
γ-Lindane Increases Microcystin Synthesis in Microcystis aeruginosa PCC7806

Abstract
:1. Introduction
2. Results and Discussion
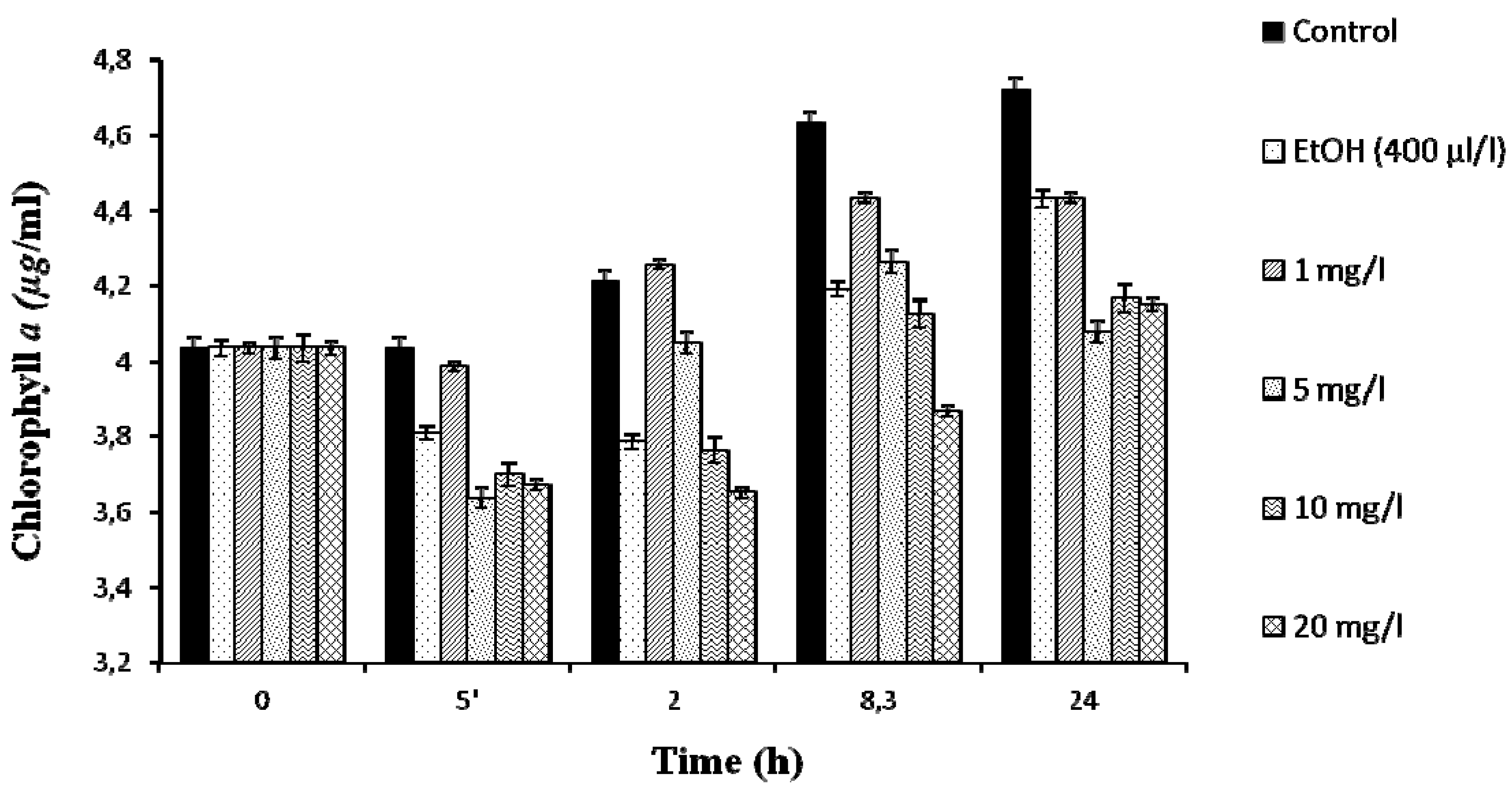
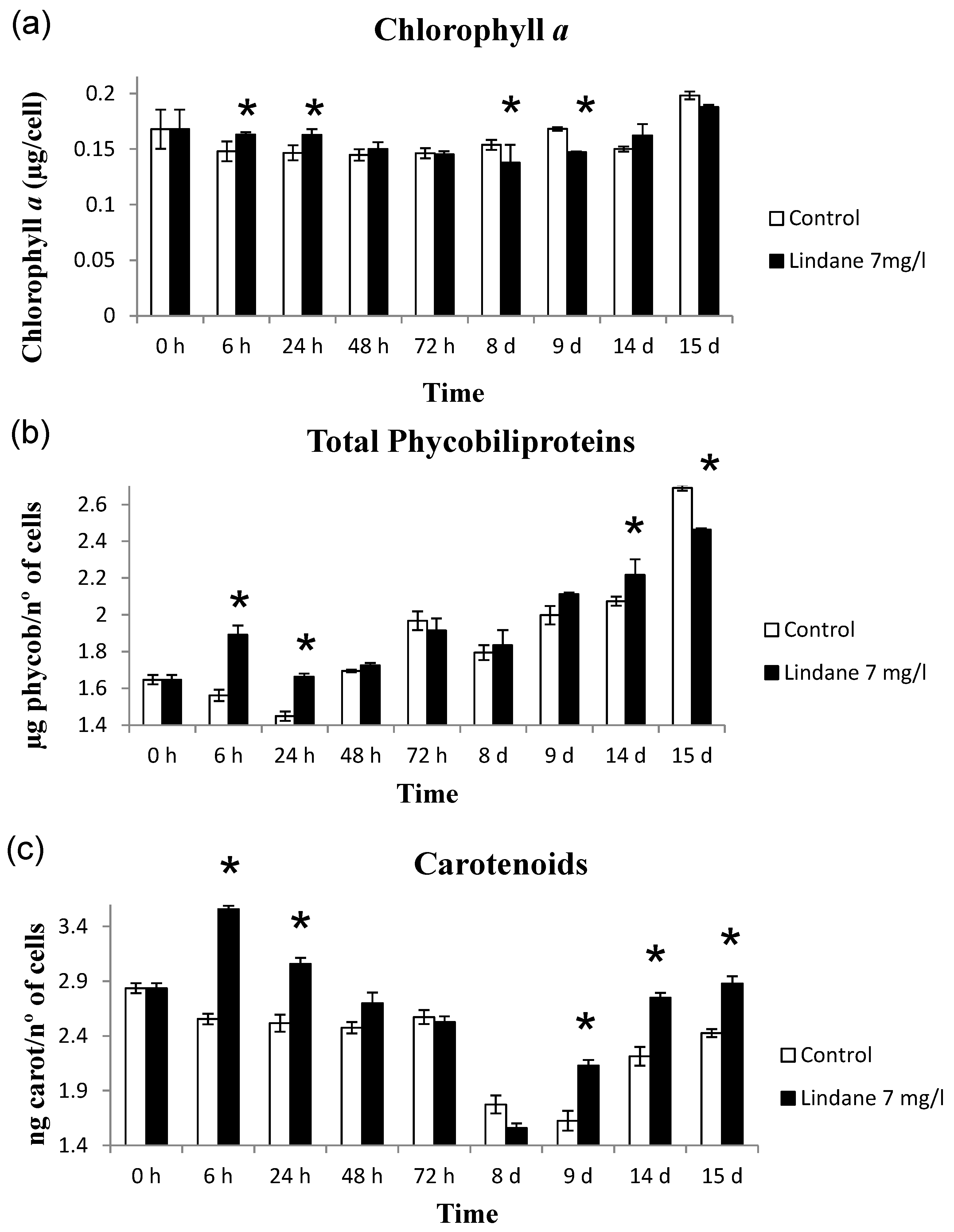
2.1. Effects of Lindane on M. Aeruginosa PCC7806 Growth


2.2. Effects of Lindane on Gene Expression Levels




2.3. M. aeruginosa Degrades Lindane

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Time 24 h mg/L lindane | Time 15 days mg/L lindane |
|---|---|---|
| M. aeruginosa culture without lindane | <0.005 | <0.005 |
| Culture media 7 mg/L lindane | 6.27 ± 0.34 | 2.08 ± 0.26 |
| M. aeruginosa culture 7 mg/L lindane | 6.19 ± 0.29 | 0.36 ± 0.05 |

2.4. Microcystin-LR Content Correlates with mcyD Transcriptional Changes Caused by Lindane
2.5. Discussion

3. Experimental Section
3.1. Growth Conditions
3.2. Analytical Methods and Microcystin Quantification
3.3. Cell Counting
3.4. Sampling and RNA Isolation
3.5. Reverse Transcription (cDNA Synthesis)
| Primers | Sequence 5′ → 3′ | Length | Tm (°C) |
|---|---|---|---|
| R16S dir | CAAGTCGAACGGGAATCTTC | 20 | 47 |
| R16S rev | CTCAAGTACCGTCAGAACTTC | 21 | |
| mcy D dir | GAGCATTAAGGGCTAAATCG | 20 | 45 |
| mcy D rev | CTTGGTTGCTTCATCAACTC | 20 | |
| mcy J dir | GCCGAAGAAACAACTTATAACG | 22 | 48 |
| mcy J rev | CTATAGCCAAGCTTCCACCGGG | 22 | |
| RT-mcy H up | GGTATGAATGCAGCAG | 16 | 45 |
| RT-mcy H dw | CGCCTGGTTCGATAGG | 16 | |
| ntcA up | GGAATTCCATATGGACTTATCATTAATACAAGATAAAC | 38 | 54 |
| ntcA dw | CCCAAGCTTTTAAGTAAATTGTTGACTGAGAG | 32 | |
| furA dir | GTCGATCGCCCATGGCTGCCTAC | 23 | 65 |
| furA rev | CAGTTGGGAATTCCCGCTAGATG | 24 | |
| αfurA dir | CGACGATTTACCGCAGTG | 18 | 53 |
| αfurA rev | CACACTGTTTGAGACTGTG | 19 | |
| nirA dir | TGCCCATTCTACTCAACCCTA | 21 | 58 |
| nirA rev | GTGTCGCTAATCCCCATAGTTG | 22 | |
| fur B myc1 dir | CAATCTATGGGWYTAGCTACYGT | 23 | 60 |
| fur B myc1 rev1 | CCGCAIARWCCAAAAAATTCIARIGTATG | 29 | |
| fur C myc dir | CATYTITCTGCTMGIGAAATTTATGATCC | 29 | 60 |
| fur C myc rev | CATGIGAATGIGAATCIGAAATAKTWCC | 28 |
3.6. RT-PCR Analysis of Gene Expression
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Carmichael, W.W.; Azevedo, S.M.; An, J.S.; Molica, R.J.; Jochimsen, E.M.; Lau, S.; Rinehart, K.L.; Shaw, G.R.; Eaglesham, G.K. Human fatalities from cyanobacteria: Chemical and biological evidence for cyanotoxins. Environ. Health Perspect. 2001, 109, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Codd, G.A.; Morrison, L.F.; Metcalf, J.S. Cyanobacterial toxins: Risk management for health protection. Toxicol. Appl. Pharmacol. 2005, 203, 264–272. [Google Scholar] [CrossRef] [PubMed]
- Black, K.; Yilmaz, M.; Phlips, E.J. Growth and toxin production by Microcystis aerugionosa PCC 7806 (Kutzing) Lemmerman at elevated salt concentrations. J. Environ. Prot. 2011, 2, 669–674. [Google Scholar] [CrossRef]
- Tillett, D.; Dittmann, E.; Erhard, M.; von Dohren, H.; Borner, T.; Neilan, B.A. Structural organization of microcystin biosynthesis in Microcystis aeruginosa PCC7806: An integrated peptide-polyketide synthetase system. Chem. Biol. 2000, 7, 753–764. [Google Scholar] [CrossRef]
- Neilan, B.A.; Pearson, L.A.; Muenchhoff, J.; Moffitt, M.C.; Dittmann, E. Environmental conditions that influence toxin biosynthesis in cyanobacteria. Environ. Microbiol. 2012, 15, 1239–1253. [Google Scholar] [CrossRef] [PubMed]
- Kuritz, T.; Wolk, C.P. Use of filamentous cyanobacteria for biodegradation of organic pollutants. Appl. Environ. Microbiol. 1995, 61, 234–238. [Google Scholar] [PubMed]
- Kuritz, T. Cyanobacteria as agents for the control of pollution by pesticides and chlorinated organic compounds. J. Appl. Microbiol. 1998, 85 (Suppl. 1), 186S–192S. [Google Scholar] [CrossRef] [PubMed]
- Vijgen, J.; Abhilash, P.C.; Li, Y.F.; Lal, R.; Forter, M.; Torres, J.; Singh, N.; Yunus, M.; Tian, C.; Schaffer, A.; Weber, R. Hexachlorocyclohexane (HCH) as new Stockholm Convention POPs—A global perspective on the management of Lindane and its waste isomers. Environ. Sci. Pollut. Res. Intl. 2011, 18, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Phillips, T.M.; Seech, A.G.; Lee, H.; Trevors, J.T. Biodegradation of hexachlorocyclohexane (HCH) by microorganisms. Biodegradation 2005, 16, 363–392. [Google Scholar] [CrossRef] [PubMed]
- Kuritz, T.; Bocanera, L.V.; Rivera, N.S. Dechlorination of lindane by the cyanobacterium Anabaena sp. strain PCC7120 depends on the function of the nir operon. J. Bacteriol. 1997, 179, 3368–3370. [Google Scholar] [PubMed]
- Gonzalez, R.; Garcia-Balboa, C.; Rouco, M.; Lopez-Rodas, V.; Costas, E. Adaptation of microalgae to lindane: A new approach for bioremediation. Aquat. Toxicol. 2012, 109, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Qian, H.; Pan, X.; Chen, J.; Zhou, D.; Chen, Z.; Zhang, L.; Fu, Z. Analyses of gene expression and physiological changes in Microcystis aeruginosa reveal the phytotoxicities of three environmental pollutants. Ecotoxicology 2012, 21, 847–859. [Google Scholar] [CrossRef] [PubMed]
- Suresh Babu, G.; Hans, R.K.; Singh, J.; Viswanathan, P.N.; Joshi, P.C. Effect of lindane on the growth and metabolic activities of cyanobacteria. Ecotoxicol. Environ. Saf. 2001, 48, 219–221. [Google Scholar] [CrossRef] [PubMed]
- Bueno, M.; Fillat, M.F.; Strasser, R.J.; Maldonado-Rodriguez, R.; Marina, N.; Smienk, H.; Gomez-Moreno, C.; Barja, F. Effects of lindane on the photosynthetic apparatus of the cyanobacterium Anabaena: Fluorescence induction studies and immunolocalization of ferredoxin-NADP+ reductase. Environ. Sci. Pollut. Res. Intl. 2004, 11, 98–106. [Google Scholar] [CrossRef]
- Kuniyoshi, T.M.; Gonzalez, A.; Lopez-Gomollon, S.; Valladares, A.; Bes, M.T.; Fillat, M.F.; Peleato, M.L. 2-oxoglutarate enhances NtcA binding activity to promoter regions of the microcystin synthesis gene cluster. FEBS Lett. 2011, 585, 3921–3926. [Google Scholar] [CrossRef] [PubMed]
- Martin-Luna, B.; Sevilla, E.; Hernandez, J.A.; Bes, M.T.; Fillat, M.F.; Peleato, M.L. Fur from Microcystis aeruginosa binds in vitro promoter regions of the microcystin biosynthesis gene cluster. Phytochemistry 2006, 67, 876–881. [Google Scholar] [CrossRef] [PubMed]
- Sevilla, E.; Martin-Luna, B.; Gonzalez, A.; Gonzalo-Asensio, J.A.; Peleato, M.L.; Fillat, M.F. Identification of three novel antisense RNAs in the fur locus from unicellular cyanobacteria. Microbiology 2011, 157, 3398–3404. [Google Scholar] [CrossRef] [PubMed]
- Wiedner, C.; Visser, P.M.; Fastner, J.; Metcalf, J.S.; Codd, G.A.; Mur, L.R. Effects of light on the microcystin content of Microcystis strain PCC 7806. Appl. Environ. Microbiol. 2003, 69, 1475–1481. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, J.; Arjol, M.A.; Cacho, C. POP-contaminated sites from HCH production in Sabinanigo, Spain. Environ. Sci. Pollut. Res. 2013, 20, 1937–1950. [Google Scholar] [CrossRef] [PubMed]
- Sevilla, E.; Martin-Luna, B.; Bes, M.T.; Fillat, M.F.; Peleato, M.L. An active photosynthetic electron transfer chain required for mcyD transcription and microcystin synthesis in Microcystis aeruginosa PCC7806. Ecotoxicology 2012, 21, 811–819. [Google Scholar] [CrossRef] [PubMed]
- Zilliges, Y.; Kehr, J.C.; Meissner, S.; Ishida, K.; Mikkat, S.; Hagemann, M.; Kaplan, A.; Borner, T.; Dittmann, E. The cyanobacterial hepatotoxin microcystin binds to proteins and increases the fitness of microcystis under oxidative stress conditions. PLoS ONE 2011, 6, e17615. [Google Scholar] [CrossRef] [PubMed]
- Pearson, L.A.; Hisbergues, M.; Borner, T.; Dittmann, E.; Neilan, B.A. Inactivation of an ABC transporter gene, mcyH, results in loss of microcystin production in the cyanobacterium Microcystis aeruginosa PCC 7806. Appl. Environ. Microbiol. 2004, 70, 6370–6378. [Google Scholar] [CrossRef] [PubMed]
- Rippka, R.; Deruelles, J.B.; Waterbury, M.; Herdman, M.; Stanier, R.Y. Genetics asignments, strain stories and properties of pure cultures of cyanobacteria. J. Gen. Microbiol. 1979, 11, 1–61. [Google Scholar] [CrossRef]
- Mackinney, G. Absorption of light by chlorophyll solutions. J. Biol. Chem. 1941, 140, 109–112. [Google Scholar]
- Sevilla, E.; Martin-Luna, B.; Vela, L.; Bes, M.T.; Fillat, M.F.; Peleato, M.L. Iron availability affects mcyD expression and microcystin-LR synthesis in Microcystis aeruginosa PCC7806. Environ. Microbiol. 2008, 10, 2476–2483. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.F.; Davies, B.H. Triterpenoid carotenoids and related lipids. Triterpenoid carotenoid aldehydes from Streptococcus faecium UNH 564P. Biochem. J. 1976, 153, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Glazer, A.N.; Apell, G.S.; Hixson, C.S.; Bryant, D.A.; Rimon, S.; Brown, D.M. Biliproteins of cyanobacteria and Rhodophyta: Homologous family of photosynthetic accessory pigments. Proc. Natl. Acad. Sci. USA 1976, 73, 428–431. [Google Scholar] [CrossRef] [PubMed]
- Thurman, R.G.; Ley, H.G.; Scholz, R. Hepatic microsomal ethanol oxidation. Hydrogen peroxide formation and the role of catalase. Eur. J. Biochem. 1972, 25, 420–430. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.; Bes, M.T.; Barja, F.; Peleato, M.L.; Fillat, M.F. Overexpression of FurA in Anabaena sp. PCC 7120 reveals new targets for this regulator involved in photosynthesis, iron uptake and cellular morphology. Plant Cell Physiol. 2010, 51, 1900–1914. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ceballos-Laita, L.; Calvo-Begueria, L.; Lahoz, J.; Bes, M.-T.; Fillat, M.F.; Peleato, M.-L. γ-Lindane Increases Microcystin Synthesis in Microcystis aeruginosa PCC7806. Mar. Drugs 2015, 13, 5666-5680. https://doi.org/10.3390/md13095666
Ceballos-Laita L, Calvo-Begueria L, Lahoz J, Bes M-T, Fillat MF, Peleato M-L. γ-Lindane Increases Microcystin Synthesis in Microcystis aeruginosa PCC7806. Marine Drugs. 2015; 13(9):5666-5680. https://doi.org/10.3390/md13095666
Chicago/Turabian StyleCeballos-Laita, Laura, Laura Calvo-Begueria, Jessica Lahoz, María-Teresa Bes, María F. Fillat, and María-Luisa Peleato. 2015. "γ-Lindane Increases Microcystin Synthesis in Microcystis aeruginosa PCC7806" Marine Drugs 13, no. 9: 5666-5680. https://doi.org/10.3390/md13095666
APA StyleCeballos-Laita, L., Calvo-Begueria, L., Lahoz, J., Bes, M. -T., Fillat, M. F., & Peleato, M. -L. (2015). γ-Lindane Increases Microcystin Synthesis in Microcystis aeruginosa PCC7806. Marine Drugs, 13(9), 5666-5680. https://doi.org/10.3390/md13095666