Chemical Profiling and Bioactivity of Body Wall Lipids from Strongylocentrotus droebachiensis

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Fatty Acid Composition of Body Wall (BW) Lipids
2.2. Sterols and Alkoxyglycerols (AOG)
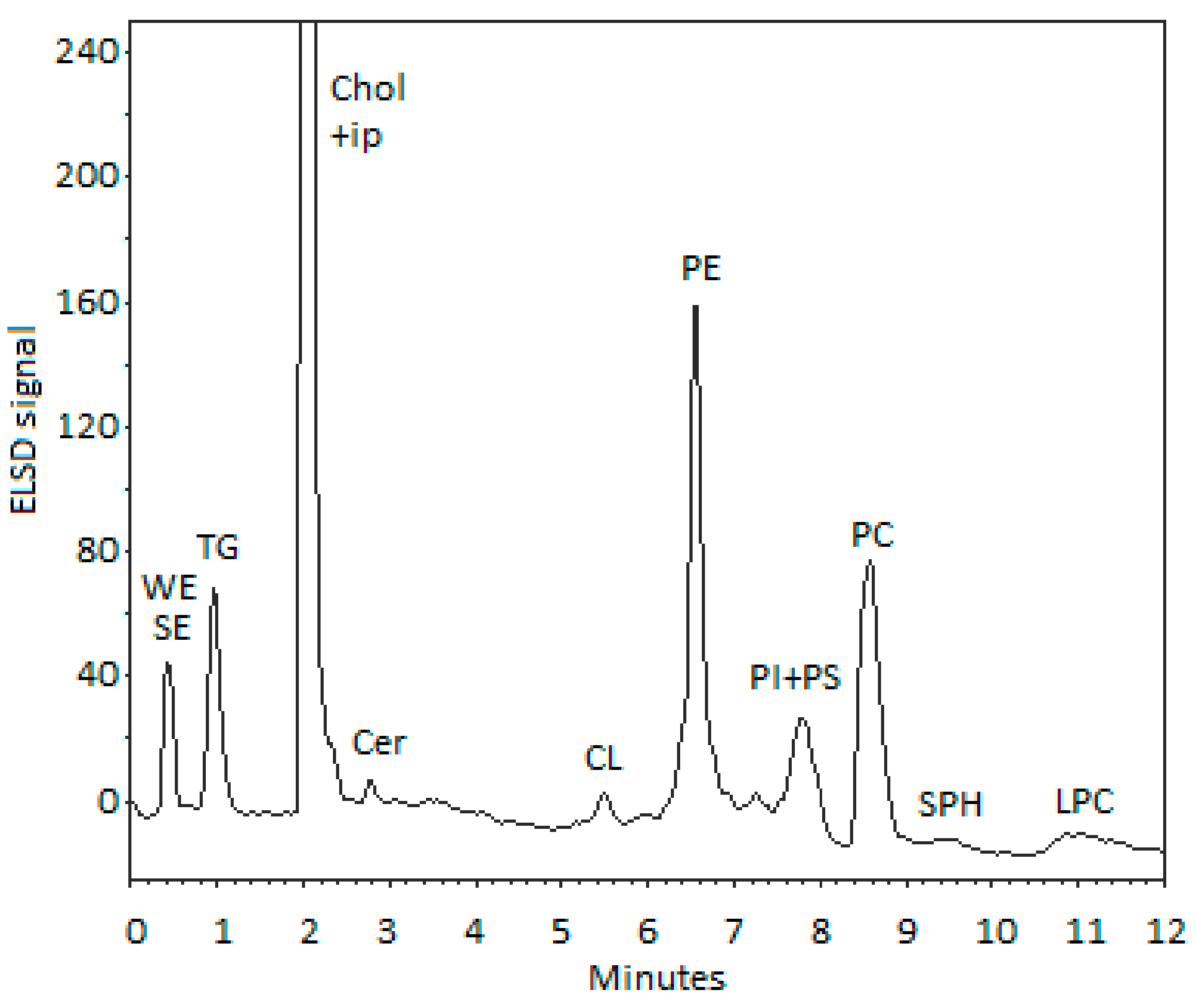
2.3. Lipid Classes
2.4. Bioactivity of Body Wall (BW) Lipids
3. Discussion
4. Material and Methods
4.1. Sample Preparation and Extraction of Lipids
4.2. Chemical Analyses
4.2.1. Analysis of Fatty Acids, Sterols and Alkoxyglycerols by GC-MS
4.2.2. Analysis of Lipid Classes by UPLC-ELSD
4.3. Biological Assays
4.3.1. Cell Lines and Cell Culture
4.3.2. Western Blotting
4.3.3. Assessment of Cyclooxygenase Activity
4.4. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- FAO. Global Production Statistics (1950–2015). Available online: http://www.fao.org/fishery/statistics/global-production/query/en (accessed on 21 June 2017).
- Liyana-Pathirana, C.; Shahidi, F.; Whittick, A.; Hooper, R. Lipid and lipid soluble components of gonads of green sea urchin (Strongylocentrotus droebachiensis). J. Food Lipids 2002, 9, 105–126. [Google Scholar] [CrossRef]
- Kalogeropoulos, K.; Mikellidi, A.; Nomikos, T.; Chiou, A. Screening of macro- and bioactive micro-constituents of commercial finfish and sea urchin eggs. LWT-Food Sci. Technol. 2012, 46, 525–531. [Google Scholar] [CrossRef]
- Pozharitskaya, O.N.; Shikov, A.N.; Laakso, I.; Seppänen-Laakso, T.; Makarenko, I.E.; Faustova, N.M.; Makarova, M.N.; Makarov, V.G. Bioactivity and chemical characterization of gonads of green sea urchin Strongylocentrotus droebachiensis from Barents Sea. J. Funct. Foods 2015, 17, 227–234. [Google Scholar] [CrossRef]
- Matsuno, T.; Tsushima, M. Carotenoids in sea urchins. Dev. Aquac. Fish. Sci. 2001, 32, 115–138. [Google Scholar]
- Dincer, T.; Cakli, S. Chemical composition and biometrical measurements of the Turkish sea urchin (Paracentrotus lividus, Lamarck, 1816). Crit. Rev. Food Sci. Nutr. 2007, 27, 21–26. [Google Scholar] [CrossRef]
- Shikov, A.N.; Laakso, I.; Pozharitskaya, O.N.; Makarov, V.G.; Hiltunen, R. Phospholipids and amino-acid composition of eggs of sea urchin from Barents Sea. Planta Med. 2012, 78, 1146. [Google Scholar] [CrossRef]
- Pozharitskaya, O.N.; Shikov, A.N.; Makarova, M.N.; Ivanova, S.A.; Kosman, V.M.; Makarov, V.G.; Bazgier, V.; Berka, K.; Otyepka, M.; Ulrichová, J. Antiallergic effects of pigments isolated from green sea urchin (Strongylocentrotus droebachiensis) shells. Planta Med. 2013, 79, 1698–1704. [Google Scholar] [CrossRef] [PubMed]
- Kovaleva, M.A.; Ivanova, S.A.; Makarova, M.N.; Pozharitskaya, O.N.; Shikov, A.N.; Makarov, V.G. Effect of a complex preparation of sea urchin shells on blood glucose level and oxidative stress parameters in type II diabetes model. Eksp. Klin. Farmakol. 2013, 76, 27–30. [Google Scholar] [PubMed]
- Agafonova, I.G.; Bogdanovich, R.N.; Kolosova, N.G. Assessment of nephroprotective potential of histochrome during induced arterial hypertension. Bull. Exp. Biol. Med. 2015, 160, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Talalaeva, O.S.; Mishchenko, N.P.; Bryukhanov, V.M.; Zverev, Y.F.; Fedoreyev, S.A.; Lampatov, V.V.; Zharikov, A.Y. The influence of histochrome on exudative and proliferative phases of the experimental inflammation. Bull. Sib. Branch RAMS 2012, 32, 28–31. [Google Scholar]
- Kuwahara, R.; Hatate, H.; Yuki, T.; Murata, H.; Tanaka, R.; Hama, Y. Antioxidant property of polyhydroxylated naphthoquinone pigments from shells of purple sea urchin Anthocidaris crassispina. LWT-Food Sci. Technol. 2009, 42, 1296–1300. [Google Scholar] [CrossRef]
- Pozharitskaya, O.N.; Ivanova, S.A.; Shikov, A.N.; Makarov, V.G. Evaluation of free radical-scavenging activity of sea urchin pigments using HPTLC with post-chromatographic derivatization. Chromatographia 2013, 76, 1353–1358. [Google Scholar] [CrossRef]
- Powell, C.; Hughes, A.D.; Kelly, M.S.; Conner, S.; McDougall, G.J. Extraction and identification of antioxidant polyhydroxynaphthoquinone pigments from the sea urchin, Psammechinus miliaris. LWT-Food Sci. Technol. 2014, 59, 455–460. [Google Scholar] [CrossRef]
- Anufriev, V.P.; Novikov, V.L.; Maximov, O.B.; Elyakov, G.B.; Levitsky, D.O.; Lebedev, A.V.; Sadretdinov, S.M.; Shvilkin, A.V.; Afonskaya, N.I.; Ruda, M.Y.; et al. Synthesis of some hydroxynaphthazarins and their cardioprotective effects under ischemia-reperfusion in vivo. Bioorg. Med. Chem. Lett. 1998, 8, 587–592. [Google Scholar] [CrossRef]
- Lakeev, Y.V.; Kosykh, V.A.; Kosenkov, E.I.; Novikov, V.L.; Lebedev, A.V.; Repin, V.S. Effect of natural and synthetic antioxidants (polyhydroxynaphthaquinones) on cholesterol metabolism in cultured rabbit hepatocytes. Bull. Exp. Biol. Med. 1992, 114, 1611–1614. [Google Scholar] [CrossRef]
- Amarowicz, R.; Synowiecki, J.; Shahidi, F. Chemical composition of shells from red (Strongylocentrotus franciscanus) and green (Strongylocentrotus droebachiensis) sea urchin. Food Chem. 2012, 133, 822–826. [Google Scholar] [CrossRef]
- Krishtopina, A.S.; Urakova, I.N.; Pozharitskaya, O.N.; Razboeva, E.V.; Kosman, V.M.; Makarov, V.G.; Shikov, A.N. Optimization of polyhydroxynaphtoquinone extraction from shells of Strongylocentrotus droebachiensis sea urchins. Pharm. Chem. J. 2017, 51, 407–410. [Google Scholar] [CrossRef]
- Phillips, K.M.; Ruggio, D.M.; Exler, J.; Patterson, K.Y. Sterol composition of shellfish species commonly consumed in the United States. Food Nutr. Res. 2012, 56, 19831. [Google Scholar] [CrossRef] [PubMed]
- Bordier, C.G.; Sellier, N.; Foucault, A.P.; Le Goffic, F. Purification and characterization of deep sea shark Centrophorus squamosus liver oil 1-O-alkylglycerol ether lipids. Lipids 1996, 31, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. n-3 polyunsaturated fatty acids, inflammation, and inflammatory diseases. Am. J. Clin. Nutr. 2006, 83, S1505–S1519. [Google Scholar]
- Takagi, T.; Eaton, C.A.; Ackman, R.G. Distribution of fatty acids in lipids of the common Atlantic sea urchin Strongylocentrotus droebachiensis. Can. J. Fish. Aquat. Sci. 1980, 37, 195–202. [Google Scholar] [CrossRef]
- Castell, J.D.; Kennedy, E.J.; Robinson, S.M.C.; Parsons, G.J.; Blair, T.J.; Gonzales-Duran, E. Effect of dietary lipids on fatty acid composition and metabolism in juvenile green sea urchin (Strongylocentrotus droebachiensis). Aquaculture 2004, 242, 417–436. [Google Scholar] [CrossRef]
- Lands, W.E.M. Diets could prevent many diseases. Lipids 2003, 38, 317–321. [Google Scholar] [CrossRef] [PubMed]
- Lands, B. A critique of paradoxes in current advice on dietary lipids. Prog. Lipid Res. 2008, 47, 77–106. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Kim, S.C.; Yu, T.; Yi, Y.S.; Rhee, M.H.; Sung, G.H.; Yoo, B.C.; Cho, J.Y. Functional roles of p38 mitogen-activated protein kinase in macrophage-mediated inflammatory responses. Mediat. Inflamm. 2014. [Google Scholar] [CrossRef] [PubMed]
- Ait-Said, F.; Elalamy, I.; Werts, C.; Gomard, M.T.; Jacquemin, C.; Couetil, J.P.; Hatmi, M. Inhibition by eicosapentaenoic acid of IL-1β-induced PGHS-2 expression in human microvascular endothelial cells: Involvement of lipoxygenase-derived metabolites and p38 MAPK pathway. Biochim. Biophys. Acta 2003, 1631, 77–84. [Google Scholar] [CrossRef]
- Moon, D.O.; Kim, K.C.; Jin, C.Y.; Han, M.H.; Park, C.; Lee, K.J.; Park, Y.M.; Choi, Y.H.; Kim, G.Y. Inhibitory effects of eicosapentaenoic acid on lipopolysaccharide-induced activation in BV2 microglia. Int. Immunopharmacol. 2007, 7, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Bagga, D.; Wang, L.; Farias-Eisner, R.; Glaspy, J.A.; Reddy, S.T. Differential effects of prostaglandin derived from ω-6 and ω-3 polyunsaturated fatty acids on COX-2 expression and IL-6 secretion. Proc. Natl. Acad. Sci. USA 2003, 100, 1751–1756. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, H.J.; Chang, K.C.; Baek, J.C.; Park, J.K.; Shin, J.K.; Choi, W.J.; Lee, J.H.; Paik, W.Y. DHA and EPA down-regulate COX-2 expression through suppression of NF-κB activity in LPS-treated human umbilical vein endothelial cells. Korean J. Physiol. Pharmacol. 2009, 13, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Seppänen-Laakso, T.; Laakso, I.; Hiltunen, R. Rapid determination of unsubstituted alkylglyceryl ethers by gas chromatography-mass spectrometry. J. Chromatogr. 1990, 530, 94–101. [Google Scholar] [CrossRef]
- Pugliese, P.T.; Jordan, K.; Cederberg, H.; Brohult, J. Some biological actions of alkylglycerols from shark liver oil. J. Altern. Complement. Med. 1998, 4, 87–99. [Google Scholar] [CrossRef] [PubMed]
- Pedrono, F.; Martin, B.; Leduc, C.; Le Lan, J.; Saïag, B.; Legrand, P.; Moulinoux, J.P.; Legrand, A.B. Natural alkylglycerols restrain growth and metastasis of grafted tumors in mice. Nutr. Cancer 2004, 48, 64–69. [Google Scholar] [CrossRef] [PubMed]
- Iannitti, T.; Palmieri, B. An update on the therapeutic role of alkylglycerols. Mar. Drugs 2010, 8, 2267–2300. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Sun, S.; Tang, N.; Cai, W.; Qian, L. Oral administration of alkylglycerols differentially modulates high-fat diet-induced obesity and insulin resistance in mice. Evid.-Based Complement. Altern. 2013, 2013, 834027. [Google Scholar] [CrossRef] [PubMed]
- Seppänen-Laakso, T.; Laakso, I.; Hiltunen, R. Analysis of fatty acids by gas chromatography, and its relevance to research on health and nutrition. Anal. Chim. Acta 2002, 465, 39–62. [Google Scholar] [CrossRef]
- Seppänen-Laakso, T.; Laakso, I.; Vanhanen, H.; Kiviranta, K.; Lehtimäki, T.; Hiltunen, R. Major human plasma lipid classes determined by quantitative high-performance liquid chromatography, their variation and associations with phospholipid fatty acids. J. Chromatogr. B 2001, 754, 437–445. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Fatty Acids | Bound Fatty Acids as FAME | % | FFA | % |
|---|---|---|---|---|
| C10:0–C13:0 | 0.4 ± 0.1 | 0.3 | - | - |
| C14:0 | 15.1 ± 0.7 | 9.6 | 3.7 ± 0.1 | 8.4 |
| C15:0 | 1.5 ± 0.1 | 0.9 | 0.5 ± 0.1 | 1.1 |
| C16:0 | 21.9 ± 1.2 | 13.9 | 5.0 ± 0.2 | 11.4 |
| C18:0 | 4.0 ± 0.2 | 2.5 | 2.1 ± 0.1 | 4.8 |
| C19:0 | 1.0 ± 0.1 | 0.6 | - | - |
| C20:0 | 0.6 ± 0.1 | 0.4 | - | - |
| Σ SaFAΣ | 44.5 ± 2.1 | 28.2 | 11.3 ± 0.3 | 25.7 |
| C14:1n5 | 1.1 ± 0.1 | 0.7 | - | - |
| C16:1n9 | 0.9 ± 0.1 | 0.6 | - | - |
| C16:1n7 | 7.8 ± 0.6 | 4.9 | 1.9 ± 0.1 | 4.3 |
| C16:1n5 | 4.5 ± 0.2 | 2.8 | - | - |
| C18:1n9 | 3.0 ± 0.2 | 1.9 | 1.4 ± 0.1 | 3.2 |
| C18:1n7 | 4.2 ± 0.2 | 2.7 | 2.9 ± 0.1 | 6.6 |
| C20:1n15 | 16.7 ± 0.8 | 10.6 | - | - |
| C20:1n9 | 5.2 ± 0.3 | 3.3 | 8.2 ± 0.4 | 18.6 |
| C20:1n7 | 1.3 ± 0.1 | 0.8 | - | - |
| C22:1n9 | 4.3 ± 0.2 | 2.7 | - | - |
| Σ MUFA | 49.0 ± 1.3 | 31.0 | 14.4 ± 0.4 | 32.7 |
| 20:2Δ5,11 | 12.2 ±1.2 | 7.7 | - | - |
| 20:2Δ5,13 | 3.0 ± 0.5 | 1.9 | - | - |
| 22:2Δ7,13 | 0.9 ± 0.2 | 0.6 | - | - |
| 22:2Δ7,15 | 4.7 ± 0.4 | 3.0 | - | - |
| Σ NMID | 20.9 ± 1.9 | 13.2 | - | - |
| C18:2n6 | 1.5 ± 0.1 | 0.9 | 5.2 ± 0.2 | 11.8 |
| C20:2n6 | 1.9 ± 0.1 | 1.2 | - | - |
| C20:3n6 | 0.9 ± 0.1 | 0.6 | - | - |
| C20:4n6 | 15.8 ± 0.6 | 10.0 | 7.3 ± 0.1 | 16.6 |
| Σ n6 PUFA | 20.1 ± 1.0 | 12.7 | 12.5 ± 0.8 | 28.4 |
| C18:3n3 | 1.2 ± 0.1 | 0.8 | - | - |
| C18:4n3 | 2.3 ± 0.1 | 1.5 | - | - |
| C20:3n3 | 2.3 ± 0.2 | 1.5 | - | - |
| C20:4n3 | 0.6 ± 0.1 | 0.4 | - | - |
| C20:5n3 | 15.2 ± 0.7 | 9.6 | 5.8 ± 1.0 | 13.2 |
| C22:5n3 | 0.2 ± 0.1 | 0.1 | - | - |
| C22:6n3 | 1.7 ± 0.1 | 1.1 | - | - |
| Σ n3 PUFA | 23.5 ± 1.1 | 14.9 | 5.8 ± 1.0 | 13.2 |
| Σ Fatty acids | 158.0 ± 5.3 | 100.0 | 44.0 ± 1.8 | 100.0 |
| n6/n3 PUFA | 0.86 | 2.16 |
| Sterols and AOGs | μg/mg |
|---|---|
| Cholesterol | 50.3 ± 2.8 |
| Non-cholesterol sterols * | 10.6 ± 0.5 |
| C16:0-AOG | 1.0 ± 0.1 |
| C18:1-AOG | 0.4 ± 0.1 |
| C18:0-AOG | 0.3 ± 0.1 |
| Σ Alkoxyglycerols | 1.7 ± 0.2 |
| Lipid Classes | μg/mg | % |
|---|---|---|
| WE + SE | 14.6 ± 0.7 | 8.4 |
| TG | 15.9 ± 1.0 | 9.1 |
| Σ Neutral lipids | 30.5 ± 1.6 | 17.5 |
| PE | 38.6 ± 1.1 | 22.2 |
| PI + PS | 27.8 ± 0.1 | 16.0 |
| PC | 63.3 ± 1.7 | 36.4 |
| LPC | 13.5 ± 0.5 | 7.8 |
| Σ Phospholipids | 143.2 ± 0.7 | 82.5 |
| Σ Total lipids | 173.7 ± 2.3 | 100.0 |
| Sample, Concentration | Percentage of MAPK p38 (%) |
|---|---|
| Intact cells (no stimulation with LPS) | 23.0 ± 1.2 |
| Control cells stimulated with LPS (1 μg/mL) | 100 |
| SB203580 (1.88 μg/mL) + LPS | 30.0 ± 1.7 |
| BWL (10 μg/mL) + LPS | 59.0 ± 1.3 |
| BWL (5 μg/mL) + LPS | 53.0 ± 1.9 |
| BWL (1 μg/mL) + LPS | 49.0 ± 0.9 |
| BWL (0.5 μg/mL) + LPS | 38.0 ± 1.6 |
| BWL (0.1 μg/mL) + LPS | 17.0 ± 1.5 |
| BWL (0.033 μg/mL) + LPS | 12.0 ± 0.5 |
| BWL (0.011 μg/mL) + LPS | 21.0 ± 1.7 |
| BWL (0.0037μg/mL) + LPS | 27.0 ± 0.7 |
| BWL (0.0012 μg/mL) + LPS | 38.0 ± 1.6 |
| BWL (0.0004 μg/mL) + LPS | 52.0 ± 1.2 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shikov, A.N.; Laakso, I.; Pozharitskaya, O.N.; Seppänen-Laakso, T.; Krishtopina, A.S.; Makarova, M.N.; Vuorela, H.; Makarov, V. Chemical Profiling and Bioactivity of Body Wall Lipids from Strongylocentrotus droebachiensis. Mar. Drugs 2017, 15, 365. https://doi.org/10.3390/md15120365
Shikov AN, Laakso I, Pozharitskaya ON, Seppänen-Laakso T, Krishtopina AS, Makarova MN, Vuorela H, Makarov V. Chemical Profiling and Bioactivity of Body Wall Lipids from Strongylocentrotus droebachiensis. Marine Drugs. 2017; 15(12):365. https://doi.org/10.3390/md15120365
Chicago/Turabian StyleShikov, Alexander N., Into Laakso, Olga N. Pozharitskaya, Tuulikki Seppänen-Laakso, Anna S. Krishtopina, Marina N. Makarova, Heikki Vuorela, and Valery Makarov. 2017. "Chemical Profiling and Bioactivity of Body Wall Lipids from Strongylocentrotus droebachiensis" Marine Drugs 15, no. 12: 365. https://doi.org/10.3390/md15120365
APA StyleShikov, A. N., Laakso, I., Pozharitskaya, O. N., Seppänen-Laakso, T., Krishtopina, A. S., Makarova, M. N., Vuorela, H., & Makarov, V. (2017). Chemical Profiling and Bioactivity of Body Wall Lipids from Strongylocentrotus droebachiensis. Marine Drugs, 15(12), 365. https://doi.org/10.3390/md15120365