes, S.; Blecker, C.; Attia, H. Dietary fibre and fibre-rich by-products of food processing: Characterisation, technological functionality and commercial applications: A review. Food Chem. 2011, 124, 411–421. [Google Scholar] [CrossRef]
- Rawson, A.; Patras, A.; Tiwari, B.K.; Noci, F.; Koutchma, T.; Brunton, N. Effect of thermal and non thermal processing technologies on the bioactive content of exotic fruits and their products: Review of recent advances. Food Res. Int. 2011, 44, 1875–1887. [Google Scholar] [CrossRef]
- Gazzani, G.; Papetti, A.; Massolini, G.; Daglia, M. Anti- and prooxidant activity of water soluble components of some common diet vegetables and the effect of thermal treatment. J. Agric. Food Chem. 1998, 46, 4118–4122. [Google Scholar] [CrossRef]
- Gawlik-Dziki, U.; Dziki, D.; Baraniak, B.; Lin, R. The effect of simulated digestion in vitro on bioactivity of wheat bread with Tartary buckwheat flavones addition. LWT Food Sci. Technol. 2009, 42, 137–143. [Google Scholar] [CrossRef]
- Carbonell-Capella, J.M.; Buniowska, M.; Barba, F.J.; Esteve, M.J.; Frígola, A. Analytical methods for determining bioavailability and bioaccessibility of bioactive compounds from fruits and vegetables: A review. Compr. Rev. Food Sci. Food Saf. 2014, 13, 155–171. [Google Scholar] [CrossRef]
- Hur, S.J.; Lim, B.O.; Decker, E.A.; McClements, D.J. In vitro human digestion models for food applications. Food Chem. 2011, 125, 1–12. [Google Scholar] [CrossRef]
- Mirjana, M.; Jelena, A.; Aleksandra, U.; Svetlana, D.; Nevena, G.; Jelena, M.; Ibrahim, M.; Ana, Š.D.; Goran, P.; Melita, V. β-Glucan administration to diabetic rats reestablishes redox balance and stimulates cellular pro-survival mechanisms. J. Funct. Foods 2013, 5, 267–278. [Google Scholar] [CrossRef]
- Tsiapali, E.; Whaley, S.; Kalbfleisch, J.; Ensley, H.E.; Browder, I.W.; Williams, D.L. Glucans exhibit weak antioxidant activity, but stimulate macrophage free radical activity. Free Radic. Biol. Med. 2001, 30, 393–402. [Google Scholar] [CrossRef] [PubMed]
- Giese, E.C.; Gascon, J.; Anzelmo, G.; Barbosa, A.M.; da Cunha, M.A.A.; Dekker, R.F.H. Free-radical scavenging properties and antioxidant activities of botryosphaeran and some other β-d-glucans. Int. J. Biol. Macromol. 2014, 72, 125–130. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef] [PubMed]
- Abu-Ghannam, N.; Cox, S. Seaweed-based functional foods. In Bioactive Compounds from Marine Foods: Plant and Animal Sources; Hernandez-Ledesma, B., Herrero, M., Eds.; Wiley & Sons Ltd.: Chichester, UK, 2014; pp. 313–327. [Google Scholar]
- Yuan, Y.V.; Bone, D.E.; Carrington, M.F. Antioxidant activity of dulse (Palmaria palmata) extract evaluated in vitro. Food Chem. 2005, 91, 485–494. [Google Scholar] [CrossRef]
- Zhou, G.; Ma, W.; Yuan, P. Chemical characterization and antioxidant activities of different sulfate content of λ-carrageenan fractions from edible red seaweed Chrondrus ocellatus. Cell.Mol. Biol. 2014, 60. [Google Scholar] [CrossRef]
- Yan, J.K.; Wang, W.Q.; Ma, H.L.; Wu, J.Y. Sulfation and enhanced antioxidant capacity of an exopolysaccharide produced by the medicinal fungus Cordyceps sinensis. Molecules 2012, 18, 167–177. [Google Scholar] [CrossRef] [PubMed]
- Schenkman, K.A.; Marble, D.R.; Burns, D.H.; Feigl, E.O. Myoglobin oxygen dissociation by multiwavelength spectroscopy. J. Appl. Physiol. 1997, 82, 86–92. [Google Scholar] [PubMed]
- Kim, D.Y.; Shin, W.S. Characterisation of bovine serum albumin–fucoidan conjugates prepared via the Maillard reaction. Food Chem. 2015, 173, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Mulloy, B. The specificity of interactions between proteins and sulfated polysaccharides. An. Acad. Brasil. Ciênc. 2005, 77, 651–664. [Google Scholar] [CrossRef] [PubMed]
- Varenne, A.; Gareil, P.; Colliec-Jouault, S.; Daniel, R. Capillary electrophoresis determination of the binding affinity of bioactive sulfated polysaccharides to proteins: study of the binding properties of fucoidan to antithrombin. Anal. Biochem. 2003, 315, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Imeson, A.P.; Ledward, D.A.; Mitchell, J.R. On the nature of the interaction between some anionic polysaccharides and proteins. J. Sci. Food Agric. 1977, 28, 661–668. [Google Scholar] [CrossRef]
- Satoh, Y.; Shikama, K. Autoxidation of oxymyoglobin. A nucleophilic displacement mechanism. J. Biol. Chem. 1981, 256, 10272–10275. [Google Scholar] [PubMed]
- Prabhasankar, P.; Ganesan, P.; Bhaskar, N.; Hirose, A.; Stephen, N.; Gowda, L.R.; Hosokawa, M.; Miyashita, K. Edible Japanese seaweed, wakame (Undaria pinnatifida) as an ingredient in pasta: Chemical, functional and structural evaluation. Food Chem. 2009, 115, 501–508. [Google Scholar] [CrossRef]
- Kuda, T.; Hishi, T.; Maekawa, S. Antioxidant properties of dried product of “haba-nori”, an edible brown alga, Petalonia binghamiae (J. Agaradh) vinogradova. Food Chem. 2006, 98, 545–550. [Google Scholar] [CrossRef]
- Rajauria, G.; Jaiswal, A.K.; Abu-Ghannam, N.; Gupta, S. Effect of hydrothermal processing on colour, antioxidant and free radical scavenging capacities of edible Irish brown seaweeds. Int. J. Food Sci. Technol. 2010, 45, 2485–2493. [Google Scholar] [CrossRef]
- Yoshiki, M.; Tsuge, K.; Tsuruta, Y.; Yoshimura, T.; Koganemaru, K.; Sumi, T.; Matsui, T.; Matsumoto, K. Production of new antioxidant compound from mycosporine-like amino acid, porphyra-334 by heat treatment. Food Chem. 2009, 113, 1127–1132. [Google Scholar] [CrossRef]
- Bedinghaus, A.; Ockerman, H. Antioxidative Maillard reaction products from reducing sugars and free amino acids in cooked ground pork patties. J. Food Sci. 1995, 60, 992–995. [Google Scholar] [CrossRef]
- Escudero, E.; Sentandreu, M.A.N.; Toldrá, F. Characterization of peptides released by in vitro digestion of pork meat. J. Agric. Food Chem. 2010, 58, 5160–5165. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.M.; Muramoto, K.; Yamauchi, F.; Fujimoto, K.; Nokihara, K. Antioxidative properties of histidine-containing peptides designed from peptide fragments found in the digests of a soybean protein. J. Agric. Food Chem. 1998, 46, 49–53. [Google Scholar] [CrossRef] [PubMed]
- Saiga, A.; Tanabe, S.; Nishimura, T. Antioxidant activity of peptides obtained from porcine myofibrillar proteins by protease treatment. J. Agric. Food Chem. 2003, 51, 3661–3667. [Google Scholar] [CrossRef] [PubMed]
- Sarmadi, B.H.; Ismail, A. Antioxidative peptides from food proteins: A review. Peptides 2010, 31, 1949–1956. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, L.; Murphy, B.; McLoughlin, P.; Duggan, P.; Lawlor, P.G.; Hughes, H.; Gardiner, G.E. Prebiotics from marine macroalgae for human and animal health applications. Mar. Drugs 2010, 8, 2038–2064. [Google Scholar] [CrossRef] [PubMed]
- Salyers, A.; Palmer, J.; Wilkins, T. Laminarinase (beta-glucanase) activity in Bacteroides from the human colon. Appl. Environ. Microbiol. 1977, 33, 1118–1124. [Google Scholar] [PubMed]
- Michel, C.; Macfarlane, G. Digestive fates of soluble polysaccharides from marine macroalgae: involvement of the colonic microflora and physiological consequences for the host. J. Appl. Bacteriol. 1996, 80, 349–369. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.Y.; Je, J.Y.; Kim, S.K. Purification and characterization of antioxidant peptide from hoki (Johnius belengerii) frame protein by gastrointestinal digestion. J. Nutr. Biochem. 2007, 18, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Soler-Rivas, C.; Ramírez-Anguiano, A.C.; Reglero, G.; Santoyo, S. Effect of cooking, in vitro digestion and Caco-2 cells absorption on the radical scavenging activities of edible mushrooms. Int. J. Food Sci. Technol. 2009, 44, 2189–2197. [Google Scholar] [CrossRef]
- Palafox-Carlos, H.; Ayala-Zavala, J.F.; González-Aguilar, G.A. The role of dietary fiber in the bioaccessibility and bioavailability of fruit and vegetable antioxidants. J. Food Sci. 2011, 76, R6–R15. [Google Scholar] [CrossRef] [PubMed]
- Qwele, K.; Hugo, A.; Oyedemi, S.O.; Moyo, B.; Masika, P.J.; Muchenje, V. Chemical composition, fatty acid content and antioxidant potential of meat from goats supplemented with Moringa (Moringa oleifera) leaves, sunflower cake and grass hay. Meat Sci. 2013, 93, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Siu, G.M.; Draper, H.H. A survey of the malonaldehyde content of retail meats and fish. J. Food Sci. 1978, 43, 1147–1149. [Google Scholar] [CrossRef]
- Brown, W.D.; Mebine, L.D. Autooxidation of oxymyoglobin. J. Biol. Chem. 1969, 244, 6696–6701. [Google Scholar] [PubMed]
- Krzywicki, K. The determination of haem pigments in meat. Meat Sci. 1982, 7, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Daly, T.; Ryan, E.; Aisling Aherne, S.; O’Grady, M.N.; Hayes, J.; Allen, P.; Kerry, J.P.; O’Brien, N.M. Bioactivity of ellagic acid-, lutein- or sesamol-enriched meat patties assessed using an in vitro digestion and Caco-2 cell model system. Food Res. Int. 2010, 43, 753–760. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moroney, N.C.; O'Grady, M.N.; Lordan, S.; Stanton, C.; Kerry, J.P. Seaweed Polysaccharides (Laminarin and Fucoidan) as Functional Ingredients in Pork Meat: An Evaluation of Anti-Oxidative Potential, Thermal Stability and Bioaccessibility. Mar. Drugs 2015, 13, 2447-2464. https://doi.org/10.3390/md13042447
Moroney NC, O'Grady MN, Lordan S, Stanton C, Kerry JP. Seaweed Polysaccharides (Laminarin and Fucoidan) as Functional Ingredients in Pork Meat: An Evaluation of Anti-Oxidative Potential, Thermal Stability and Bioaccessibility. Marine Drugs. 2015; 13(4):2447-2464. https://doi.org/10.3390/md13042447
Chicago/Turabian StyleMoroney, Natasha C., Michael N. O'Grady, Sinéad Lordan, Catherine Stanton, and Joseph P. Kerry. 2015. "Seaweed Polysaccharides (Laminarin and Fucoidan) as Functional Ingredients in Pork Meat: An Evaluation of Anti-Oxidative Potential, Thermal Stability and Bioaccessibility" Marine Drugs 13, no. 4: 2447-2464. https://doi.org/10.3390/md13042447
APA StyleMoroney, N. C., O'Grady, M. N., Lordan, S., Stanton, C., & Kerry, J. P. (2015). Seaweed Polysaccharides (Laminarin and Fucoidan) as Functional Ingredients in Pork Meat: An Evaluation of Anti-Oxidative Potential, Thermal Stability and Bioaccessibility. Marine Drugs, 13(4), 2447-2464. https://doi.org/10.3390/md13042447