Effect of Marine-Derived Ice-Binding Proteins on the Cryopreservation of Marine Microalgae


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
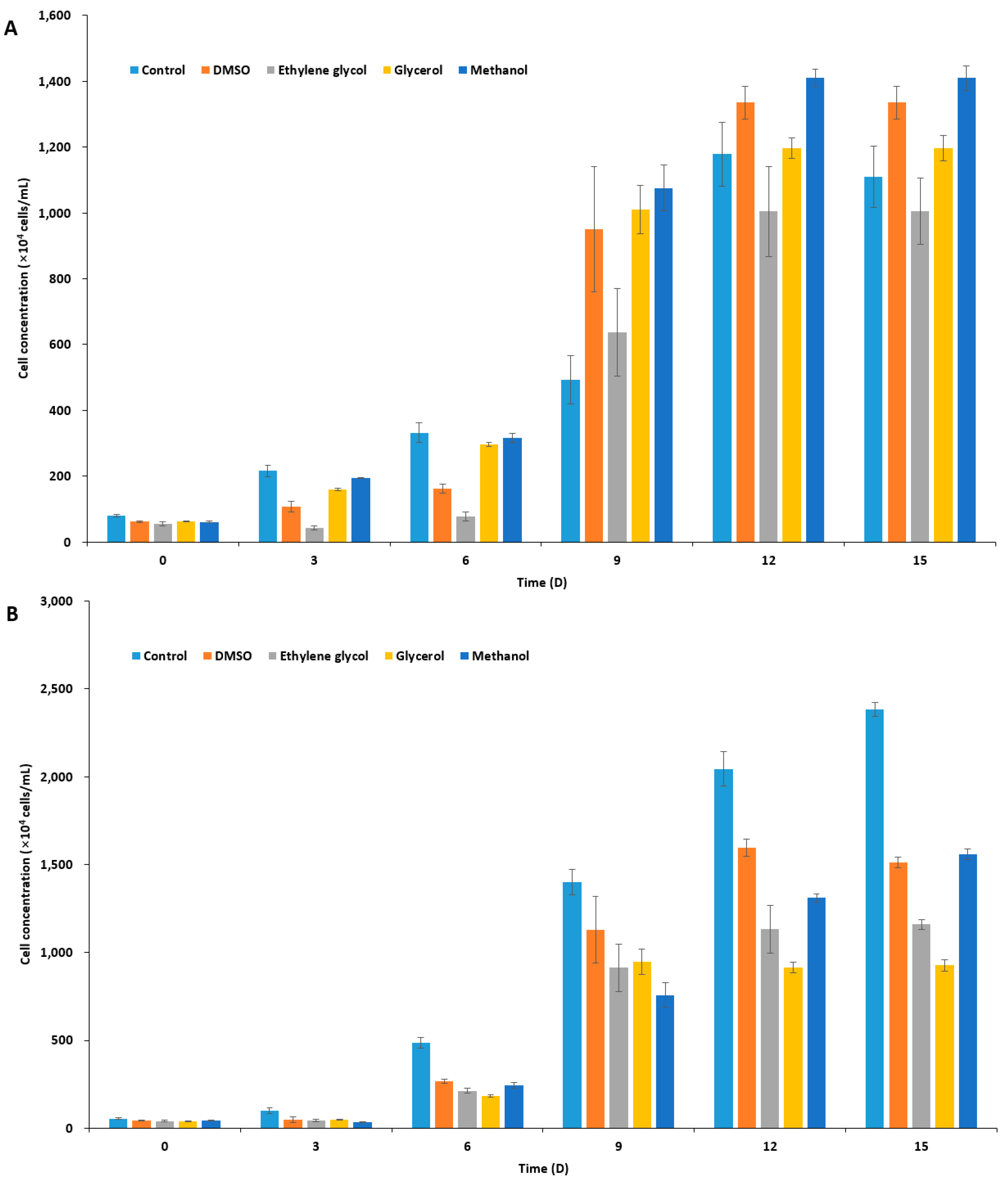
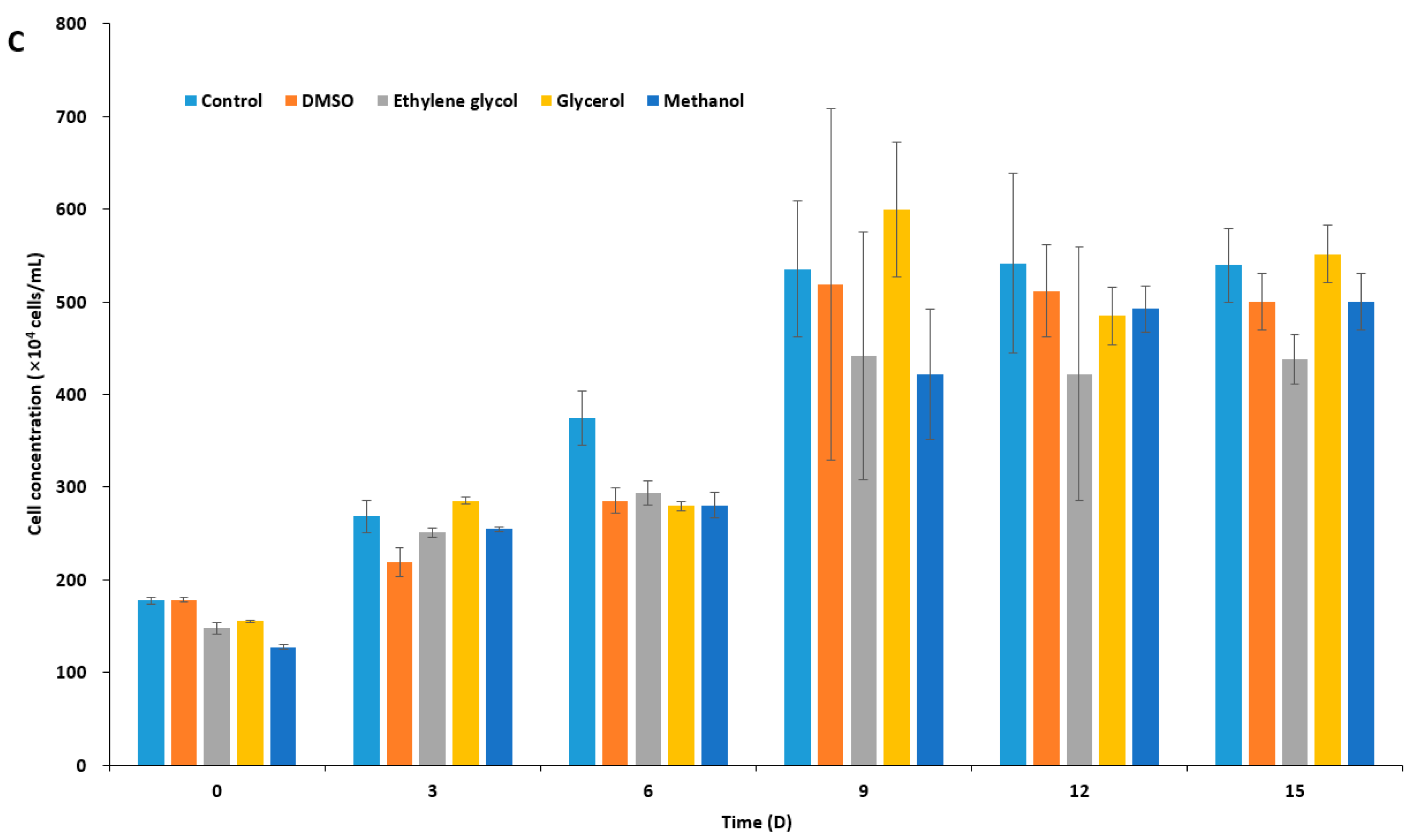
2.1. Effect of CPAs on Unfrozen Marine Microalgae
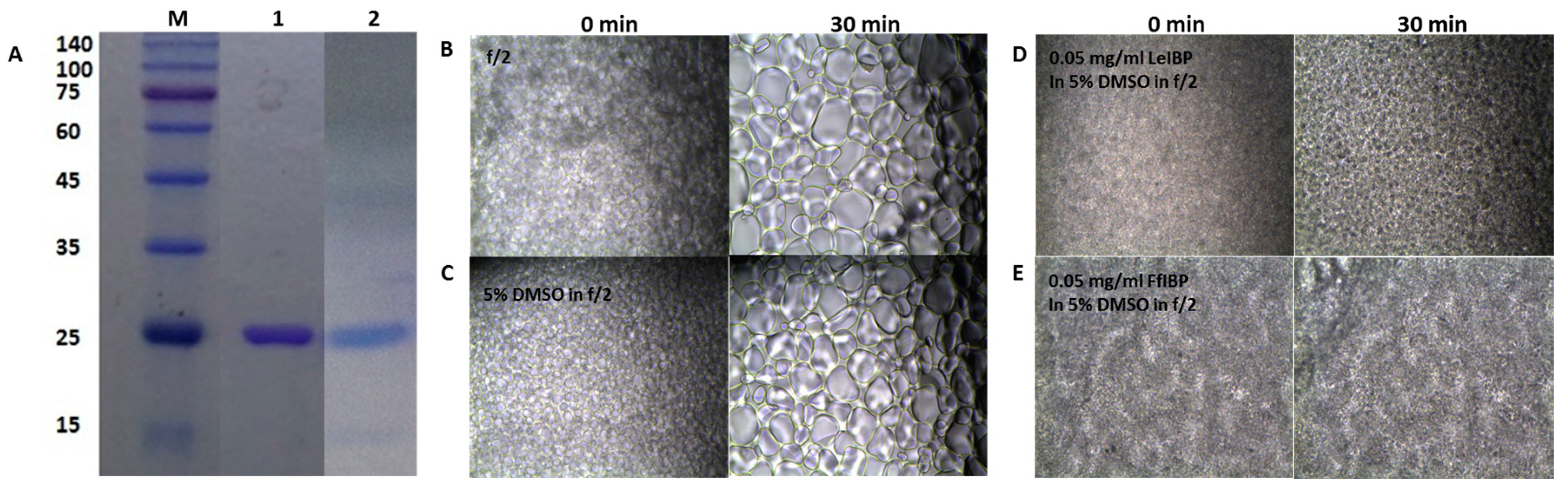
2.2. Ice Recrystallization Inhibition of Marine IBPs in Freezing Media
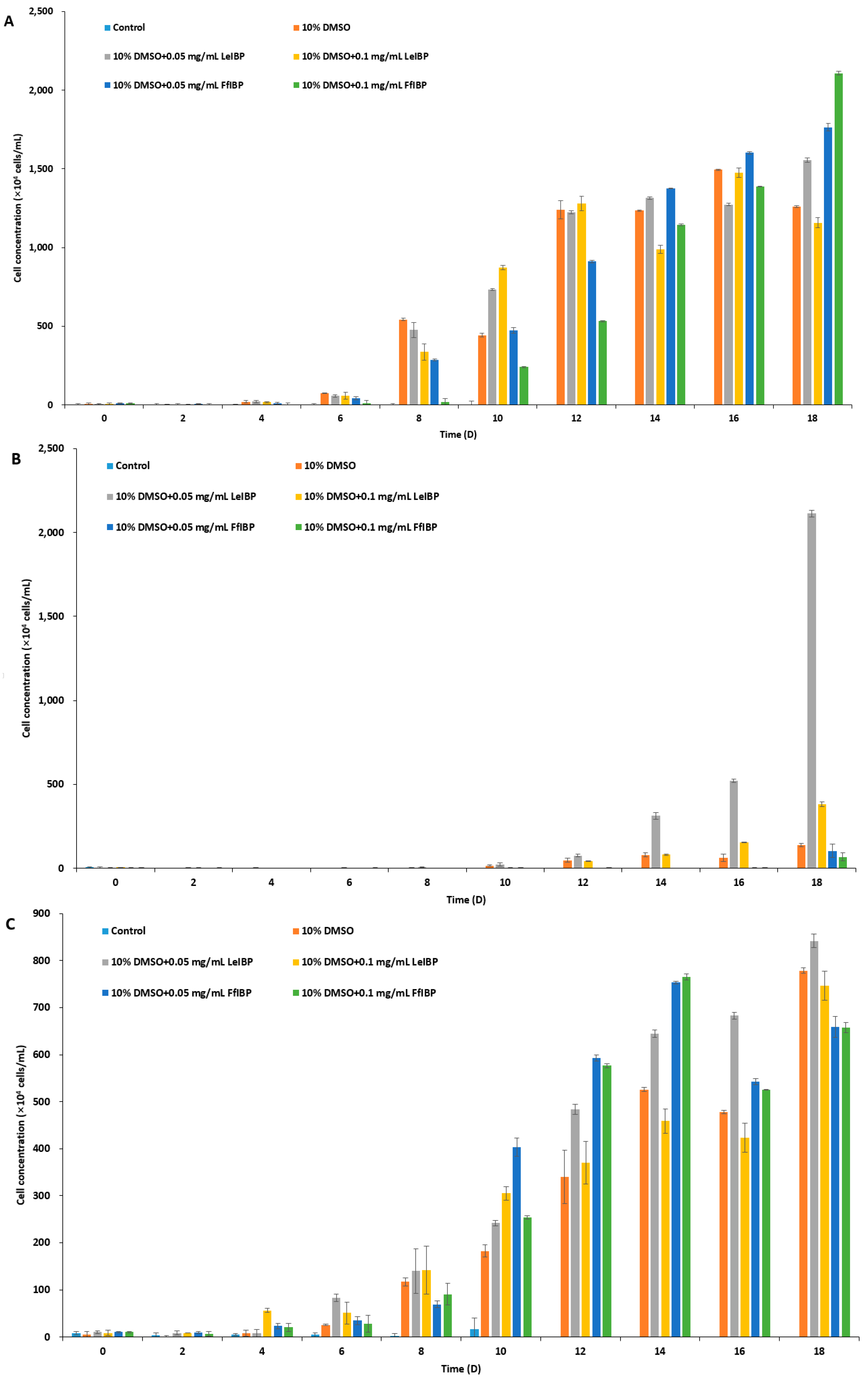
2.3. Viability of Cryopreserved Microalgae
3. Materials and Methods
3.1. Chemicals and Ice-Binding Proteins
3.2. Microalgae Strains and Culture Conditions
3.3. Ice Recrystallization Inhibition (IRI) Assay
3.4. Toxicity of Four Penetrating CPAs
3.5. Cryopreservation and Viability of Marine Microalgae
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kim, H.J.; Lee, J.H.; Hur, Y.B.; Lee, C.W.; Park, S.-H.; Koo, B.-W. Marine Antifreeze Proteins: Structure, Function, and Application to Cryopreservation as a Potential Cryoprotectant. Mar. Drugs 2017, 15, 27. [Google Scholar] [CrossRef] [PubMed]
- Davies, P.L. Ice-binding proteins: A remarkable diversity of structures for stopping and starting ice growth. Trends Biochem. Sci. 2014, 39, 548–555. [Google Scholar] [CrossRef] [PubMed]
- Kristiansen, E.; Zachariassen, K.E. The mechanism by which fish antifreeze proteins cause thermal hysteresis. Cryobiology 2005, 51, 262–280. [Google Scholar] [CrossRef] [PubMed]
- Raymond, J.A.; Fritsen, C.; Shen, K. An ice-binding protein from an Antarctic sea ice bacterium. FEMS Microbiol. Ecol. 2007, 61, 214–221. [Google Scholar] [CrossRef] [PubMed]
- Raymond, J.A. Algal ice-binding proteins change the structure of sea ice. Proc. Natl. Acad. Sci. USA 2011, 108, E198. [Google Scholar] [CrossRef] [PubMed]
- Raymond, J.A.; Janech, M.G. Cryoprotective property of diatom ice-active substance. Cryobiology 2003, 46, 203–204. [Google Scholar] [CrossRef]
- Raymond, J.A.; Knight, C.A. Ice binding, recrystallization inhibition, and cryoprotective properties of ice-active substances associated with Antarctic sea ice diatoms. Cryobiology 2003, 46, 174–181. [Google Scholar] [CrossRef]
- Bayer-Giraldi, M.; Weikusat, I.; Besir, H.; Dieckmann, G. Characterization of an antifreeze protein from the polar diatom Fragilariopsis cylindrus and its relevance in sea ice. Cryobiology 2011, 63, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Garnham, C.P.; Whitney, J.C.; Graham, L.A.; Davies, P.L. Re-evaluation of a bacterial antifreeze protein as an adhesin with ice-binding activity. PLoS ONE 2012, 7, e48805. [Google Scholar] [CrossRef] [PubMed]
- Mangiagalli, M.; Bar-Dolev, M.; Tedesco, P.; Natalello, A.; Kaleda, A.; Brocca, S.; Pascale, D.; Pucciarelli, S.; Miceli, C.; Bravslavsky, I. Cryo-protective effect of an ice-binding protein derived from Antarctic bacteria. FEBS J. 2016, 284, 163–177. [Google Scholar] [CrossRef] [PubMed]
- Knight, C.A.; DeVries, A.L.; Oolman, L.D. Fish antifreeze protein and the freezing and recrystallization of ice. Nature 1984, 308, 295–296. [Google Scholar] [CrossRef] [PubMed]
- Knight, C.A.; Duman, J.G. Inhibition of recrystallization of ice by insect thermal hysteresis proteins: A possible cryoprotective role. Cryobiology 1986, 23, 256–262. [Google Scholar] [CrossRef]
- Raymond, J.A.; Christner, B.C.; Schuster, S.C. A bacterial ice-binding protein from the Vostok ice core. Extremophiles 2008, 12, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.S.; Raymond, J.A. Reduction of freeze-thaw-induced hemolysis of red blood cells by an algal ice-binding protein. Cryoletters 2004, 25, 307–310. [Google Scholar] [PubMed]
- Lee, S.G.; Koh, H.Y.; Lee, J.H.; Kang, S.H.; Kim, H.J. Cryopreservative effects of the recombinant ice-binding protein from the arctic yeast leucosporidium sp. on red blood cells. Appl. Biochem. Biotechnol. 2012, 167, 824–834. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.K.; Park, K.S.; Park, S.; Park, H.; Song, Y.H.; Kang, S.H.; Kim, H.J. An extracellular ice-binding glycoprotein from an Arctic psychrophilic yeast. Cryobiology 2010, 60, 222–228. [Google Scholar] [CrossRef] [PubMed]
- Park, K.S.; Do, H.; Lee, J.H.; Park, S.I.; Kim, E.J.; Kim, S.J.; Kang, S.H.; Kim, H.J. Characterization of the ice-binding protein from Arctic yeast Leucosporidium sp. AY30. Cryobiology 2012, 64, 286–296. [Google Scholar] [CrossRef] [PubMed]
- Do, H.; Lee, J.H.; Lee, S.G.; Kim, H.J. Crystallization and preliminary X-ray crystallographic analysis of an ice-binding protein (FfIBP) from Flavobacterium frigoris PS1. Acta Crystallogr. Sect. F Struct. Biol. Cryst. Commun. 2012, 68, 806–809. [Google Scholar] [CrossRef] [PubMed]
- Do, H.; Kim, S.J.; Kim, H.J.; Lee, J.H. Structure-based characterization and antifreeze properties of a hyperactive ice-binding protein from the Antarctic bacterium Flavobacterium frigoris PS1. Acta Crystallogr. D Biol. Crystallogr. 2014, 70, 1061–1073. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Park, A.K.; Do, H.; Park, K.S.; Moh, S.H.; Chi, Y.M.; Kim, H.J. Structural basis for the antifreeze activity of an ice-binding protein from an Arctic yeast. J. Biol. Chem. 2012. [Google Scholar] [CrossRef] [PubMed]
- Olijve, L.L.C.; Oude Vrielink, A.S.; Voets, I.K. A simple and quantitative method to evaluate ice recrystallization kinetics using the circle Hough Transform algorithm. Cryst. Growth Des. 2016, 16, 4190–4195. [Google Scholar] [CrossRef]
- Yu, S.O.; Brown, A.; Middleton, A.J.; Tomczak, M.M.; Walker, V.K.; Davies, P.L. Ice restructuring inhibition activities in antifreeze proteins with distinct differences in thermal hysteresis. Cryobiology 2010, 61, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, P.A.; Márquez, S.L.; González-Nilo, F.D.; Márquez-Miranda, V.; Blamey, J.M. Structure and application of antifreeze proteins from Antarctic bacteria. Microb. Cell Fact. 2017, 16, 138. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Shim, H.E.; Lee, J.H.; Kang, Y.-C.; Hur, Y.B. Ice-Binding Protein Derived from Glaciozyma Can Improve the Viability of Cryopreserved Mammalian Cells. J. Microbiol. Biotechnol. 2015, 25, 1989–1996. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.H.; Lee, H.J.; Kim, H.J.; Lee, J.H.; Ko, Y.; Kim, S.M.; Lee, J.R.; Suh, C.S.; Kim, S.H. Effects of Antifreeze Proteins on the Vitrification of Mouse Oocytes: Comparison of Three Different Antifreeze Proteins. Hum. Reprod. 2015, 30, 2110–2119. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kim, S.K.; Youm, H.W.; Kim, H.J.; Lee, J.R.; Suh, C.S.; Kim, S.H. Effects of Three Different Types of Antifreeze Proteins on Mouse Ovarian Tissue Cryopreservation and Transplantation. PLoS ONE 2015, 10, e0126252. [Google Scholar] [CrossRef] [PubMed]
- Koh, H.Y.; Lee, J.H.; Han, S.J.; Park, H.; Lee, S.G. Effect of the Antifreeze Protein from the Arctic Yeast Leucosporidium sp. AY30 on Cryopreservation of the Marine Diatom Phaeodactylum tricornutum. Appl. Biochem. Biotechnol. 2015, 175, 677–686. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Yoon, J.H.; Park, G.H.; Bae, S.H.; Kim, H.J.; Kim, M.S.; Hwang, Y.J.; Kim, D.Y. Influence of antifreeze proteins on boar sperm DNA damaging during cryopreservation. Dev. Biol. 2011, 356, 195. [Google Scholar] [CrossRef]
- Day, J.G.; Stacey, G. Cryopreservation and Freeze-Drying Protocols; Springer Science & Business Media: New York, NY, USA, 2007; Volume 368, ISBN 1588293777. [Google Scholar]
- Liu, L.; Pohnert, G.; Wei, D. Extracellular metabolites from industrial microalgae and their biotechnological potential. Mar. Drugs 2016, 14, 191. [Google Scholar] [CrossRef] [PubMed]
- Tseng, C.K.; Chen, J.; Zhang, Z. On a new species of Pavlova (Prymnesiophyceae) from China. Chin. J. Oceanol. Limnol. 1992, 10, 22–38. [Google Scholar] [CrossRef]
- Walter, C.P.; Posten, C. (Eds.) Microalgal Biotechnology: Potential and Production; De Gruyter: Berlin, Germany, 2012. [Google Scholar]
- Tzovenis, I.; De Pauw, N.; Sorgeloos, P. Effect of different light regimes on the docosahexaenoic acid (DHA) content of Isochrysis aff. galbana (clone T-ISO). Aquac. Int. 1997, 5, 489–507. [Google Scholar] [CrossRef]
- Molina Grima, E.; Sánchez Pérez, J.A.; García Camacho, F.; Acién Fernández, F.G.; López Alonso, D.; Segura del Castillo, C.I. Preservation of the marine microalga, Isochrysis galbana: Influence on the fatty acid profile. Aquaculture 1994, 123, 377–385. [Google Scholar] [CrossRef]
- Haas, S.; Bauer, J.L.; Adakli, A.; Meyer, S.; Lippemeier, S.; Schwarz, K.; Schulz, C. Marine microalgae Pavlova viridis and Nannochloropsis sp. as n-3 PUFA source in diets for juvenile European sea bass (Dicentrarchus labrax L.). J. Appl. Phycol. 2016, 28, 1011–1021. [Google Scholar] [CrossRef]
- Aldana-Aranda, D.; Suarez, V.P. Overview of diets used in larviculture of three Caribbean Conchs: Queen Conch Strombus gigas, Milk Conch Strombus costatus and Fighting Conch Strombus pugilis. Aquaculture 1998, 167, 163–178. [Google Scholar] [CrossRef]
- Bendif, E.M.; Probert, I.; Schroeder, D.C.; de Vargas, C. On the description of Tisochrysis lutea gen. nov. sp. nov. and Isochrysis nuda sp. nov. in the Isochrysidales, and the transfer of Dicrateria to the Prymnesiales (Haptophyta). J. Appl. Phycol. 2013, 25, 1763–1776. [Google Scholar] [CrossRef]
- Heasman, M.; Diemar, J.; O’connor, W.; Sushames, T.; Foulkes, L. Development of extended shelf-life microalgae concentrate diets harvested by centrifugation for bivalve molluscs—A summary. Aquac. Res. 2000, 31, 637–659. [Google Scholar] [CrossRef]
- Zhu, C.J.; Lee, Y.K.; Chao, T.M. Effects of temperature and growth phase on lipid and biochemical composition of Isochrysis galbana TK1. J. Appl. Phycol. 1997, 9, 451–457. [Google Scholar] [CrossRef]
- Liu, C.-P.; Lin, L.-P. Ultrastructural study and lipid formation of Isochrysis sp. CCMP1324. Bot. Bull. Acad. Sin. 2001, 42, 207–214. [Google Scholar]
- Barakoni, R.; Awal, S.; Christie, A. Growth performance of the marine microalgae Pavlova salina and Dunaliella tertiolecta using different commercially available fertilizers in natural seawater and inland saline ground water. J. Algal Biomass Utln. 2015, 6, 15–25. [Google Scholar]
- Eikrem, W.; Medlin, L.K.; Henderiks, J.; Rokitta, S.; Rost, B.; Probert, I.; Throndsen, J.; Edvardsen, B. Haptophyta. In Handbook of the Protists; Springer: Cham, Switzerland, 2016; pp. 1–61. [Google Scholar]
- Imam, S.H.; Buchanan, M.J.; Shin, H.-C.; Snell, W.J. The Chlamydomonas cell wall: Characterization of the wall framework. J. Cell Biol. 1985, 101, 1599–1607. [Google Scholar] [CrossRef] [PubMed]
- FAO. FAO Year Book: Fishery and Aquaculture Statistics 2015; FAO: Rome, Italy, 2017. [Google Scholar]
- Tzovenis, I.; Triantaphyllidis, G.; Naihong, X.; Chatzinikolaou, E.; Papadopoulou, K.; Xouri, G.; Tafas, T. Cryopreservation of marine microalgae and potential toxicity of cryoprotectants to the primary steps of the aquacultural food chain. Aquaculture 2004, 230, 457–473. [Google Scholar] [CrossRef]
- Rhodes, L.; Smith, J.; Tervit, R.; Roberts, R.; Adamson, J.; Adams, S.; Decker, M. Cryopreservation of economically valuable marine micro-algae in the classes Bacillariophyceae, Chlorophyceae, Cyanophyceae, Dinophyceae, Haptophyceae, Prasinophyceae, and Rhodophyceae. Cryobiology 2006, 52, 152–156. [Google Scholar] [CrossRef] [PubMed]
- Brand, J.J.; Diller, K.R. Application and theory of algal cryopreservation. Nova Hedwig. 2004, 79, 175–189. [Google Scholar] [CrossRef]
- Abreu, L.; Borges, L.; Marangoni, J.; Abreu, P.C. Cryopreservation of some useful microalgae species for biotechnological exploitation. J. Appl. Phycol. 2012, 24, 1579–1588. [Google Scholar] [CrossRef]
- Salas-Leiva, J.S.; Dupré, E. Cryopreservation of the microalgae Chaetoceros calcitrans (Paulsen): Analysis of the effect of DMSO temperature and light regime during different equilibrium periods. Lat. Am. J. Aquat. Res. 2011, 39, 271–279. [Google Scholar] [CrossRef]
- Poncet, J.M.; Véron, B. Cryopreservation of the unicellular marine alga, Nannochloropsis oculata. Biotechnol. Lett. 2003, 25, 2017–2022. [Google Scholar] [CrossRef] [PubMed]
- Boroda, A.V.; Aizdaicher, N.A.; Odintsova, N.A. The influence of ultra-low temperatures on marine microalgal cells. J. Appl. Phycol. 2014, 26, 387–397. [Google Scholar] [CrossRef]
- Taylor, R.; Fletcher, R.L. Cryopreservation of eukaryotic algae—A review of methodologies. J. Appl. Phycol. 1998, 10, 481–501. [Google Scholar] [CrossRef]
- Wagner, S.; Hoffer, S. Comparative Analysis of Two Cryopreservatives on Two Marine Plankton Species: Isochrysis and Pseudo-nitzschia. UWT J. Environ. 2005, 3, 1–8. [Google Scholar]
- Cañavate, J.P.; Lubian, L.M. Relationship between cooling rates, cryoprotectant concentrations and salinities in the cryopreservation of marine microalgae. Mar. Biol. 1995, 124, 325–334. [Google Scholar] [CrossRef]
- Wang, Q.; Shi, R.; Cheng, A. Cryopreservation of 3 golden algae (Chrysophyta) used as feed in mariculture. J. Fish. Sci. China 1999, 6, 89–92. [Google Scholar]
- Hong, S.S.; Lee, S.Y.; Kim, Y.N.; Kang, S.-H.; Kim, H.J. A Modified Cryopreservation Method of Psychrophilic Chlorophyta Pyramimonas sp. from Antarctica. Ocean Polar Res. 2011, 33, 303–308. [Google Scholar] [CrossRef]
- Christaki, U.; Belviso, S.; Dolan, J.R.; Corn, M. Assessment of the role of copepods and ciliates in the release to solution of particulate DMSP. Mar. Ecol. Prog. Ser. 1996, 114, 119–127. [Google Scholar] [CrossRef]
- Asher, E.C.; Dacey, J.W.H.; Jarniková, T.; Tortell, P.D. Measurement of DMS, DMSO, and DMSP in natural waters by automated sequential chemical analysis. Limnol. Oceanogr. Methods 2015, 13, 451–462. [Google Scholar] [CrossRef]
- Sunda, W.; Kieber, D.J.; Kiene, R.P.; Huntsman, S. An antioxidant function for DMSP and DMS in marine algae. Nature 2002, 418, 317. [Google Scholar] [CrossRef] [PubMed]
- Kirst, G.O.; Thiel, C.; Wolff, H.; Nothnagel, J.; Wanzek, M.; Ulmke, R. Dimethylsulfoniopropionate (DMSP) in icealgae and its possible biological role. Mar. Chem. 1991, 35, 381–388. [Google Scholar] [CrossRef]
- Fowler, A.; Toner, M. Cryo-injury and biopreservation. Ann. N. Y. Acad. Sci. 2006, 1066, 119–135. [Google Scholar] [CrossRef] [PubMed]
- Tam, R.Y.; Rowley, C.N.; Petrov, I.; Zhang, T.; Afagh, N.A.; Woo, T.K.; Ben, R.N. Solution conformation of C-linked antifreeze glycoprotein analogues and modulation of ice recrystallization. J. Am. Chem. Soc. 2009, 131, 15745–15753. [Google Scholar] [CrossRef] [PubMed]
- Chaytor, J.L.; Tokarew, J.M.; Wu, L.K.; Leclère, M.; Tam, R.Y.; Capicciotti, C.J.; Guolla, L.; Von Moos, E.; Findlay, C.S.; Allan, D.S.; et al. Inhibiting ice recrystallization and optimization of cell viability after cryopreservation. Glycobiology 2012, 22, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.K.; Tokarew, J.M.; Chaytor, J.L.; Von Moos, E.; Li, Y.; Palii, C.; Ben, R.N.; Allan, D.S. Carbohydrate-mediated inhibition of ice recrystallization in cryopreserved human umbilical cord blood. Carbohydr. Res. 2011, 346, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Halwani, D.O.; Brockbank, K.G.; Duman, J.G.; Campbell, L.H. Recombinant Dendroides canadensis antifreeze proteins as potential ingredients in cryopreservation solutions. Cryobiology 2014, 68, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Jo, J.W.; Jee, B.C.; Lee, J.R.; Suh, C.S. Effect of antifreeze protein supplementation in vitrification medium on mouse oocyte developmental competence. Fertil. Steril. 2011, 96, 1239–1245. [Google Scholar] [CrossRef] [PubMed]
- Payne, S.R.; Oliver, J.E.; Upreti, G.C. Effect of antifreeze proteins on the motility of ram spermatozoa. Cryobiology 1994, 31, 180–184. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, G.L.; Hew, C.L.; Davies, P.L. Antifreeze proteins of teleost fishes. Annu. Rev. Physiol. 2001, 63, 359–390. [Google Scholar] [CrossRef] [PubMed]
- Jia, Z.; Davies, P.L. Antifreeze proteins: An unusual receptor-ligand interaction. Trends Biochem. Sci. 2002, 27, 101–106. [Google Scholar] [CrossRef]
- Carpenter, J.F.; Hansen, T.N. Antifreeze protein modulates cell survival during cryopreservation: Mediation through influence on ice crystal growth. Proc. Natl. Acad. Sci. USA 1992, 89, 8953–8957. [Google Scholar] [CrossRef] [PubMed]
- Chao, H.; Davies, P.L.; Carpenter, J.F. Effects of antifreeze proteins on red blood cell survival during cryopreservation. J. Exp. Biol. 1996, 199, 2071–2076. [Google Scholar] [PubMed]
- Lee, J.H.; Lee, S.G.; Do, H.; Park, J.C.; Kim, E.; Choe, Y.H.; Han, S.J.; Kim, H.J. Optimization of the pilot-scale production of an ice-binding protein by fed-batch culture of Pichia pastoris. Appl. Microbiol. Biotechnol. 2013, 97, 3383–3393. [Google Scholar] [CrossRef] [PubMed]
- Knight, C.A.; Hallett, J.; DeVries, A.L. Solute effects on ice recrystallization: An assessment technique. Cryobiology 1988, 25, 55–60. [Google Scholar] [CrossRef]
- Jackman, J.; Noestheden, M.; Moffat, D.; Pezacki, J.P.; Findlay, S.; Ben, R.N. Assessing antifreeze activity of AFGP 8 using domain recognition software. Biochem. Biophys. Res. Commun. 2007, 354, 340–344. [Google Scholar] [CrossRef] [PubMed]





© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, H.J.; Koo, B.-W.; Kim, D.; Seo, Y.S.; Nam, Y.K. Effect of Marine-Derived Ice-Binding Proteins on the Cryopreservation of Marine Microalgae. Mar. Drugs 2017, 15, 372. https://doi.org/10.3390/md15120372
Kim HJ, Koo B-W, Kim D, Seo YS, Nam YK. Effect of Marine-Derived Ice-Binding Proteins on the Cryopreservation of Marine Microalgae. Marine Drugs. 2017; 15(12):372. https://doi.org/10.3390/md15120372
Chicago/Turabian StyleKim, Hak Jun, Bon-Won Koo, Doa Kim, Ye Seul Seo, and Yoon Kwon Nam. 2017. "Effect of Marine-Derived Ice-Binding Proteins on the Cryopreservation of Marine Microalgae" Marine Drugs 15, no. 12: 372. https://doi.org/10.3390/md15120372
APA StyleKim, H. J., Koo, B. -W., Kim, D., Seo, Y. S., & Nam, Y. K. (2017). Effect of Marine-Derived Ice-Binding Proteins on the Cryopreservation of Marine Microalgae. Marine Drugs, 15(12), 372. https://doi.org/10.3390/md15120372