
New Typical Vector of Neurotoxin β-N-Methylamino-l-Alanine (BMAA) in the Marine Benthic Ecosystem

Abstract
:1. Introduction
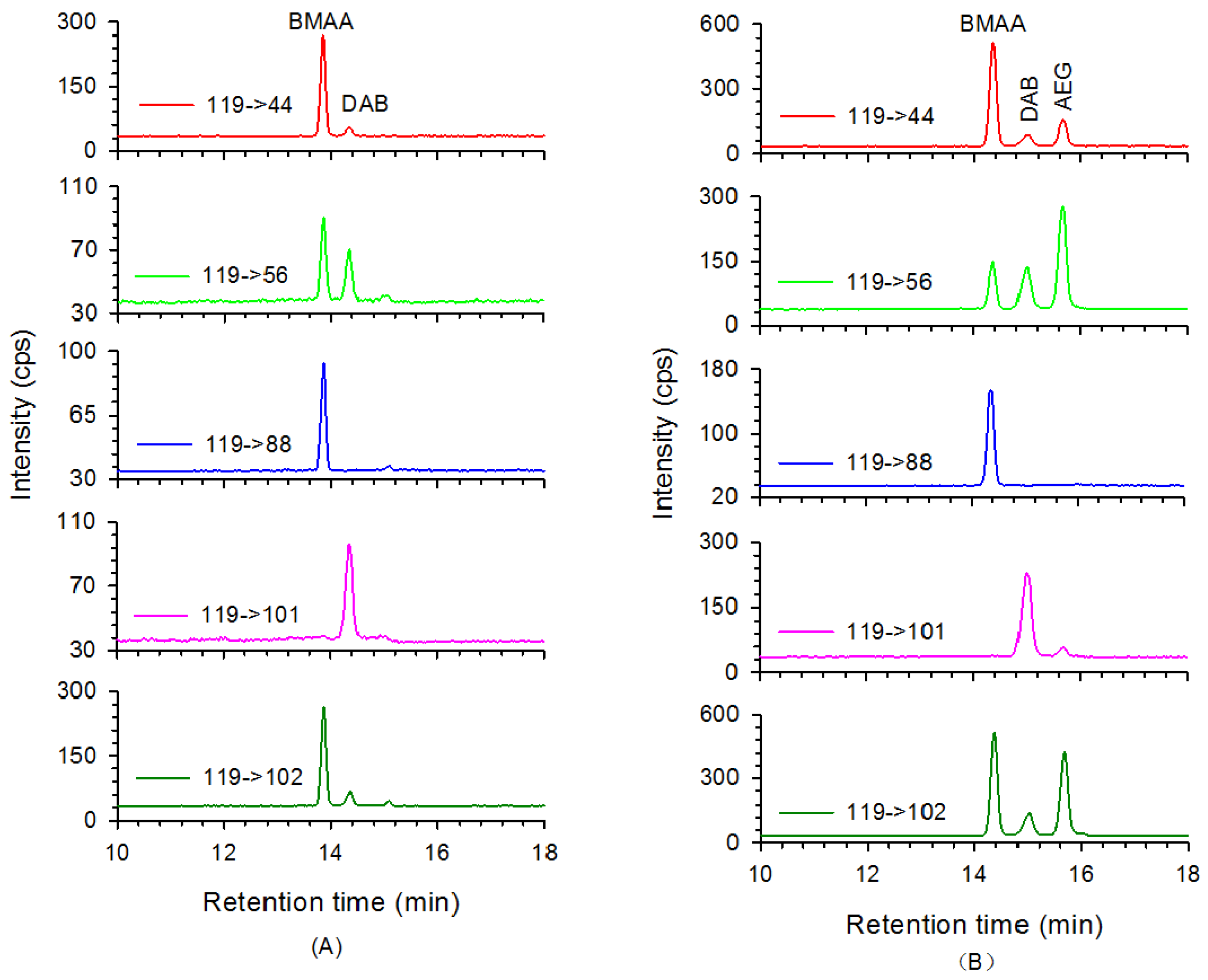
2. Results
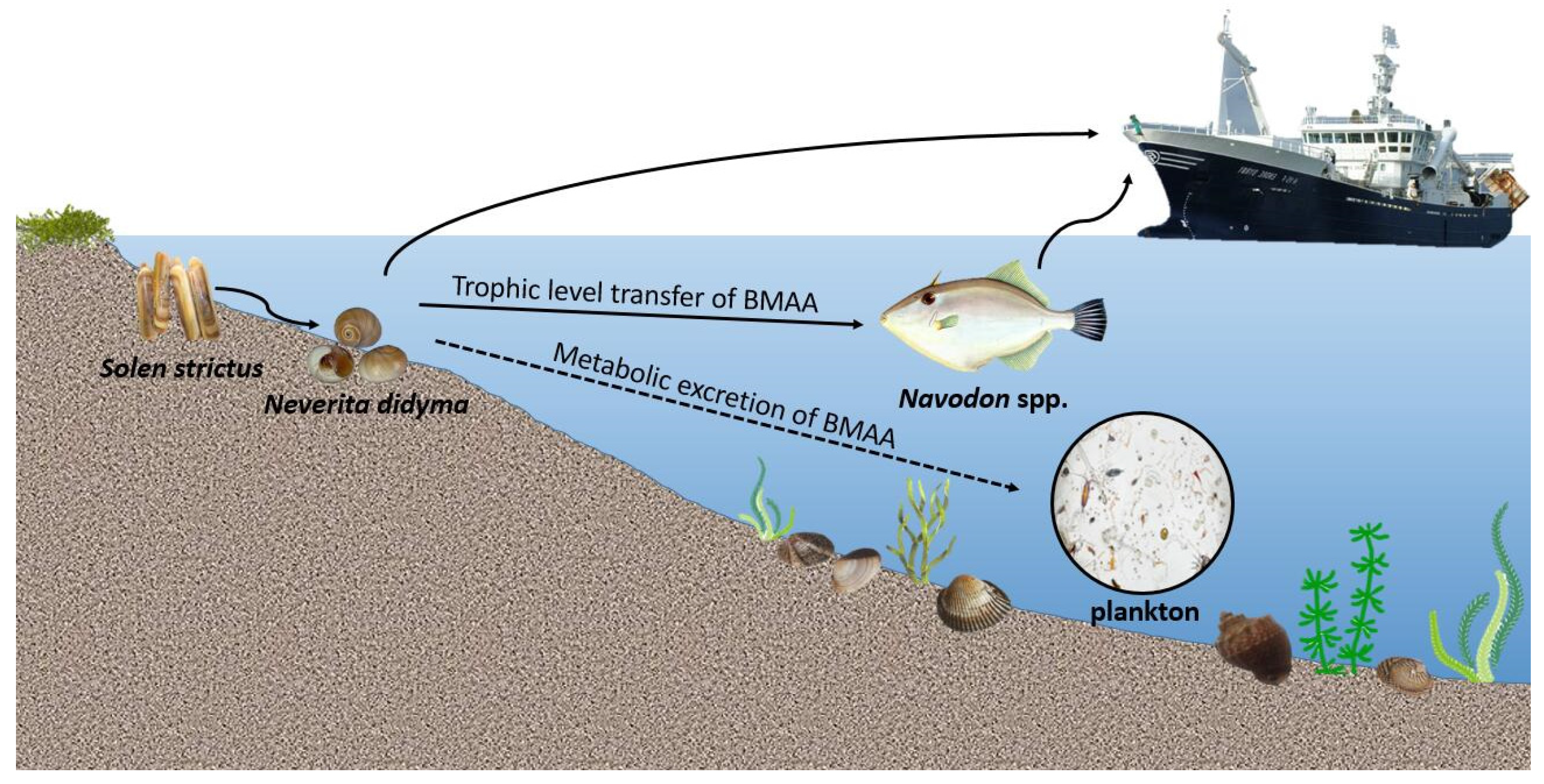
3. Discussion
4. Experimental Section
4.1. Chemicals
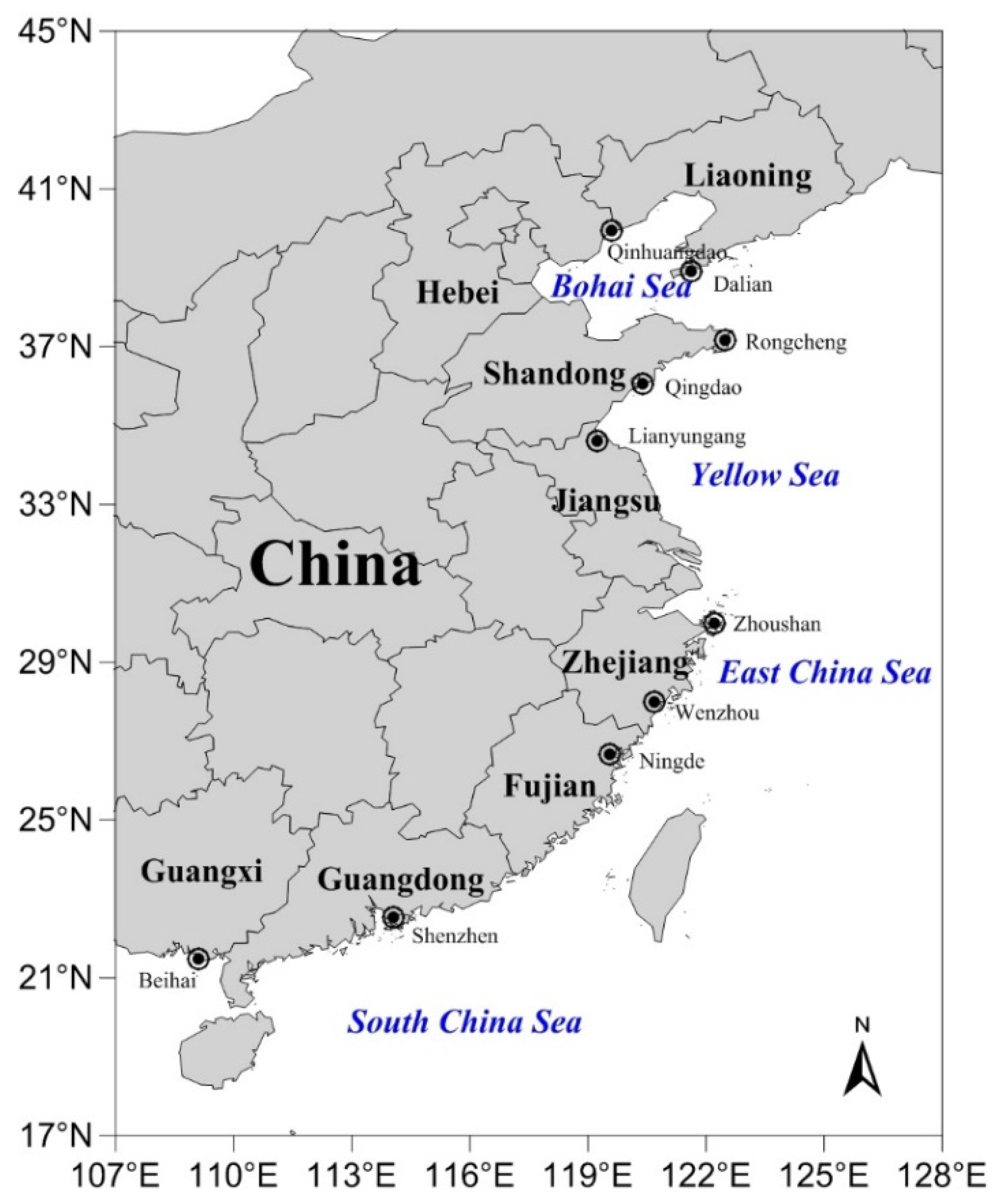
4.2. Samples and Sampling Locations
4.3. Sample Preparation
4.4. LC-MS/MS Analysis
4.5. Amino Acid Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cox, P.A.; Banack, S.A.; Murch, S.J. Biomagnification of cyanobacterial neurotoxins and neurodegenerative disease among the Chamorro people of Guam. Proc. Natl. Acad. Sci. USA 2003, 100, 13380–13383. [Google Scholar] [CrossRef] [PubMed]
- Murch, S.J.; Cox, P.A.; Banack, S.A.; Steele, J.C.; Sacks, O.W. Occurrence of β-methylamino-l-alanine (BMAA) in ALS/PDC patients from Guam. Acta Neurol. Scand. 2004, 110, 267–269. [Google Scholar] [CrossRef] [PubMed]
- Brownson, D.M.; Mabry, T.J.; Leslie, S.W. The cycad neurotoxic amino acid, β-N-methylamino-l-alanine (BMAA), elevates intracellular calcium levels in dissociated rat brain cells. J. Ethnopharmacol. 2002, 82, 159–167. [Google Scholar] [CrossRef]
- Nedeljkov, V.; Lopicic, S.; Pavlovic, D.; Cemerikic, D. Electrophysiological effect of β-N-methylamino-l-alanine on retzius nerve cells of the leech Haemopis sanguisuga. Ann. N. Y. Acad. Sci. 2005, 1048, 349–351. [Google Scholar] [CrossRef] [PubMed]
- Rao, S.D.; Banack, S.A.; Cox, P.A.; Weiss, J.H. BMAA selectively injures motor neurons via AMPA/kainate receptor activation. Exp. Neurol. 2006, 201, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Santiago, M.; Matarredona, E.R.; Machado, A.; Cano, J. Acute perfusion of BMAA in the rat’s striatum by in vivo microdialysis. Toxicol. Lett. 2006, 167, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Lobner, D.; Piana, P.M.T.; Salous, A.K.; Peoples, R.W. β-N-methylamino-l-alanine enhances neurotoxicity through multiple mechanisms. Neurobiol. Dis. 2007, 25, 360–366. [Google Scholar] [CrossRef] [PubMed]
- Esterhuizen-Londt, M.; Pflugmacher, S.; Downing, T.G. The effect of β-N-methylamino-l-alanine (BMAA) on oxidative stress response enzymes of the macrophyte Ceratophyllum demersum. Toxicon 2011, 57, 803–810. [Google Scholar] [CrossRef] [PubMed]
- Contardo-Jara, V.; Funke, M.S.; Peuthert, A.; Pflugmacher, S. β-N-methylamino-l-alanine exposure alters defense against oxidative stress in aquatic plants Lomariopsis lineata, Fontin alisantipyretica, Riccia fluitans and Taxiphyllum barbieri. Ecotoxicol. Environ. Saf. 2013, 88, 72–78. [Google Scholar] [CrossRef] [PubMed]
- Lürling, M.; Faassen, E.J.; Eenennaam, J.S.V. Effects of the cyanobacterial neurotoxin β-N-methylamino-l-alanine (BMAA) on the survival, mobility and reproduction of Daphnia magna. J. Plankton Res. 2011, 33, 333–342. [Google Scholar] [CrossRef]
- Esterhuizen-Londt, M.; Wiegand, C.; Downing, T.G. β-N-methylamino-l-alanine (BMAA) uptake by the animal model, Daphnia magna and subsequent oxidative stress. Toxicon 2015, 100, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Purdie, E.L.; Metcalf, J.S.; Kashmiri, S.; Codd, G.A. Toxicity of the cyanobacterial neurotoxin beta-N-methylamino-l-alanine to three aquatic animal species. Amyotroph. Lateral Scler. 2009, 10, 67–70. [Google Scholar] [CrossRef] [PubMed]
- Purdie, E.L.; Samsudin, S.; Eddy, F.B.; Codd, G.A. Effects of the cyanobacterial neurotoxin β-N-methylamino-l-alanine on the early-life stage development of zebrafish (Danio rerio). Aquat. Toxicol. 2009, 95, 279–284. [Google Scholar] [CrossRef] [PubMed]
- Cox, P.A.; Banack, S.A.; Murch, S.J. Diverse taxa of cyanobacteria produce β-N-methylamino-l-alanine, a neurotoxic amino acid. Proc. Natl. Acad. Sci. USA 2005, 102, 5074–5078. [Google Scholar] [CrossRef] [PubMed]
- Esterhuizen, M.; Downing, T.G. β-N-methylamino-l-alanine (BMAA) in novel South African cyanobacterial isolates. Ecotoxicol. Environ. Saf. 2008, 71, 309–313. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Ilag, L.L. Detection of endogenous BMAA in dinoflagellate (Heterocapsa triquetra) hints at evolutionary conservation and environmental concern. PubRaw Sci. 2014, 1, 1–8. [Google Scholar]
- Jiang, L.; Eriksson, J.; Lage, S.; Jonasson, S.; Shams, S.; Mehine, M.; Ilag, L.L.; Rasmussen, U. Diatoms: A novel source for the neurotoxin BMAA in aquatic environments. PLoS ONE 2014, 9, e84578. [Google Scholar] [CrossRef] [PubMed]
- Banack, S.A.; Johnson, H.E.; Cheng, R.; Cox, P.A. Production of the neurotoxin BMAA by a marine cyanobacterium. Mar. Drugs 2007, 5, 180–196. [Google Scholar] [CrossRef] [PubMed]
- Cervantes Cianca, R.C.; Baptista, M.S.; Lopes, V.R.; Vasconcelos, V.M. The non-protein amino acid β-N-methylamino-l-alanine in Portuguese cyanobacterial isolates. Amino Acids 2012, 42, 2473–2479. [Google Scholar] [CrossRef] [PubMed]
- Metcalf, J.S.; Banack, S.A.; Lindsay, J.; Morrison, L.F.; Cox, P.A.; Codd, G.A. Co-occurrence of β-N-methylamino-l-alanine, a neurotoxic amino acid with other cyanobacterial toxins in British waterbodies, 1990–2004. Environ. Microbiol. 2008, 10, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Johnson, H.E.; King, S.R.; Banack, S.A.; Webster, C.; Callanaupa, W.J.; Cox, P.A. Cyanobacteria (Nostoc commune) used as a dietary item in the Peruvian highlands produce the neurotoxic amino acid BMAA. J. Ethnopharmacol. 2008, 118, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Brand, L.E.; Pablo, J.; Compton, A.; Hammerschlag, N.; Mash, D.C. Cyanobacterial blooms and the occurrence of the neurotoxin, beta-N-methylamino-l-alanine (BMAA), in South Florida aquatic food webs. Harmful Algae 2010, 9, 620–635. [Google Scholar] [CrossRef] [PubMed]
- Jonasson, S.; Eriksson, J.; Berntzon, L.; Spáčil, Z.; Ilag, L.L.; Ronnevi, L.; Rasmussen, U.; Bergman, B. Transfer of a cyanobacterial neurotoxin within a temperate aquatic ecosystem suggests pathways for human exposure. Proc. Natl. Acad. Sci. USA 2010, 107, 9252–9257. [Google Scholar] [CrossRef] [PubMed]
- Masseret, E.; Banack, S.; Boumédiène, F.; Abadie, E.; Brient, L.; Pernet, F.; Juntas-Morales, R.; Pageot, N.; Metcalf, J.; Cox, P.; et al. Dietary BMAA exposure in an amyotrophic lateral sclerosis cluster from southern France. PLoS ONE 2013, 8, e83406. [Google Scholar] [CrossRef] [PubMed]
- Christensen, S.J.; Hemscheidt, T.K.; Trapido-Rosenthal, H.; Laws, E.A.; Bidigare, R.R. Detection and quantification of β-methylamino-l-alanine in aquatic invertebrates. Limnol. Oceanogr. Meth. 2012, 10, 891–898. [Google Scholar] [CrossRef]
- Mondo, K.; Hammerschlag, N.; Basile, M.; Pablo, J.; Banack, S.A.; Mash, D.C. Cyanobacterial neurotoxin β-N-Methylamino-l-alanine (BMAA) in shark fins. Mar. Drugs 2012, 10, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Mondo, K.; Glover, W.B.; Murch, S.J.; Liu, G.; Cai, Y.; Davis, D.A.; Mash, D.C. Environmental neurotoxins β-N-methylamino-l-alanine (BMAA) and mercury in shark cartilage dietary supplements. Food Chem. Toxicol. 2014, 70, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Lage, S.; Costa, P.R.; Moita, T.; Eriksson, J.; Rasmussen, U.; Rydberg, S.J. BMAA in shellfish from two Portuguese transitional water bodies suggests the marine dinoflagellate Gymnodinium catenatum as a potential BMAA source. Aquat. Toxicol. 2014, 152, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Chen, Q.; Chen, X.; Wang, X.; Liao, X.; Jiang, L.; Wu, J.; Yang, L. Occurrence and transfer of a cyanobacterial neurotoxin β-N-methylamino-l-alanine within the aquatic food webs of Gonghu Bay (Lake Taihu, China) to evaluate the potential human health risk. Sci. Total Environ. 2014, 468–469, 457–463. [Google Scholar] [CrossRef] [PubMed]
- Lage, S.; Annadotter, H.; Rasmussen, U.; Rydberg, S. Biotransfer of β-N-Methylamino-l-alanine (BMAA) in a eutrophicated freshwater lake. Mar. Drugs 2015, 13, 1185–1201. [Google Scholar] [CrossRef] [PubMed]
- Rosén, J.; Hellenäs, K. Determination of the neurotoxin BMAA (β-N-methylamino-l-alanine) in cycad seed and cyanobacteria by LC-MS/MS (liquid chromatography tandem mass spectrometry). Analyst 2008, 133, 1785–1789. [Google Scholar] [CrossRef] [PubMed]
- Krüger, T.; Mönch, B.; Oppenhäuser, S.; Luckas, B. LC-MS/MS determination of the isomeric neurotoxins BMAA (β-N-methylamino-l-alanine) and DAB (2,4-diaminobutyric acid) in cyanobacteria and seeds of Cycas revoluta and Lathyrus latifolius. Toxicon 2010, 55, 547–557. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Tian, Z.; Li, J.; Yu, R.; Banack, S.A.; Wang, Z. Detection of the neurotoxin BMAA within cyanobacteria isolated from freshwater in China. Toxicon 2010, 55, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Li, A.; Fan, H.; Ma, F.; McCarron, P.; Thomas, K.; Tang, X.; Quilliam, M.A. Elucidation of matrix effects and performance of solid-phase extraction for LC-MS/MS analysis of β-N-methylamino-l-alanine (BMAA) and 2,4-diaminobutyric acid (DAB) neurotoxins in cyanobacteria. Analyst 2012, 137, 1210–1219. [Google Scholar] [CrossRef] [PubMed]
- Réveillon, D.; Abadie, E.; Séchet, V.; Brient, L.; Savar, V.; Bardouil, M.; Hess, P.; Amzil, Z. Beta-N-methylamino-l-alanine: LC-MS/MS optimization, screening of cyanobacterial strains and occurrence in shellfish from Thau, a French mediterranean lagoon. Mar. Drugs 2014, 12, 5441–5467. [Google Scholar] [CrossRef] [PubMed]
- Beach, D.G.; Kerrin, E.S.; Quilliam, M.A. Selective quantitation of the neurotoxin BMAA by use of hydrophilic-interaction liquid chromatography-differential mobility spectrometry-tandem mass spectrometry (HILIC-DMS-MS/MS). Anal. Bioanal. Chem. 2015, 407, 8397–8409. [Google Scholar] [CrossRef] [PubMed]
- Réveillon, D.; Abadie, E.; Séchet, V.; Masseret, E.; Hess, P.; Amzil, Z. β-N-methylamino-l-alanine (BMAA) and isomers: Distribution in different food web compartments of Thau lagoon, French Mediterranean Sea. Mar. Environ. Res. 2015, 110, 8–18. [Google Scholar] [CrossRef] [PubMed]
- Réveillon, D.; Séchet, V.; Hess, P.; Amzil, Z. Systematic detection of BMAA (β-N-methylamino-l-alanine) and DAB (2,4-diaminobutyric acid) in mollusks collected in shellfish production areas along the French coasts. Toxicon 2016, 110, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Faassen, E.J. Presence of the neurotoxin BMAA in aquatic ecosystems: What do we really know? Toxins 2014, 6, 1109–1138. [Google Scholar] [CrossRef] [PubMed]
- Cox, P.A.; Davis, D.A.; Mash, D.C.; Metcalf, J.S.; Banack, S.A. Dietary exposure to an environmental toxin triggers neurofibrillary tangles and amyloid deposits in the brain. Proc. R. Soc. B 2016, 283, 20152397. [Google Scholar] [CrossRef] [PubMed]
- Dunlop, R.A.; Cox, P.A.; Banack, S.A.; Rodgers, K.J. The non-protein amino acid BMAA is misincorporated into human proteins in place of l-serine causing protein misfolding and aggregation. PLoS ONE 2013, 8, e75376. [Google Scholar] [CrossRef] [PubMed]
- Main, B.J.; Dunlop, R.A.; Rodgers, K.J. The use of l-serine to prevent β-methylamino-l-alanine (BMAA)-induced proteotoxic stress in vitro. Toxicon 2016, 109, 7–12. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sampling Location | Sampling Date | Mollusk Species | Toxin Concentration | |
|---|---|---|---|---|
| BMAA | DAB | |||
| Beihai City | 19 March 2016 | Atrina pectinata | nd | nd |
| Crassostrea sp. | nd | 0.53 | ||
| Perna viridis | nd | 0.87 | ||
| Antigona lamellaris | nd | 0.26 | ||
| Meretrix lusoria | nd | nd | ||
| Periglypta petechialis | nd | 0.16 | ||
| Zhoushan City | 22 March 2016 | Crassostrea sp. | nd | 0.23 |
| Mytilus coruscus | 0.45 | 1.61 | ||
| Mytilus galloprovincialis | nd | 0.71 | ||
| Rongcheng City | 23 March 2016 | Crassostrea sp. | nd | 0.23 |
| Chlamys farreri | nd | 0.81 | ||
| Mytilus galloprovincialis | nd | 0.50 | ||
| Solen strictus | 0.66 | nd | ||
| Mactra chinensis | nd | 0.47 | ||
| Ruditapes philippinarum | nd | 0.30 | ||
| Neverita didyma | 2.15 | 0.32 | ||
| Ningde City | 24 March 2016 | Atrina pectinata | nd | nd |
| Crassostrea sp. | nd | 0.61 | ||
| Mytilus galloprovincialis | nd | 0.41 | ||
| Sinonovacula constricta | nd | 0.17 | ||
| Tegillarca granosa | nd | nd | ||
| Periglypta petechialis | nd | nd | ||
| Haliotis discus hannai | nd | 0.25 | ||
| Turritella bacillum | nd | nd | ||
| Natica maculosa | nd | 0.15 | ||
| Batillaria zonalis | nd | nd | ||
| Ruditapes philippinarum | nd | 0.74 | ||
| Wenzhou City | 25 March 2016 | Crassostrea sp. | nd | 0.70 |
| Mytilus coruscus | nd | 0.63 | ||
| Sinonovacula constricta | nd | 0.39 | ||
| Tegillarca granosa | nd | 0.18 | ||
| Turritella bacillum | nd | nd | ||
| Moerella iridescens | nd | 0.051 | ||
| Dalian City | 25 March 2016 | Atrina pectinata | nd | 0.35 |
| Crassostrea sp. | nd | 0.22 | ||
| Chlamys farreri | nd | 0.67 | ||
| Mytilus galloprovincialis | nd | 0.29 | ||
| Scapharca subcrenata | nd | 0.55 | ||
| Mactra chinensis | nd | 0.57 | ||
| Ruditapes philippinarum | nd | 0.57 | ||
| Volutharpa ampullacea | nd | 2.65 | ||
| Neptunea cumingii | nd | 0.54 | ||
| Neverita didyma | 3.97 | 0.21 | ||
| Lianyungang City | 26 March 2016 | Crassostrea sp. | nd | 0.83 |
| Chlamys farreri | nd | 1.12 | ||
| Mytilus galloprovincialis | nd | 0.58 | ||
| Sinonovacula constricta | nd | 2.01 | ||
| Arca inflata | nd | 0.24 | ||
| Mercenaria mercenaria | nd | 0.17 | ||
| Ruditapes philippinarum | nd | 0.71 | ||
| Turritella bacillum | nd | nd | ||
| Rapana venosa | nd | nd | ||
| Neverita didyma | 0.99 | 0.40 | ||
| Qinhuangdao City | 26 March 2016 | Crassostrea sp. | nd | 0.45 |
| Argopecten irradians | nd | nd | ||
| Mytilus galloprovincialis | nd | 0.74 | ||
| Rapana venosa | nd | 0.29 | ||
| Arca inflata | nd | 1.02 | ||
| Shenzhen City | 28 March 2016 | Crassostrea sp. | nd | nd |
| Mimachlamys nobilis | nd | nd | ||
| Perna viridis | nd | 0.31 | ||
| Gafrarium tumidum | nd | 1.00 | ||
| Qingdao City | 31 March 2016 | Crassostrea sp. | nd | 0.31 |
| Chlamys farreri | nd | 1.07 | ||
| Mytilus galloprovincialis | nd | 0.60 | ||
| Haliotis discus hannai | nd | nd | ||
| Rapana venosa | nd | 0.16 | ||
| Ruditapes philippinarum | nd | 0.70 | ||
| Laizhou City | 26 April 2016 | Neverita didyma | 0.86 | 0.30 |
| Qingdao City | 22 May 2016 | Neverita didyma | 1.43 | 0.39 |
| Standard Solution | Spiked Sample | Sample Extract | |||||||
|---|---|---|---|---|---|---|---|---|---|
| BMAA | DAB | AEG | BMAA | DAB | AEG | BMAA | DAB | ||
| Transitions | 119 → 44 | 41.6 ± 0.7 | 12.5 ± 0.3 | 15.1 ± 0.3 | 40.7 | 11.5 | 15.6 | 40.4 ± 0.5 | 12.1 ± 0.6 |
| 119 → 56 | 8.4 ± 0.2 | 21.9 ± 0.2 | 32.4 ± 0.2 | 9.3 | 22.0 | 32.0 | 10.1 ± 0.6 | 21.8 ± 1.6 | |
| 119 → 88 | 10.3 ± 0.2 | 0 | 0 | 10.1 | 0.0 | 0.0 | 10.1 ± 0.6 | 0 | |
| 119 → 101 | 0 | 43.6 ± 0.2 | 2.4 ± 0.2 | 0.0 | 44.2 | 2.6 | 0 | 44.7 ± 1.0 | |
| 119 → 102 | 39.7 ± 0.3 | 22.1 ± 0.3 | 50.1 ± 0.3 | 39.9 | 22.3 | 49.8 | 39.4 ± 0.9 | 21.4 ± 1.2 | |
| Retention times | 15.33 | 16.27 | 16.97 | 14.36 | 15.01 | 15.67 | 14.32 | 14.99 | |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, A.; Song, J.; Hu, Y.; Deng, L.; Ding, L.; Li, M. New Typical Vector of Neurotoxin β-N-Methylamino-l-Alanine (BMAA) in the Marine Benthic Ecosystem. Mar. Drugs 2016, 14, 202. https://doi.org/10.3390/md14110202
Li A, Song J, Hu Y, Deng L, Ding L, Li M. New Typical Vector of Neurotoxin β-N-Methylamino-l-Alanine (BMAA) in the Marine Benthic Ecosystem. Marine Drugs. 2016; 14(11):202. https://doi.org/10.3390/md14110202
Chicago/Turabian StyleLi, Aifeng, Jialiang Song, Yang Hu, Longji Deng, Ling Ding, and Meihui Li. 2016. "New Typical Vector of Neurotoxin β-N-Methylamino-l-Alanine (BMAA) in the Marine Benthic Ecosystem" Marine Drugs 14, no. 11: 202. https://doi.org/10.3390/md14110202
APA StyleLi, A., Song, J., Hu, Y., Deng, L., Ding, L., & Li, M. (2016). New Typical Vector of Neurotoxin β-N-Methylamino-l-Alanine (BMAA) in the Marine Benthic Ecosystem. Marine Drugs, 14(11), 202. https://doi.org/10.3390/md14110202