The Marine Fungi Rhodotorula sp. (Strain CNYC4007) as a Potential Feed Source for Fish Larvae Nutrition

Abstract
:1. Introduction
2. Results
2.1. RNA/DNA Ratio as a Proxy for the Nutritional Condition of Danio rerio
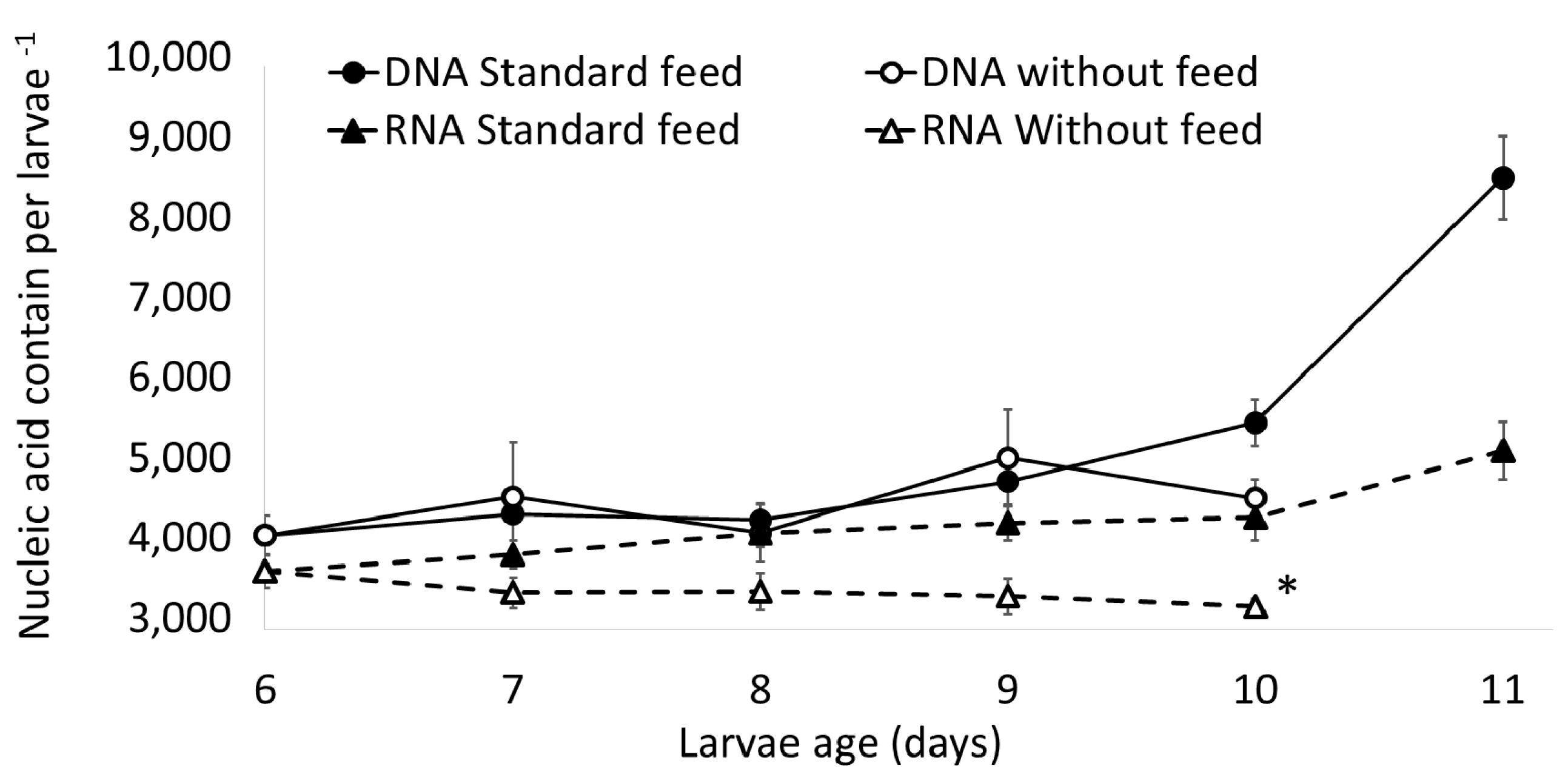
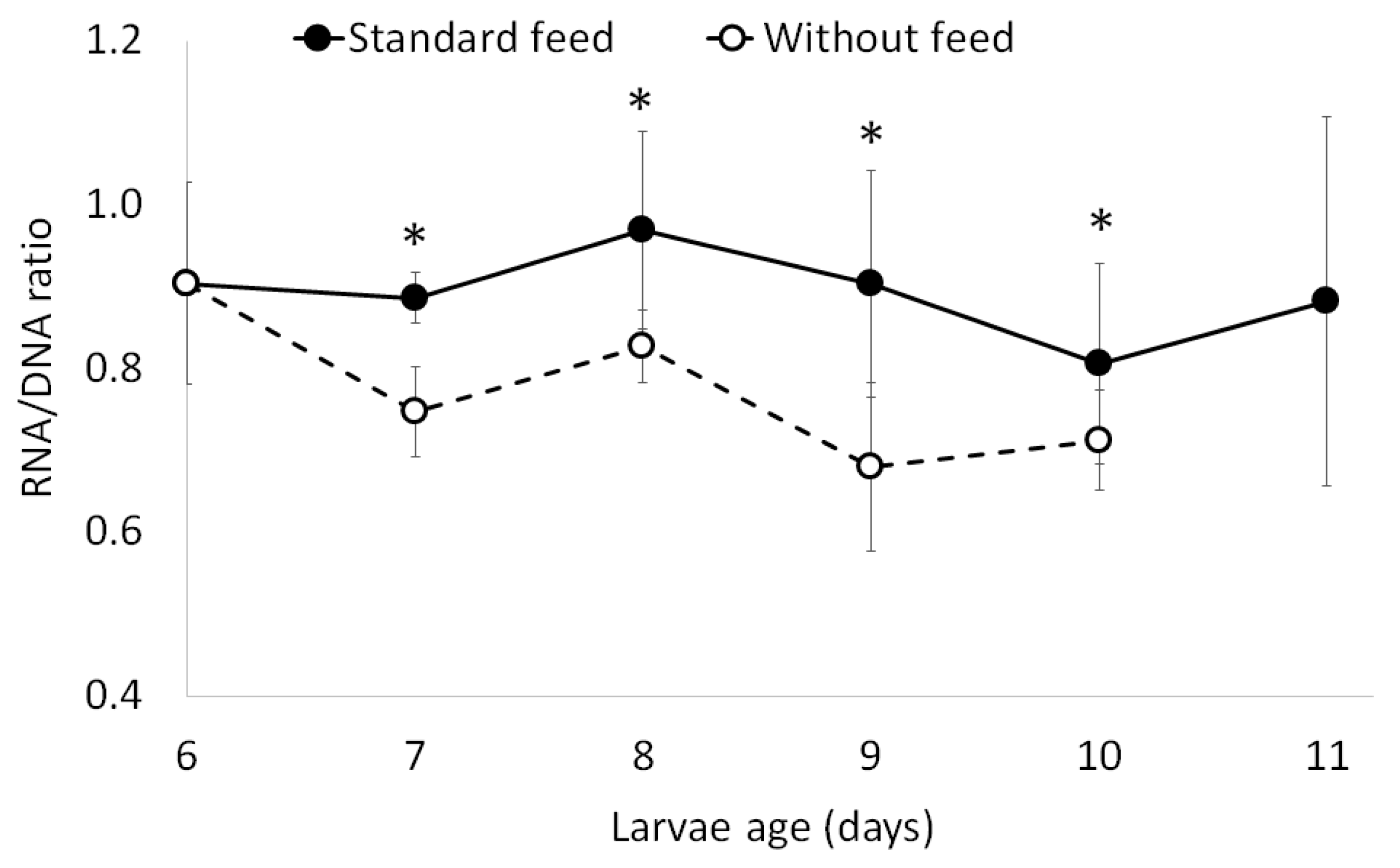
2.2. Assay with Six-Day-Old Larvae (First-Feeding Larvae, One Day Post-Yolk Absorption)
2.3. Feeding Six-Day-Old Larvae (Post-Yolk Absorption) with Encapsulation in Brachionus sp.
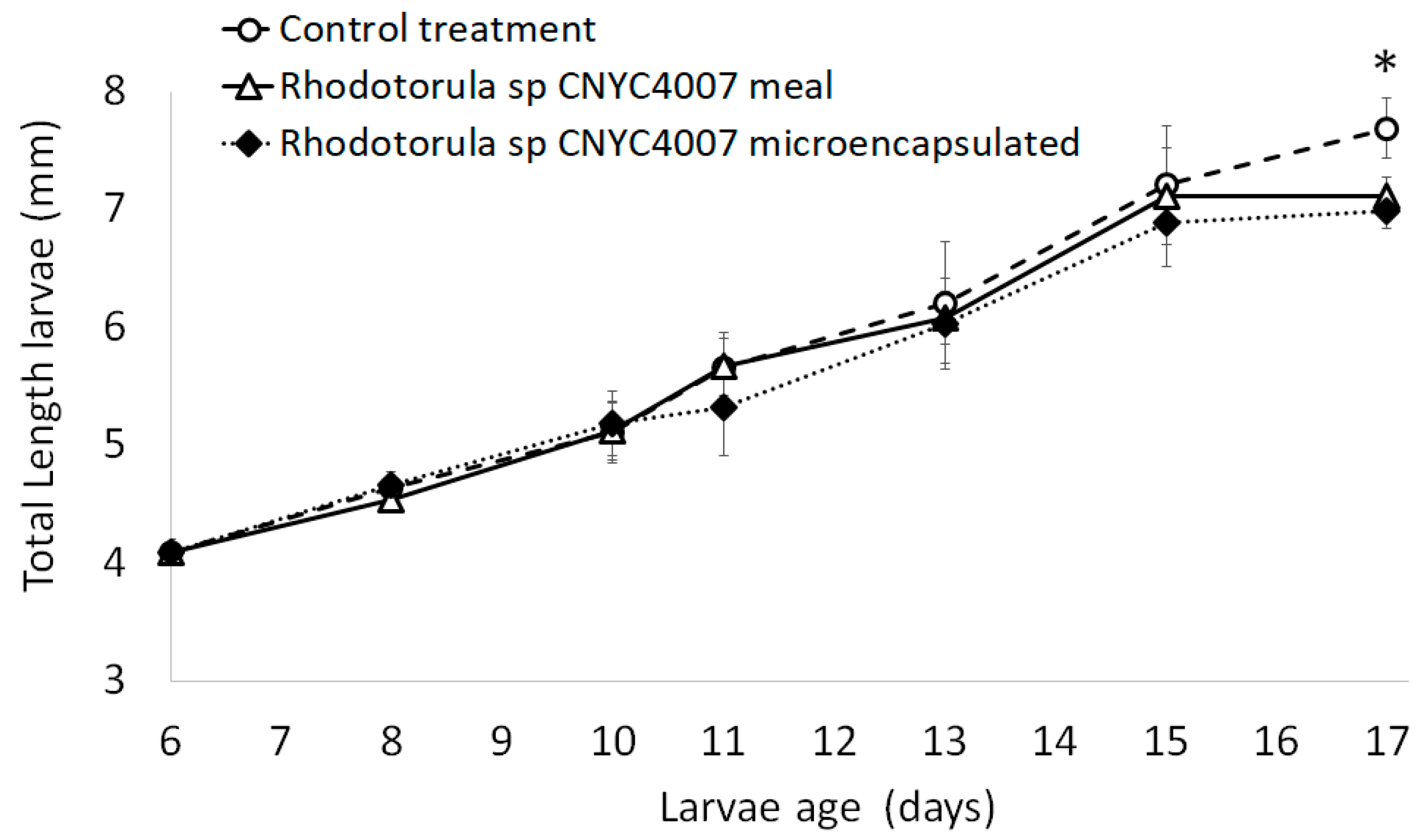
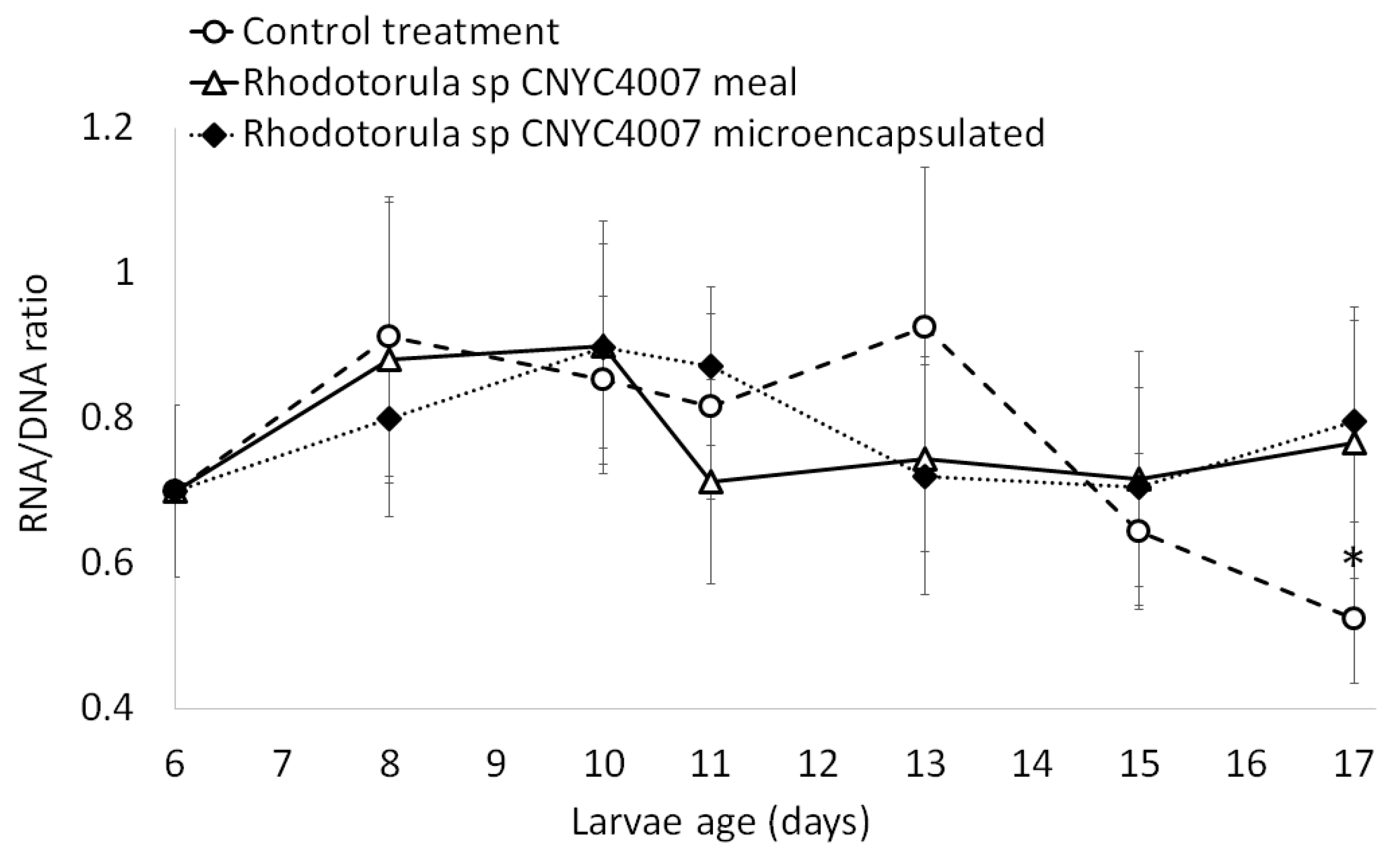
2.4. Experiments with 12-Day-Old Larvae
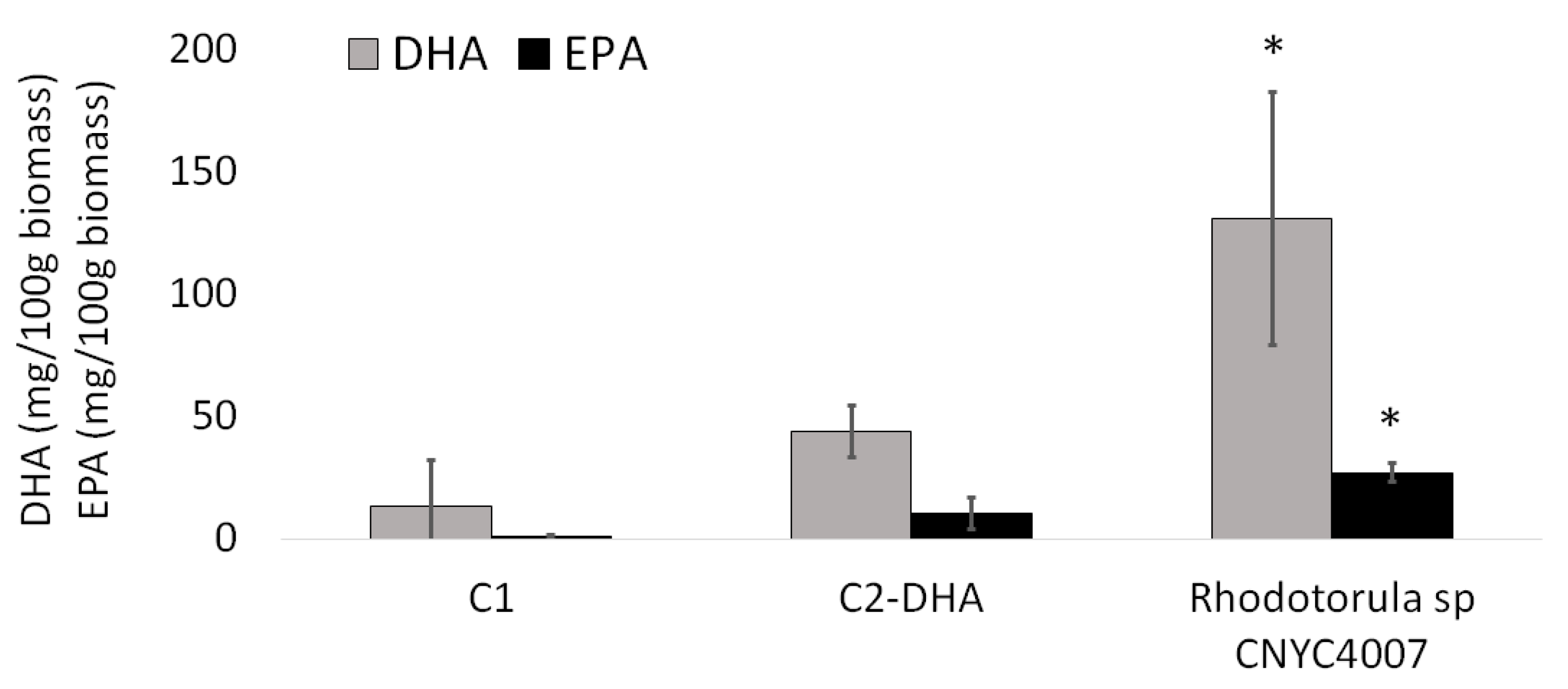
2.5. DHA and EPA Enrichment of Artemia sp. nauplii
3. Discussion
4. Materials and Methods
4.1. Obtaining and Producing the Strain Rhodotorula sp. CNYC4007
4.2. Obtaining and Maintaining Danio rerio Larvae
4.3. Validation of the RNA/DNA Method for Danio rerio Larvae
4.4. Estimating Larval Growth
4.5. Experiments with First-Feeding Age Larvae
4.5.1. Feeding with Rhodotorula sp. CNYC4007 Lyophilized in Flour and in Microcapsules
4.5.2. Larval Feeding with Bioencapsulation in Brachionus sp.
4.6. Experiments with Larvae at 12 Days Post-Yolk Absorption
4.7. Obtaining the RNA/DNA Ratio Used as a Proxy for the Condition of Danio rerio Larvae
4.8. Analysis of Fatty Acid Content in Artemia sp. nauplii
4.9. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Food and Agriculture Organization (FAO). El Estado Mundial de la Pesca y Acuicultura; Departamento de Pesca y Acuicultura: Roma, Italy, 2004; pp. 1–115. [Google Scholar]
- Food and Agriculture Organization (FAO). El Estado Mundial de la Pesca y Acuicultura; Departamento de Pesca y Acuicultura: Roma, Italy, 2012; pp. 1–251. [Google Scholar]
- Tocher, D.R. Fatty acid requirement in ontogeny of marine and freshwater fish. Aquac. Res. 2010, 41, 717–732. [Google Scholar] [CrossRef]
- Food and Agriculture Organization (FAO). El Estado Mundial de la Pesca y Acuicultura; Departamento de Pesca y Acuicultura: Roma, Italy, 2008; pp. 1–280. [Google Scholar]
- Hardy, R.W.; Higgs, D.A.; Lalla, S.P.; Tacon, A.G.J. Alternative dietary protein and lipid sources for sustainable production of salmonids. Fisken og Havet 2001, 8, 44. [Google Scholar]
- Izquierdo, M.S.; Turkmen, S.; Montero, D.; Zamorano, M.J.; Afonso, J.M.; Karalazos, V.; Fernández-Palacios, H. Nutritional programming through broodstock diets to improve utilization of very low fishmeal and fish oil diets in gilthead sea bream. Aquaculture 2015, 449, 18–26. [Google Scholar] [CrossRef]
- Izquierdo, M.; Socorro, J.; Arantzamendi, L.; Hernandez-Cruz, C. Recent advances in lipid nutrition in fish larvae. Fish Physiol. Biochem. 2000, 22, 97–107. [Google Scholar] [CrossRef]
- Hardy, R.W. Utilization on plant proteins in fish diets: Effects of global demand and supplies of fishmeal. Aquac. Res. 2010, 41, 770–776. [Google Scholar] [CrossRef]
- Arslan, M.; Sirkecioglu, N.; Bayir, A.; Arslan, H.; Aras, M. The Influence of substitution of dietary fish oil with different vegetable oils on performance and fatty acid composition of brown trout, Salmo trutta. Turk. J. Fish. Aquat. Sci. 2012, 12, 575–583. [Google Scholar] [CrossRef]
- Díaz, N. Efecto de la Relación EPA/DHA en Larvas de Puye (Galaxias maculatus, Jenyns. 1842), Cultivadas en Diferentes Salinidades. Tesis de Grado, Facultad de Acuicultura y Ciencias Veterinarias, Universidad Católica de Temuco, Temuco, Chile, 2004; p. 70. [Google Scholar]
- Kaushik, S.; Georga, I.; Koumoundouros, G. Growth and body composition of zebrafish (Danio rerio) larvae fed a compound feed from first feeding onward: Toward implications on nutrients requirements. Zebrafish 2011, 8, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Li, K.; Ostensen, M.; Attramadal, K.; Winge, P.; Sparstad, T.; Bones, A.; Vadstein, O.; Kjørsvik, E.; Olsen, Y. Gene regulation of lipid and phospholipid metabolism in Atlantic cod (Gadus morhua) larvae. Comp. Biochem. Physiol. Part B 2015, 190, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Hamre, K.; Harboe, T. Artemia enriched with high n-3 HUFA may give a large improvement in performance of Atlantic halibut (Hippoglossus hippoglossus L.) larvae. Aquaculture 2008, 277, 239–243. [Google Scholar] [CrossRef]
- Estudillo del Castillo, C.; Gapasin, R.; Leaño, E. Enrichment potential of HUFA-rich thraustochytrid Schizochytrium mangrovei for the rotifer Brachionus plicatilis. Aquaculture 2009, 293, 57–61. [Google Scholar] [CrossRef]
- Satuino, C.; Hirayama, K. Fat-soluble vitamin requirements of the rotifer Brachionus plicatilis. In The First Asian Fisheries Forum; Maclean, L., Dizon, L.B., Hosillos, L.V., Eds.; Asian Fisheries Society: Manila, Philippines, 1986; pp. 619–622. [Google Scholar]
- Hawkyard, M.; Sæle, Ø.; Nordgreen, A.; Langdon, C.; Hamre, K. Effect of iodine enrichment of Artemia sp on their nutritional value for larval zebrafish (Danio rerio). Aquaculture 2001, 316, 37–43. [Google Scholar] [CrossRef]
- Ben Khemis, I.; de la Noue, J.; Audet, C. Feeding larvae of winter flounder Pseudopleuronectes americanus (Walbaum) with live prey or microencapsulated diet: Linear growth and protein, RNA and DNA content. Aquac. Res. 2000, 31, 377–386. [Google Scholar] [CrossRef]
- Robinson, S.; Ware, D. Ontogenetic development of growth rates in larval Pacific herring, Clupea harengus pallasi, measured with RNA/DNA ratios in the Strait of Georgia, British Columbia. Can. J. Fish. Aquat. Sci. 1988, 45, 1422–1429. [Google Scholar] [CrossRef]
- Chícharo, M.; Chícharo, L. RNA: DNA Ratio and Other Nucleic Acid Derived Indices in Marine Ecology. Int. J. Mol. Sci. 2008, 9, 1453–1471. [Google Scholar] [CrossRef] [PubMed]
- Wing, N.; Campbell, P.; Dick, J.; Bell, J. Interactive effect of dietary palm oil concentration and water temperature on lipid digestibility in rainbow trout, Oncorhynchus mykiss. Lipids 2003, 38, 1031–1038. [Google Scholar]
- Turchini, G.; Torstensen, B.; Ng, W. Fish oil replacement in finfish nutrition. Rev. Aquac. 2009, 1, 10–57. [Google Scholar] [CrossRef]
- Valenzuela, A.; Sanhueza, J.; De la Barra, F. El aceite de pescado: Un desecho industrial transformado en un producto de alto valor comercial. Grasas Aceites 2011, XXII, 84–98. [Google Scholar]
- Morais, S.; Pratoomyot, J.; Taggart, J.; Bron, J.; Guy, D.; Gordon, J.; Tocher, D. Genotype-specific responses in Atlantic salmon (Salmo salar) subject to dietary fish oil replacement by vegetable oil: A liver transcriptomic analysis. BMC. Genom. 2011, 12, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Kiron, V. Fish immune system and its nutritional modulation for preventive health care. Anim. Feed Sci. 2012, 173, 111–133. [Google Scholar] [CrossRef]
- Miller, M.; Nichols, P.; Carter, C. Replacement of fish oil with thraustochytrid Schizochytrium sp L oil in Atlantic salmon parr (Salmo salar L) diets. Comp. Biochem. Physiol. Part A 2007, 148, 382–392. [Google Scholar] [CrossRef] [PubMed]
- Armenta, R.E.; Valentine, M.C. Single-Cell Oils as a Source of Omega-3 Fatty Acids: An Overview of Recent Advances. J. Am. Oil Chem. Soc. 2013, 90, 167–182. [Google Scholar] [CrossRef]
- Pino, N.L.; Socias, C.; González, R.R. Marine fungoid producers of DHA, EPA and carotenoids from central and southern Chilean marine ecosystems. Rev. Biol. Mar. Oceanogr. 2015, 50, 507–520. [Google Scholar] [CrossRef]
- Iida, I.; Nakahara, T.; Yocochi, T.; Kamisaka, Y.; Yagi, H.; Yamaoka, M.; Suzuki, O. Improvement of docosahexaenoic acid production in a culture of Thraustochytrium aureum by medium optimization. J. Ferment. Bioeng. 1996, 81, 76–78. [Google Scholar] [CrossRef]
- Akanbi, T.O.; Barrow, C.J. Candida antarctica lipase A effectively concentrates DHA from fish and thraustochytrid oils. Food Chem. 2017, 229, 509–516. [Google Scholar] [CrossRef] [PubMed]
- Fan, K.W.; Chen, F.; Jones, E.B.G.; Vrijmoed, L.L.P. Eicosapentaenoic and docosahexaenoic acids production by and okara-utilizing potential of thraustochytrids. J. Ind. Microbiol. Biotechnol. 2001, 27, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Ganuza, E.; Benitez-Santana, T.; Atalah, E.; Vega-Orellana, O.; Ganga, R.; Izquierdo, M. Crypthecodinium cohnii and Schizochytrium sp as potential substitutes to fisheries-derived oils from seabream (Sparus aurata) microdiets. Aquaculture 2008, 277, 109–116. [Google Scholar] [CrossRef]
- Alestrom, P.; Holter, J.; Nourizadeh-Lillabadi, R. Zebrafish in functional genomics and aquatic biomedicine. Trends Biotechnol. 2006, 24, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Ulloa, P.; Iturra, P.; Neira, R.; Araneda, C. Zebrafish as a model organism for nutrition and growth: Towards comparative studies of nutritional genomics applied to aquacultured fishes. Rev. Fish Biol. Fish. 2011, 21, 649–666. [Google Scholar] [CrossRef]
- Ribas, L.; Piferrer, F. The zebrafish (Danio rerio) as a model organism, with emphasis on applications for finfish aquaculture research. Rev. Aquac. 2014, 6, 209–240. [Google Scholar] [CrossRef]
- Ulloa, P.E.; Medrano, J.F.; Feijoo, C.G. Zebrafish as animal model for aquaculture nutrition research. Front. Genet. 2014, 5, 313. [Google Scholar] [CrossRef] [PubMed]
- Hedrera, M.I.; Galdames, J.A.; Jimenez-Reyes, M.F.; Reyes, A.E.; Avendaño-Herrera, R.; Romero, J.; Feijóo, C.G. Soybean Meal Induces Intestinal Inflammation in Zebrafish Larvae. PLoS ONE 2013, 8, e69983. [Google Scholar] [CrossRef] [PubMed]
- Fulton, T. Rate of Growth of Sea-Fishes; Neill & Co.: Edinburgh, UK, 1902; Volume 1, p. 20. [Google Scholar]
- Ikeda, T.; San, F.; Yamaguchi, A.; Matsuishi, T. RNA: DNA ratios of calanoid copepods from the epipelagic through abyssopelagic zones of the North Pacific Ocean. Aquat. Biol. 2007, 1, 99–108. [Google Scholar] [CrossRef]
- Bulow, J.F. RNA-DNA ratios as indicators of growth rates in fish: A review. In The Age and Growth of Fish; Summerfelt, R.C., Hall, G.E., Eds.; The Iowa State University Press: Ames, IA, USA, 1987; pp. 45–64. [Google Scholar]
- Caldarone, E.; Onge-Burns, J.; Buckley, L. Relationship of RNA/DNA ratio and temperature to growth in larvae of Atlantic cod Gadus morhua. Mar. Ecol. Prog. Ser. 2003, 262, 229–240. [Google Scholar] [CrossRef]
- Buckley, L. RNA-DNA ratio: An index of larval fish growth in the sea. Mar. Biol. 1984, 80, 291–298. [Google Scholar] [CrossRef]
- Olivar, M.; Díaz, M.; Chícharo, A. Tissue effect on RNA: DNA ratios of marine fish larvae. Sci. Mar. 2009, 73, 171–182. [Google Scholar] [CrossRef] [Green Version]
- Chung, K.; Segnini, M. RNA-DNA ratio as physiological condition of rainbow trout fry fasted and fed. Ital. J. Zool. 1998, 65, 517–519. [Google Scholar] [CrossRef]
- Narayan, S.; Archana, L.; Ranjan, R. Effects of dietary protein concentrations on growth and RNA: DNA ratio of rainbow trout (Oncorhynchus mykiss) cultured in Nuwakot district of Nepal. Int. J. Fish. Aquat. Stud. 2014, 1, 184–188. [Google Scholar]
- Moreau, D.; Rosenberg, M. Microstructure and fat extractability in microcapsules based on whey proteins or mixtures of whey proteins and lactose. Food Struct. 1993, 12, 457–468. [Google Scholar]
- Habi Mat Dian, N.; Sudin, N.; Affandi, Y. Characteristics of microencapsulated palm-based oil as affected by type of wall material. J. Sci. Food Agric. 1996, 70, 422–426. [Google Scholar] [CrossRef]
- Navarro, J.; Henderson, J.; McEvoy, L.; Bell, M.; Amat, F. Lipid conversions during enrichment of Artemia. Aquaculture 1999, 174, 155–166. [Google Scholar] [CrossRef]
- Silva, A. Cultivo de Peces Marinos; Departamento de Acuicultura, Facultad de Ciencias del Mar, Universidad Católica del Norte: Coquimbo, Chile, 2005. [Google Scholar]
- Barclay, W.; Zeller, S. Nutritional enhancement of n-3 and n-6 fatty acids in rotifers and Artemia nauplii by feeding spray-dried Schizochytrium sp. J. World Aquac. Soc. 1996, 27, 314–322. [Google Scholar] [CrossRef]
- Johansen, K.A.; Overturf, K. Alterations in expression of genes associated with muscle metabolism and growth during nutritional restriction and refeeding in rainbow trout. Comp. Biochem. Physiol. Part B 2006, 144, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, M.; Arakawa, T.; Takeuchi, R.; Haroun, R.; Watanabe, T. Effect of ω-3 HUFA levels in Artemia on growth of larval Japanese flounder (Paralicthys olilaceus). Aquaculture 1992, 105, 73–82. [Google Scholar] [CrossRef]
- Masuda, Y.; Oku, H.; Okumura, T.; Nomura, K.; Kurokawa, T. Feeding restriction alters expression of some ATP related genes more sensitively than the RNA/DNA ratio in zebrafish, Danio rerio. Comp. Biochem. Physiol. Part B 2009, 152, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Salem, M.; Silverstain, J.; Rexroad, C.; Yao, J. Effect of starvation on global gene expression and proteolysis in rainbow trout (Oncorhynchus mykiss). BMC. Genom. 2007, 8, 328. [Google Scholar] [CrossRef] [PubMed]
- Frengova, G.I.; Beshkova, D.M. Carotenoids from Rhodotorula and Phaffia: Yeasts of biotechnological importance. J. Ind. Microbiol. Biotechnol. 2009, 36, 163–180. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, M.C.; Arivalagan, P.; Barre, D.E.; MacInnis, J.A.; D’Cunha, G.B. Rhodotorula glutinis Phenylalanine/Tyrosine Ammonia Lyase Enzyme Catalyzed Synthesis of the Methyl Ester of para-Hydroxycinnamic Acid and its Potential Antibacterial Activity. Front. Microbiol. 2016, 7, 281. [Google Scholar] [CrossRef] [PubMed]
- Taskin, M.; Ortucu, S.; Aydogan, M.N.; Arslan, N.P. Lipid production from sugar beet molasses under non-aseptic culture conditions using the oleaginous yeast Rhodotorula glutinis TR29. Renew. Energy 2016, 99, 198–204. [Google Scholar] [CrossRef]
- Sprague, M.; Betancor, M.B.; Tocher, D.R. Microbial and genetically engineered oils as replacements for fish oil in aquaculture feeds. Biotechnol. Lett. 2017, 39, 1599–1609. [Google Scholar] [CrossRef] [PubMed]
- Pino, N.; Gómez, C.; González, R.R. Proceso de Obtención de Biomasa del Fungoide Marino CNYC 4007, Uso de la Biomasa y Suplemento Alimenticio que Contiene Esta Biomasa; Solicitud CL201402916; Instituto de Propiedad Industrial: Santiago, Chile, 2014.
- Clemmensen, C. RNA/DNA ratios of laboratory reared and wild herring larvae determined with a highly sensitive fluorescence method. J. Fish Biol. 1989, 35, 331–333. [Google Scholar] [CrossRef]
- Li, Z.; Gu, T.; Kelder, B.; Kopchick, J. Analysis of fatty acids in mouse cells using Reversed-Phase High-Performance Liquid. Chromatographia 2001, 54, 463–467. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Larvae Age (Days) | Total Length Larvae (mm) | |||||
|---|---|---|---|---|---|---|
| Brachionus sp. | Artemia sp. nauplii | |||||
| C1 | C2-DHA | CNYC4007 | C1 | C2-DHA | CNYC4007 | |
| 6 | 5.0 ± 0.2 | 5.0 ± 0.2 | 5.0 ± 0.2 | n/a | n/a | n/a |
| 8 | 5.2 ± 0.3 | 5.5 ± 0.4 | 5.1 ± 0.2 | n/a | n/a | n/a |
| 10 | 5.6 ± 0.1 | 5.6 ± 0.3 | 5.6 ± 0.3 | n/a | n/a | n/a |
| 12 | 5.6 ± 0.1 | 5.7 ± 0.2 | 5.8 ± 0.2 | 5.6 ± 0.3 | 5.6 ± 0.3 | 5.6 ± 0.3 |
| 14 | n/a | n/a | n/a | 5.6 ± 0.3 | 5.7 ± 0.3 | 5.8 ± 0.4 |
| 16 | n/a | n/a | n/a | 5.6 ± 0.2 | 6.0 ± 0.3 | 6.0 ± 0.4 |
| 18 | n/a | n/a | n/a | 6.6 ± 0.1 | 6.9 ± 0.3 | 7.0 ± 0.3 |
| 20 | n/a | n/a | n/a | 9.5 ± 0.4 | 10.2 ± 0.3 | 10.1 ± 0.7 |
| 22 | n/a | n/a | n/a | 9.9 ± 0.8 | 10.6 ± 0.3 | 10.6 ± 0.5 |
| Larvae Age (Days) | RNA/DNA Ratio | |||||
|---|---|---|---|---|---|---|
| Brachionus sp. | Artemia sp. nauplii | |||||
| C1 | C2-DHA | CNYC4007 | C1 | C2-DHA | CNYC4007 | |
| 6 | 1.1 ± 0.2 | 1.1 ± 0.2 | 1.1 ± 0.2 | n/a | n/a | n/a |
| 8 | 1.3 ± 0.2 | 1.1 ± 0.3 | 1.4 ± 0.3 | n/a | n/a | n/a |
| 10 | 1.2 ± 0.2 | 1.5 ± 0.1 | 1.5 ± 0.3 | n/a | n/a | n/a |
| 12 | 1.0 ± 0.2 | 1.1 ± 0.1 | 1.4 ± 0.2 * | 0.8 ± 0.2 | 0.8 ± 0.2 | 0.8 ± 0.2 |
| 14 | n/a | n/a | n/a | 1.1 ± 0.1 | 0.9 ± 0.2 | 1.1 ± 0.3 |
| 16 | n/a | n/a | n/a | 1.0 ± 0.1 | 1.1 ± 0.2 | 1.1 ± 0.3 |
| 18 | n/a | n/a | n/a | 1.0 ± 0.2 | 1.1 ± 0.1 | 1.2 ± 0.2 |
| 20 | n/a | n/a | n/a | 0.8 ± 0.1 | 1.1 ± 0.1 | 1.1 ± 0.1 |
| 22 | n/a | n/a | n/a | 0.8 ± 0.1 * | 1.1 ± 0.1 | 1.0 ± 0.1 |
| Components (%) | C1 | C2-DHA | CNYC4007 Microencapsulated | CNYC4007 Meal |
|---|---|---|---|---|
| Protein | 49 | 17.6 | n/a | n/a |
| Carbohydrate | n/a | 15.9 | 92.5 | 36.0 |
| Fat | 8.5 | 56.2 | 0.3 | 7.6 |
| Nitrogen compounds | n/a | n/a | 1.4 | 6.1 |
| Fiber | 3 | n/a | 1.2 | 1.7 |
| Moisture | 6 | 2.1 | 3.7 | 83.1 |
| Ash | n/a | 8.2 | 0.5 | 7.6 |
| Calories (kcal) | n/a | 640 | 378.2 | 169.8 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barra, M.; Llanos-Rivera, A.; Cruzat, F.; Pino-Maureira, N.; González-Saldía, R.R. The Marine Fungi Rhodotorula sp. (Strain CNYC4007) as a Potential Feed Source for Fish Larvae Nutrition. Mar. Drugs 2017, 15, 369. https://doi.org/10.3390/md15120369
Barra M, Llanos-Rivera A, Cruzat F, Pino-Maureira N, González-Saldía RR. The Marine Fungi Rhodotorula sp. (Strain CNYC4007) as a Potential Feed Source for Fish Larvae Nutrition. Marine Drugs. 2017; 15(12):369. https://doi.org/10.3390/md15120369
Chicago/Turabian StyleBarra, M., A. Llanos-Rivera, F. Cruzat, N. Pino-Maureira, and R. R. González-Saldía. 2017. "The Marine Fungi Rhodotorula sp. (Strain CNYC4007) as a Potential Feed Source for Fish Larvae Nutrition" Marine Drugs 15, no. 12: 369. https://doi.org/10.3390/md15120369
APA StyleBarra, M., Llanos-Rivera, A., Cruzat, F., Pino-Maureira, N., & González-Saldía, R. R. (2017). The Marine Fungi Rhodotorula sp. (Strain CNYC4007) as a Potential Feed Source for Fish Larvae Nutrition. Marine Drugs, 15(12), 369. https://doi.org/10.3390/md15120369