
λ-Carrageenan Suppresses Tomato Chlorotic Dwarf Viroid (TCDVd) Replication and Symptom Expression in Tomatoes

Abstract
:1. Introduction
2. Results
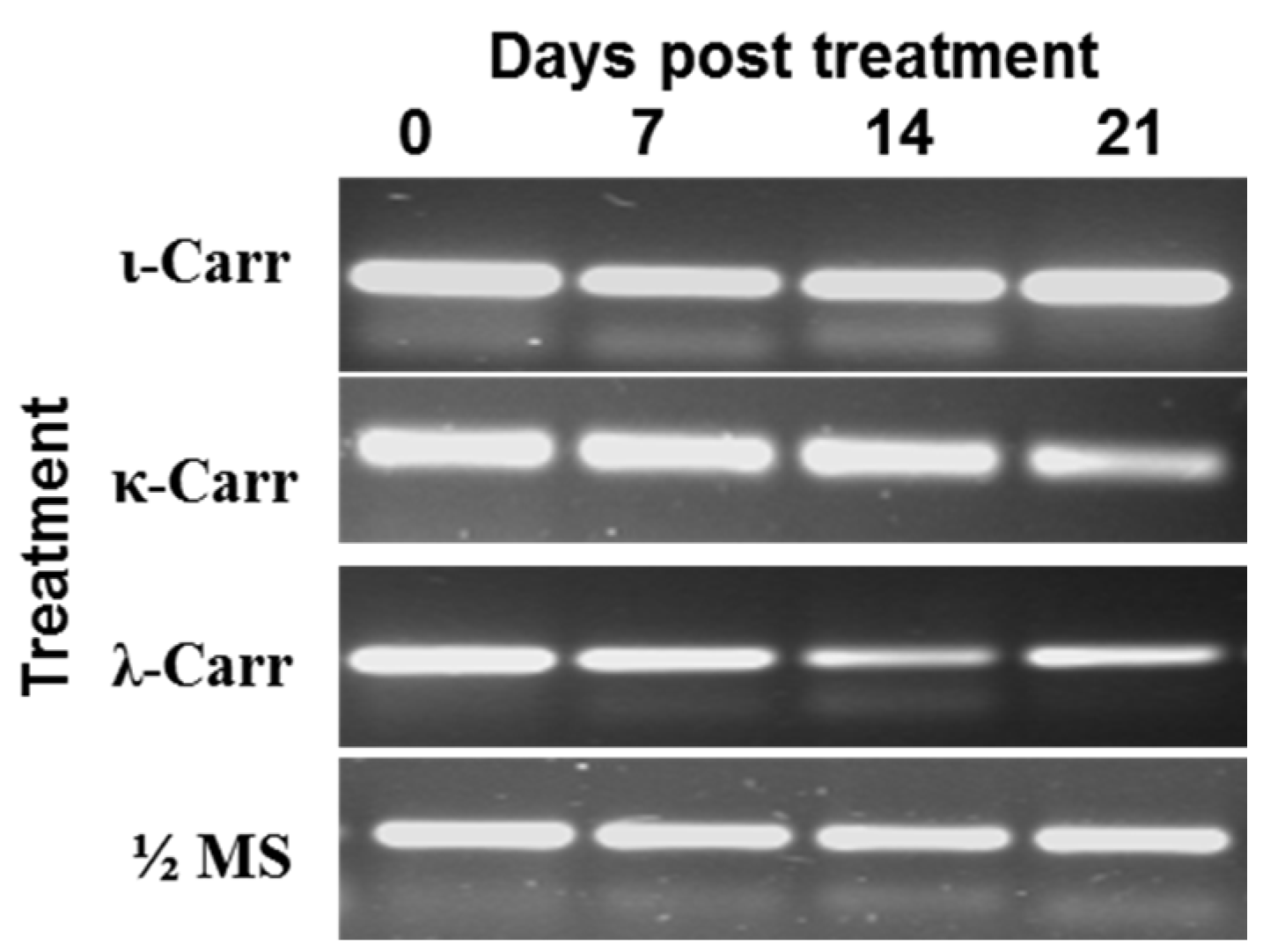
2.1. Effect of Carrageenans on TCDVd Infection in Tomatoes


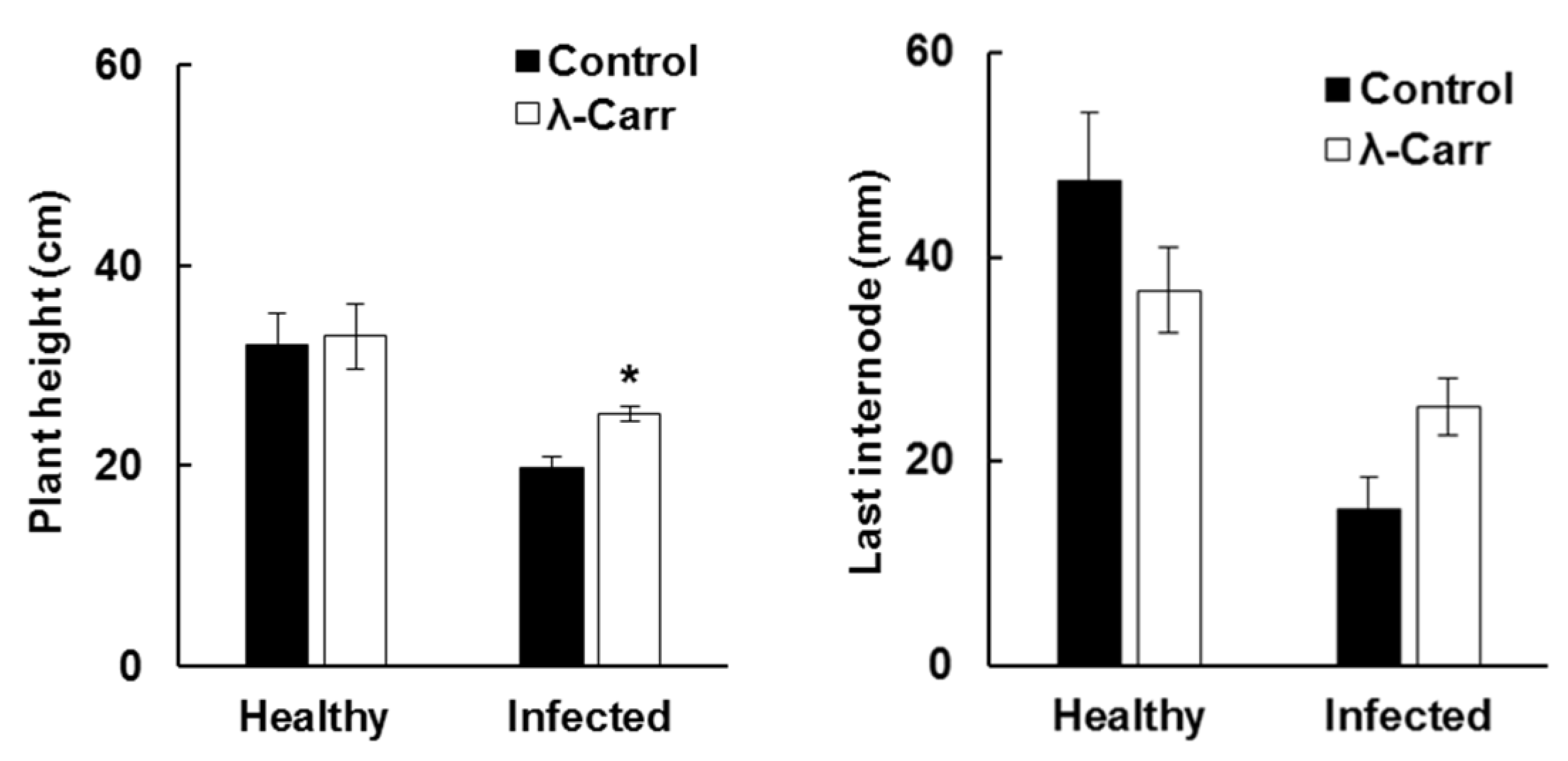
2.2. Effect of Carrageenan Treatments on Plant Height

2.3. Effect of λ-Carrageenan on TCDVd Replication

2.3.1. λ-Carrageenan Induced Differential Expression of Tomato Proteins

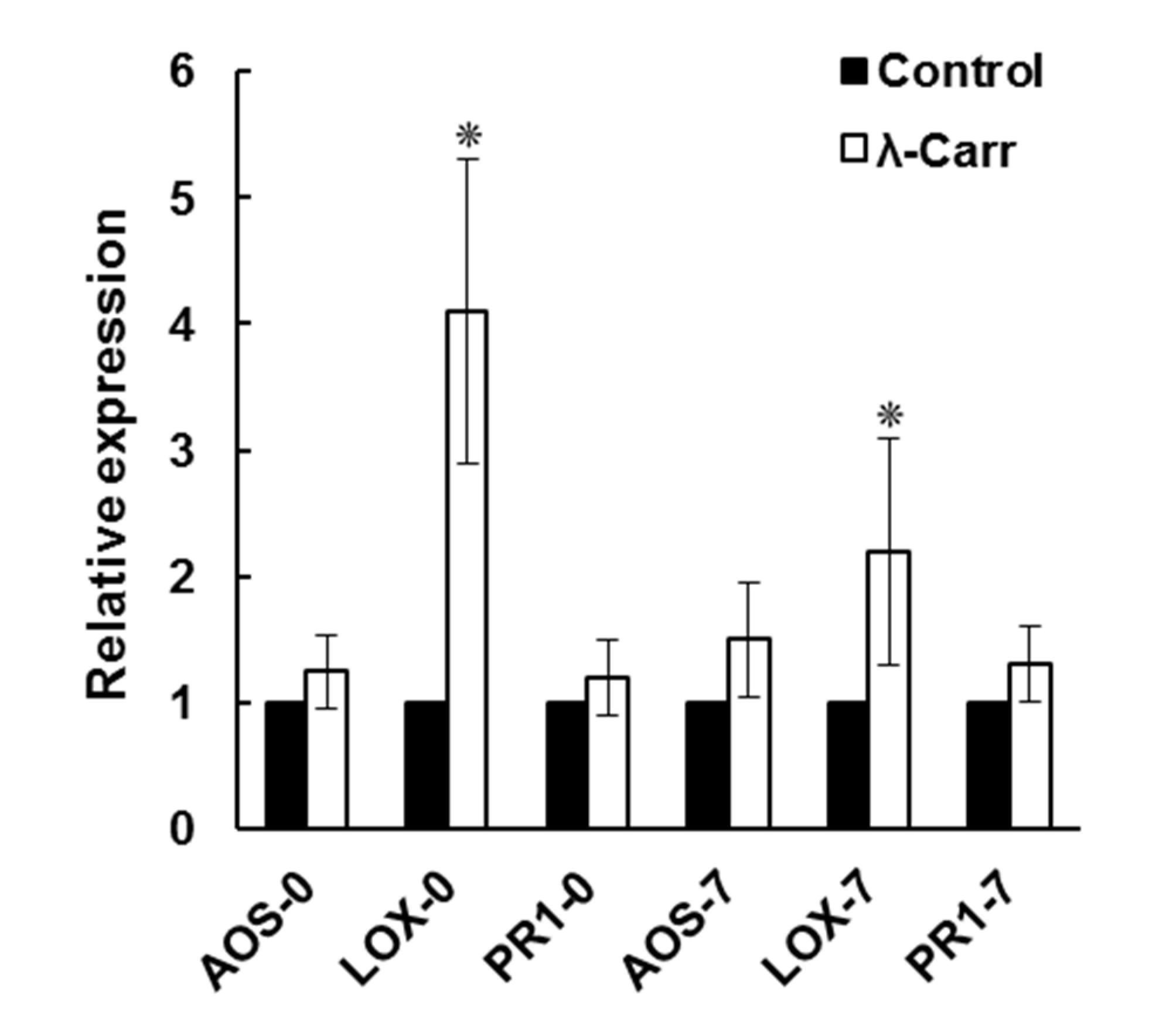
2.3.2. Effect of λ-Carrageenan on the Expression of Defense Response Genes during Viroid Infection

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot ID | Protein Identity | Accession | Molecular Mass (kDa) | Isoelectric Point (pI) | Fold Change | No. of Unique Peptides | Coverage (%) |
|---|---|---|---|---|---|---|---|
| Proteins over expressed in λ-carrageenan treated tomato with viroid infection | |||||||
| 1 | Cytochrome b6-f complex iron-sulfur subunit | Q69GY7-UCRIA_SOLTU | 24.2 | 4.85 | 2.39 | 3 | 12.6 |
| 2 | Photo-system II oxygen-evolving complex protein 2 | Q7M1Y7-Q7M1Y7_ORYSA | 04.0 | 4.90 | 2.37 | 1 | 35.1 |
| 3 | Pathogenesis-related protein 10 | Q4KYL1_ORYSA | 17.5 | 5.59 | 2.45 | 3 | 6.82 |
| 4 | 17.7 kDa class I small heat-shock protein | A1E463_9ASTR | 17.5 | 5.73 | 1.92 | 1 | 14.3 |
| 5 | Superoxide dismutase 1 | P14830-SODC1_SOLLC | 15.2 | 5.66 | 2.70 | 5 | 23.7 |
| 6 | 17.7 kDa classI heat shock protein | O82011-HSP11_SOLPE | 17.7 | 4.65 | 2.43 | 3 | 18.8 |
| 7 | Ribulose bisphosphate (RuBP) carboxylase small chain Fragment | A0A3A2_ARTAN | 19.7 | 5.40 | 2.35 | 1 | 4.62 |
| 8 | Superoxide dismutase | Q6X1D0_SOLLC | 27.9 | 6.45 | 2.90 | 4 | 15.3 |
| 9 | Cytosolic cysteine synthase | Q9FS27_SOLTU | 34.2 | 5.92 | 1.99 | 2 | 7.38 |
| 10 | Proteasome subunit alpha type | Q93X34_TOBAC | 27.1 | 6.21 | 2.49 | 5 | 18.5 |
| 11 | Uracil phosphoribosyltransferase | P93394-UPP_TOBAC | 24.1 | 4.73 | 2.60 | 3 | 16.1 |
| 12 | Cytosolic ascorbate peroxidase 1 | B1Q3F7_SOLLC | 27.3 | 6.00 | 2.85 | 9 | 37.6 |
| 13 | Succinic semialdehyde reductase isoform 2 | B1Q3F7_SOLLC | 38.2 | 4.51 | 1.88 | 1 | 2.75 |
| 14 | Ferredoxin-NADP reductase, leaf-type isozyme, chloroplastic | O04977-FENR1_TOBAC | 40.4 | 6.62 | 1.73 | 3 | 23.8 |
| Proteins under expressed in λ-carrageenan treated tomato with viroid infection | |||||||
| 15 | Carbonic anhydrase | Q5NE20_SOLLC | 34.4 | 6.21 | 1.79 | 3 | 10.6 |
| 16 | Peptidyl-prolyl cis-trans isomerase | A0MTQ0_SOLSG | 17.9 | 5.72 | 1.97 | 1 | 7.02 |
| 17 | Germin like protein | B9A6I8_TOBAC | 21.9 | 4.68 | 2.39 | 1 | 4.27 |

3. Discussion
4. Material and Methods
4.1. Plant and Viroid Culture
4.2. Treatments
4.3. Inoculation of Tomato Plant with TCDVd
4.4. Effect of Carrageenans on TCDVd in Infected Shoots
4.5. Viroid Nucleic Acid Extraction and RT-PCR
4.6. Carrageenan Induced Defense Gene Expression in Tomatoes
4.7. Effect of Carrageenan on Tomato Proteomes
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflict of Interest
References
- Singh, R.P.; Nie, X.; Singh, M. Tomato chlorotic dwarf viroid: An evolutionary link in the origin of pospiviroids. J. Gen. Virol. 1999, 80, 2823–2828. [Google Scholar] [PubMed]
- Diener, T.O. Potato spindle tuber “virus” IV.A replicating, low molecular weight RNA. Virology 1971, 45, 411–428. [Google Scholar]
- Singh, R.P.; Clark, M.C. Infectious low-molecular weight ribonucleic acid from tomato. Biochem. Biophys. Res. Commun. 1971, 44, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Tabler, M.; Tsagris, M. Viroids: Petite RNA pathogens with distinguished talents. Trends Plant Sci. 2004, 9, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Teixeria da Silva, J.A. Ornamental plants: Silent carrier of evolving viroids. In Floriculture, Ornamental and Plant Biotechnology; Teixeria da Silva, J.A., Ed.; Global Science Books: London, UK, 2006; Volume III, pp. 531–539. [Google Scholar]
- Itaya, A.; Matsuda, Y.; Gonzales, R.A.; Nelson, R.S.; Ding, B. Potato spindle tuber viroid strains of different pathogenicity induces and suppresses expression of common and unique genes in infected tomato. Mol. Plant Microbe Interact. 2002, 15, 990–999. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shibuya, M.; Taneda, A.; Kurauchi, T.; Senda, M.; Owens, R.A.; Sano, T. Accumulation of Potato spindle tuber viroid-specific small RNAs is accompanied by specific changes in gene expression in two tomato cultivars. Virology 2011, 413, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Gross, H.J.; Domday, H.; Lossow, C.; Jank, P.; Raba, M.; Alberty, H.; Sänger, H.L. Nucleotide sequence and secondary structure of potato spindle tuber viroid. Nature 1978, 273, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Flores, R.; Serra, P.; Minoia, S.; di Serio, F.; Navarro, B. Viroids: From genotype to phenotype just relying on RNA sequence and structural motifs. Front. Microbiol. 2012, 3, 1–13. [Google Scholar] [PubMed]
- Kovalskaya, N.; Hammond, R.W. Molecular biology of viroid-host interactions and disease control strategies. Plant Sci. 2014, 228, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Crowley, C.F. Evaluation of polyacrylamide gel electrophoresis, bioassay, and dot-blot methods for the survey of potato spindle tuber viroid. Can. Plant Dis. Surv. 1985, 65, 61–63. [Google Scholar]
- Singh, R.P. Piperonyl butoxide as a protectant against Potato spindle tuber viroid infection. Phytopathology 1977, 67, 933–935. [Google Scholar] [CrossRef]
- Bellés, J.M.; Hansen, A.J.; Granell, A.; Conejero, V. Antiviroid effect of ribavirin on citrus exocortis viroid infection in Gynura aurantiaca DC. Physiol. Mol. Plant Pathol. 1986, 28, 61–65. [Google Scholar] [CrossRef]
- Niblett, C.L.; Dickson, E.; Fernow, K.H.; Horst, R.K.; Zaitlin, M. Cross protection among four viroids. Virology 1978, 91, 198–203. [Google Scholar] [CrossRef] [PubMed]
- Postman, J.; Hadidi, A. Eliminati on of apple scar skin viroid from pears by in vitro thermotherapy and apical meristem culture. Acta Hortic. 1995, 386, 536–543. [Google Scholar]
- Matoušek, J.; Trněná, L.; Svoboda, P.; Oriniaková, P.; Lichtenstein, C.P. The gradual reduction of viroid levels in hop mericlones following heat therapy: A possible role for a nuclease degrading dsRNA. Biol. Chem. Hoppe Seyler 1995, 376, 715–722. [Google Scholar] [CrossRef] [PubMed]
- Kasai, A.; Sano, T.; Harada, T. Scion on a stock producing siRNAs of potato spindle tuber viroid (PSTVd) attenuates accumulation of the viroid. PLoS ONE 2013, 2, e57736. [Google Scholar] [CrossRef]
- Harris, P.S.; Miller-Jones, D.M.; Howell, P.J. Control of potato spindle tuber viroid: The special problems of a disease in plant breeders’ material. In Plant Health: The Scientific Basis for Administrative Control of Plant Parasites; Ebbels, D.L., King, J.E., Eds.; Blackwell: Oxford, UK, 1979; pp. 232–237. [Google Scholar]
- Pfannenstiel, M.A.; Slack, S.A. Response of potato cultivars to infection by potato spindle tuber viroid. Phytopathology 1980, 70, 922–926. [Google Scholar] [CrossRef]
- Matsushita, Y.; Aoki, K.; Sumitomo, K. Selection and inheritance of resistance to Chrysanthemum stunt viroid. Crop Prot. 2012, 35, 1–4. [Google Scholar] [CrossRef]
- Omori, H.; Hosokawa, M.; Shiba, H.; Shitsukawa, N.; Murai, K.; Yazawa, S. Screening of chrysanthemum plants with strong resistance to Chrysanthemum stunt viroid. J. Jpn. Soc. Hortic. Sci. 2009, 78, 350–355. [Google Scholar] [CrossRef]
- Schwind, N.; Zwiebel, M.; Itaya, A.; Ding, B.; Wang, M.-B.; Krczal, G.; Wassenegger, M. RNAi mediated resistance to Potato spindle tuber viroid in transgenic tomato expressing a viroid hairpin RNA construct. Mol. Plant Pathol. 2009, 10, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.J.; Dicke, M. Plant interactions with microbes and insects: From molecular mechanisms to ecology. Trends Plant Sci. 2007, 12, 564–569. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar] [CrossRef] [PubMed]
- Scheel, D. Resistance response physiology and signal transduction. Curr. Opin. Plant Biol. 1998, 1, 305–310. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.J.; van Wees, S.C.M.; van Pelt, J.A.; Knoester, M.; Laan, R.; Gerrits, H.; Weisbeek, P.J.; van Loon, L.C. A novel signaling pathway controlling induced systemic resistance in Arabidopsis. Plant Cell 1998, 10, 1571–1580. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Harrison, M.J.; Lamb, C.J. Early events in the activation of plant defense responses. Annu. Rev. Plant Phytopathol. 1994, 32, 479–501. [Google Scholar] [CrossRef]
- Mercier, L.; Lafitte, C.; Borderies, G.; Briand, X.; Esquerré-Tugayé, M.T.; Fournier, J. The algal polysaccharide carrageenans can act as an elicitor of plant defence. New Phytol. 2001, 149, 43–51. [Google Scholar] [CrossRef]
- Bektas, Y.; Eulgem, T. Synthetic Plant Defense Elicitors. Front. Plant Sci. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Klarzynski, O.; Plesse, B.; Joubert, J.M.; Yvin, J.C.; Kopp, M.; Kloareg, B.; Fritig, B. Linear beta-1, 3 glucans are elicitors of defense responses in tobacco. Plant Physiol. 2000, 124, 1027–1038. [Google Scholar] [CrossRef] [PubMed]
- Sangha, J.S.; Ravichandran, S.; Prithiviraj, K.; Critchley, A.T.; Prithiviraj, B. Sulfated macroalgal polysaccharides λ-carrageenan and ι-carrageenan differentially alter Arabidopsis thaliana resistance to Sclerotinia sclerotiorum. Physiol. Mol. Plant Pathol. 2010, 75, 38–45. [Google Scholar] [CrossRef]
- Gordon-Mills, E.M.; McCandless, E.L. Carrageenans in the cell walls of Chondrus crispus Stack. (Rhodophyceae, Gigartinales). I. Localization with fluorescent antibody. Phycologia 1975, 14, 275–281. [Google Scholar]
- Buck, C.B.; Thompson, C.D.; Roberts, J.N.; Müller, M.; Lowy, D.R.; Schiller, J.T. Carrageenan is a potent inhibitor of papillomavirus infection. PLoS Pathog. 2006, 2, e69. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, A.; Tai, A.; Kanzaki, H.; Kawazu, K. Elicitor-active oligosaccharides from algal laminaran stimulate the production of antifungal compounds in alfalfa. Z. Naturforschung 1993, 48, 575–579. [Google Scholar]
- Bouarab, K.; Potin, P.; Correa, J.; Kloareg, B. Sulfated oligosaccharides mediate the interaction between a marine red alga and its green algal pathogenic endophyte. Plant Cell 1999, 11, 1635–1650. [Google Scholar] [CrossRef] [PubMed]
- Khan, W.; Rayirath, U.P.; Subramanian, S.; Jithesh, M.N.; Rayorath, P.; Hodges, D.M.; Critchley, A.T.; Craigie, J.S.; Norrie, J.; Prithiviraj, P. Seaweed extracts as biostimulants of plant growth and development. J. Plant Growth Regul. 2009, 28, 386–399. [Google Scholar] [CrossRef]
- Singh, R.P.; Dilworth, A.D.; Singh, M.; Babcock, K.M. An alkaline solution simplifies nucleic acid preparation for RT-PCR and infectivity assays of viroids from crude sap and spotted membrane. J. Virol. Methods 2006, 132, 204–211. [Google Scholar] [CrossRef] [PubMed]
- Bostock, R.M.; Karban, R.; Thaler, J.S.; Weyman, P.D.; Gilchrist, D. Signal interactions in induced resistance to pathogens and insect herbivores. Eur. J. Plant Pathol. 2008, 107, 103–111. [Google Scholar] [CrossRef]
- Zimmerli, L.; Jakab, G.; Métraux, J.P.; Mauch-Mani, B. Potentiation of pathogen-specific defense mechanisms in Arabidopsis by beta aminobutyric acid. Proc. Natl. Acad. Sci. USA 2000, 97, 12920–12925. [Google Scholar] [CrossRef] [PubMed]
- Nagorskaia, V.P.; Reunov, A.V.; Lapshina, L.A.; Ermak, I.M.; Barabanova, A.O. Influence of kappa/beta-carrageenan from red alga Tichocarpus crinitus on development of local infection induced by tobacco mosaic virus in Xanthi-nc tobacco leaves. Izv. Akad. Nauk Ser. Biol. 2007, 3, 360–364. [Google Scholar] [PubMed]
- Sangha, J.S.; Khan, W.; Ji, X.; Zhang, J.; Mills, A.A.; Critchley, A.T.; Prithiviraj, B. Carrageenans, sulphated polysaccharides of red seaweeds, differentially affect Arabidopsis thaliana resistance to Trichoplusia ni (Cabbage Looper). PLoS ONE 2011, 6, e26834. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.P.; Dilworth, A.D.; Ao, X.; Singh, M.; Misra, S. Molecular and biological characterization of a severe isolate of Tomato chlorotic dwarf viroid containing a novel terminal right (TR) domain sequence. Eur. J. Plant Pathol. 2010, 127, 63–72. [Google Scholar] [CrossRef]
- Rampitsch, C.; Natalia, V.B. Proteomics and plant disease: Advances in combating a major threat to the global food supply. Proteomics 2012, 12, 673–690. [Google Scholar] [CrossRef] [PubMed]
- Bostan, H.; Nie, X.; Singh, R.P. An RT-PCR primer pair for the detection of Pospiviroid and its application in surveying ornamental plants for viroids. J. Virol. Methods 2004, 116, 189–193. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, A.; Wilm, M.; Vorm, O.; Mann, M. Mass spectrometric sequencing of proteins from silver-stained polyacrylamide gels. Anal. Chem. 1996, 68, 850–858. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sangha, J.S.; Kandasamy, S.; Khan, W.; Bahia, N.S.; Singh, R.P.; Critchley, A.T.; Prithiviraj, B. λ-Carrageenan Suppresses Tomato Chlorotic Dwarf Viroid (TCDVd) Replication and Symptom Expression in Tomatoes. Mar. Drugs 2015, 13, 2875-2889. https://doi.org/10.3390/md13052875
Sangha JS, Kandasamy S, Khan W, Bahia NS, Singh RP, Critchley AT, Prithiviraj B. λ-Carrageenan Suppresses Tomato Chlorotic Dwarf Viroid (TCDVd) Replication and Symptom Expression in Tomatoes. Marine Drugs. 2015; 13(5):2875-2889. https://doi.org/10.3390/md13052875
Chicago/Turabian StyleSangha, Jatinder S., Saveetha Kandasamy, Wajahatullah Khan, Navratan Singh Bahia, Rudra P. Singh, Alan T. Critchley, and Balakrishnan Prithiviraj. 2015. "λ-Carrageenan Suppresses Tomato Chlorotic Dwarf Viroid (TCDVd) Replication and Symptom Expression in Tomatoes" Marine Drugs 13, no. 5: 2875-2889. https://doi.org/10.3390/md13052875
APA StyleSangha, J. S., Kandasamy, S., Khan, W., Bahia, N. S., Singh, R. P., Critchley, A. T., & Prithiviraj, B. (2015). λ-Carrageenan Suppresses Tomato Chlorotic Dwarf Viroid (TCDVd) Replication and Symptom Expression in Tomatoes. Marine Drugs, 13(5), 2875-2889. https://doi.org/10.3390/md13052875