
Protective Effects of Fucoidan on Aβ25–35 and d-Gal-Induced Neurotoxicity in PC12 Cells and d-Gal-Induced Cognitive Dysfunction in Mice

Abstract
:1. Introduction
2. Results and Discussion
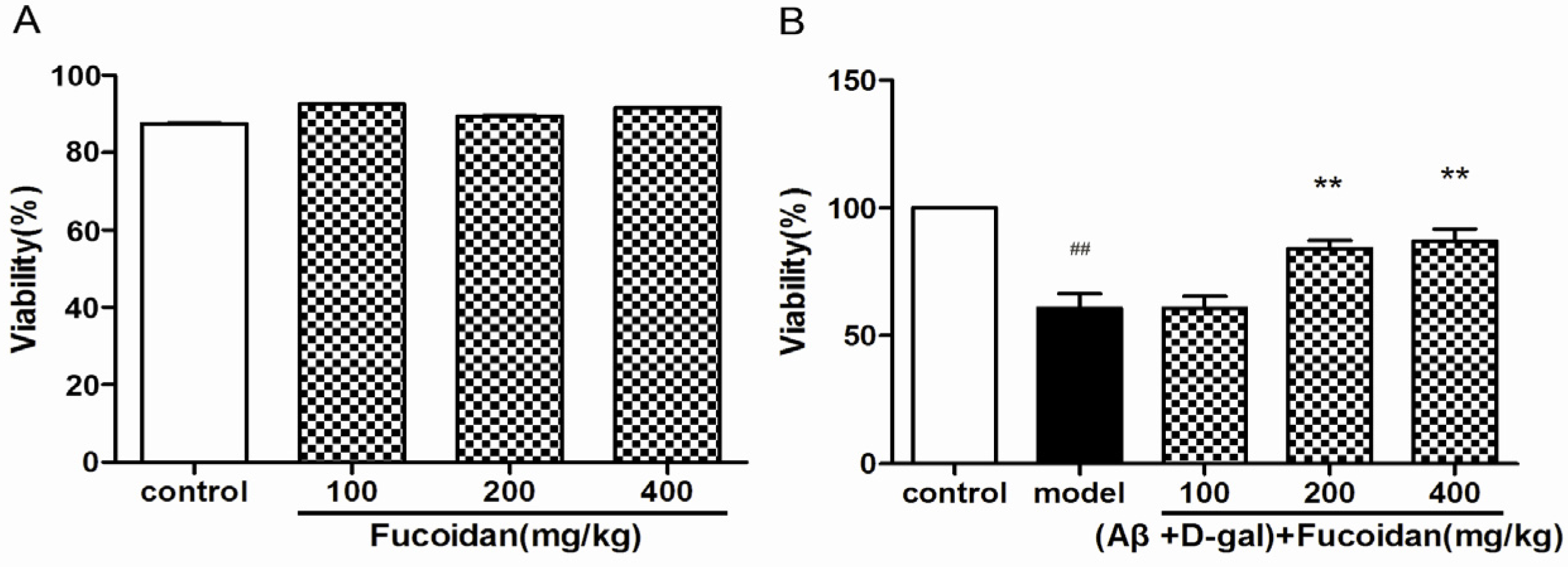
2.1. Fucoidan Improves the Viability of PC12 Cells Damaged by Aβ and d-Gal
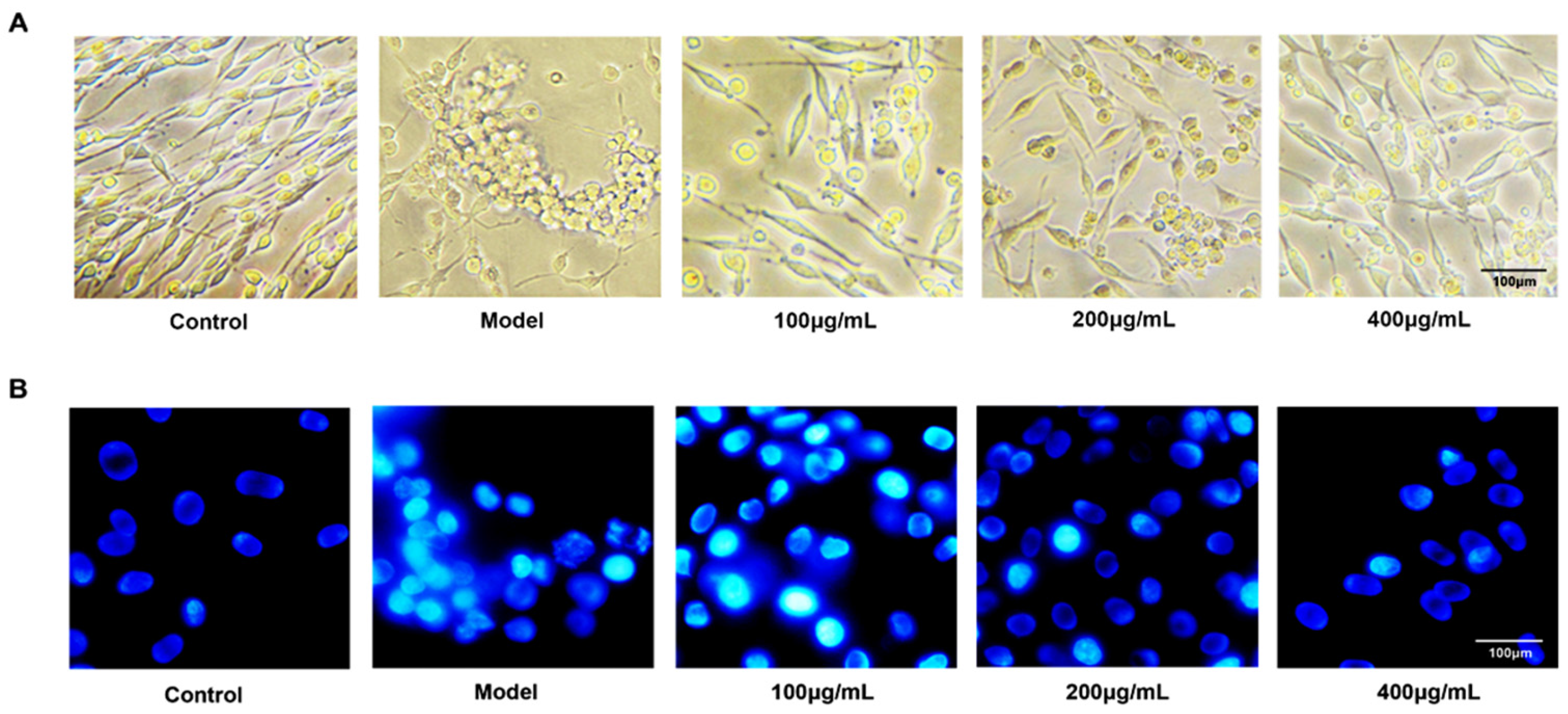
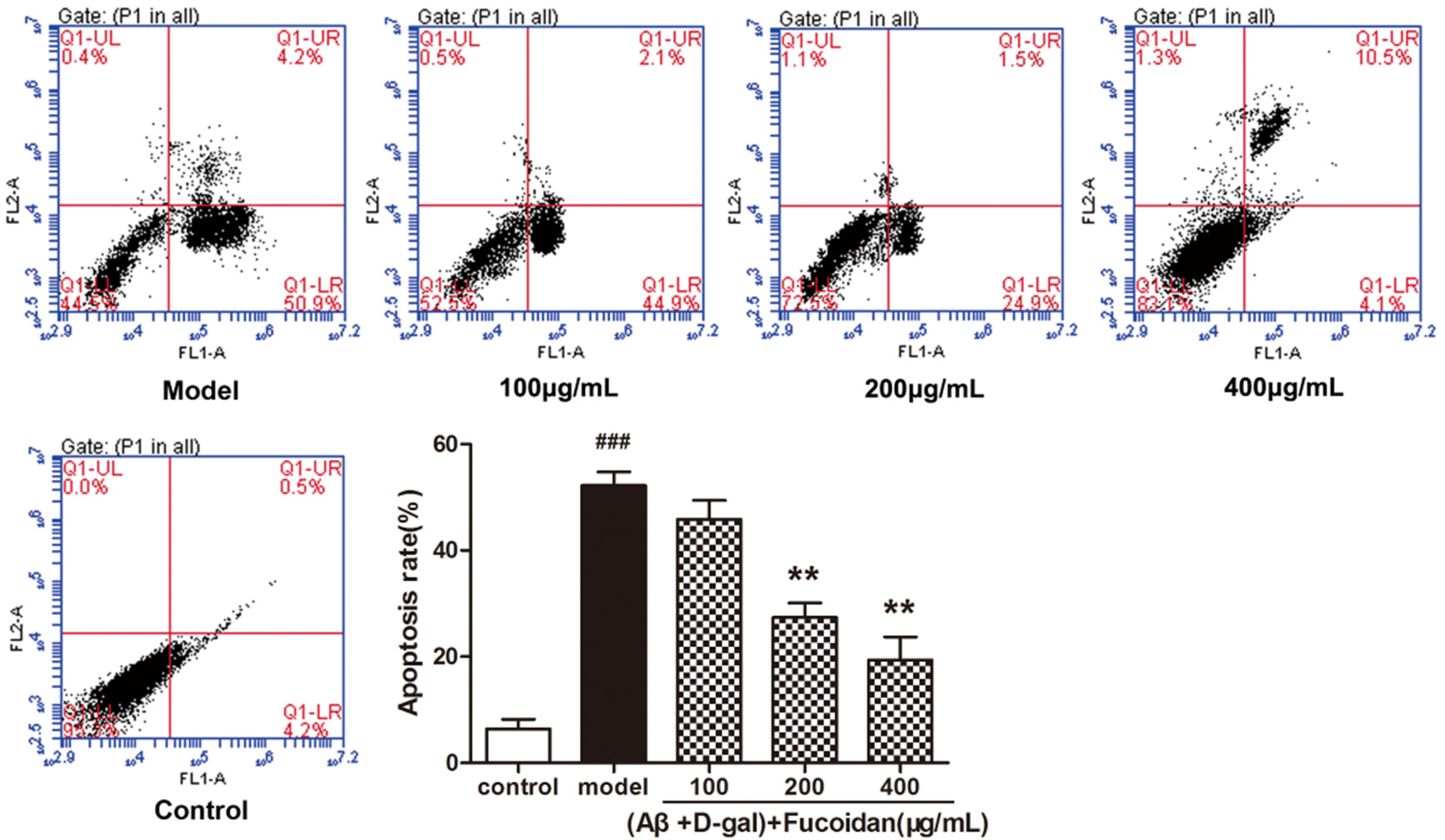
2.2. Fucoidan Prevents Apoptosis in PC12 Cell Induced by Aβ and d-Gal
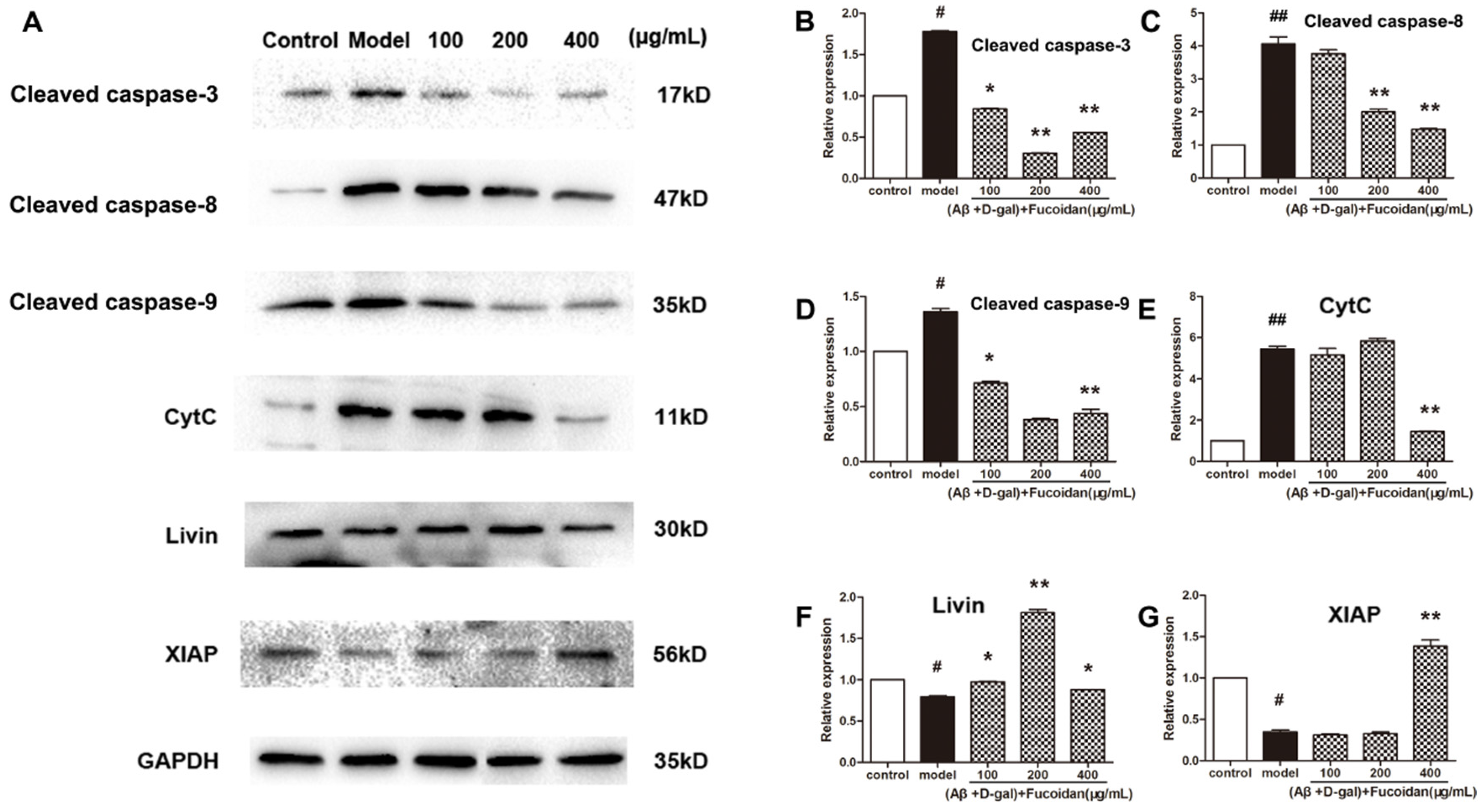
2.3. Effect of Fucoidan on Levels of Apoptosis-Related Proteins in PC12 Cells
2.4. Effect of Fucoidan on SOD Activity and GSH Content in PC12 Cells
2.5. Effects of Fucoidan on Learning and Memory Improvements of AD Model Mice
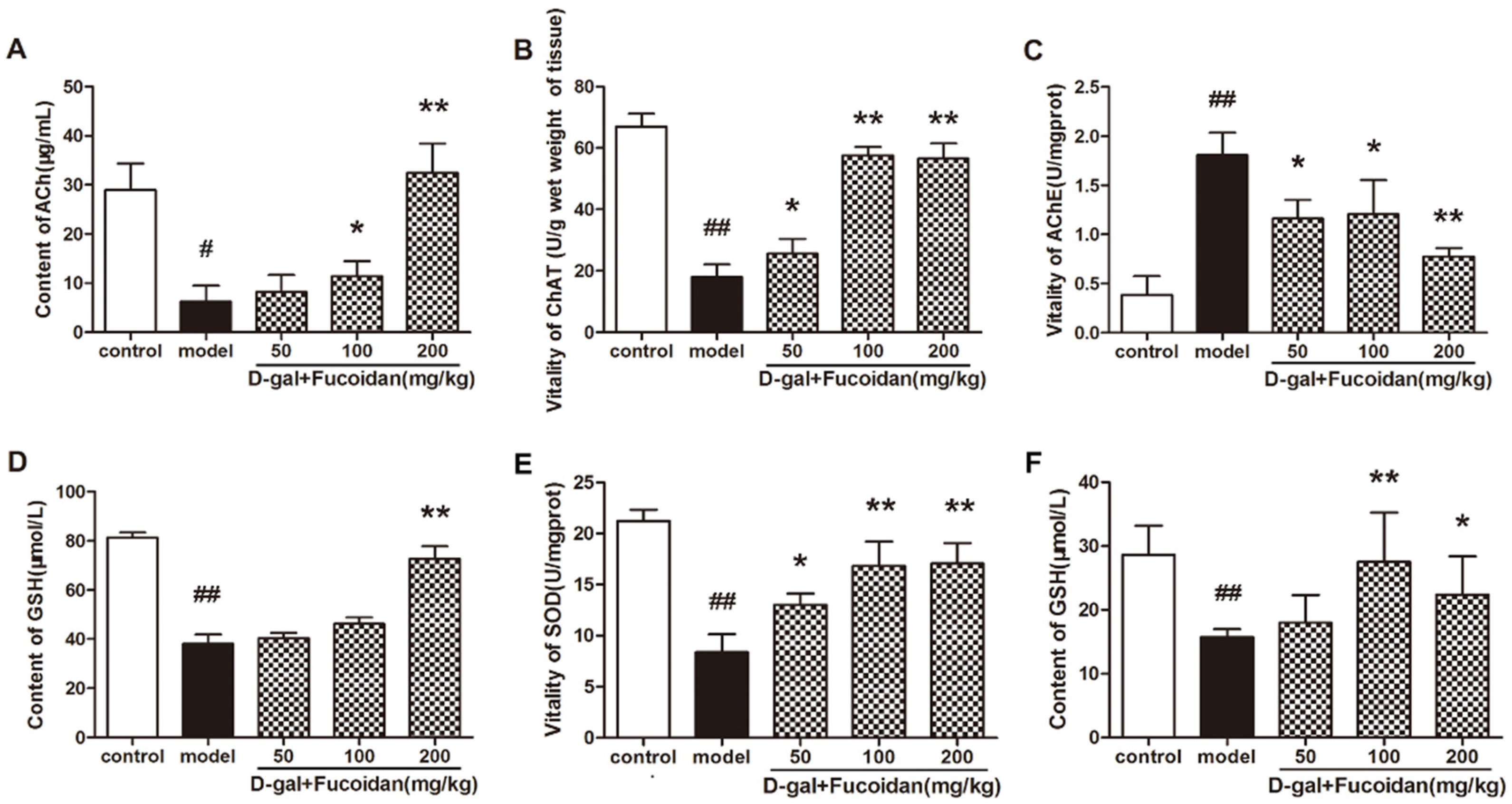
2.6. Effect of Fucoidan on Cholinergic System and Biochemistry Indexes In Vivo
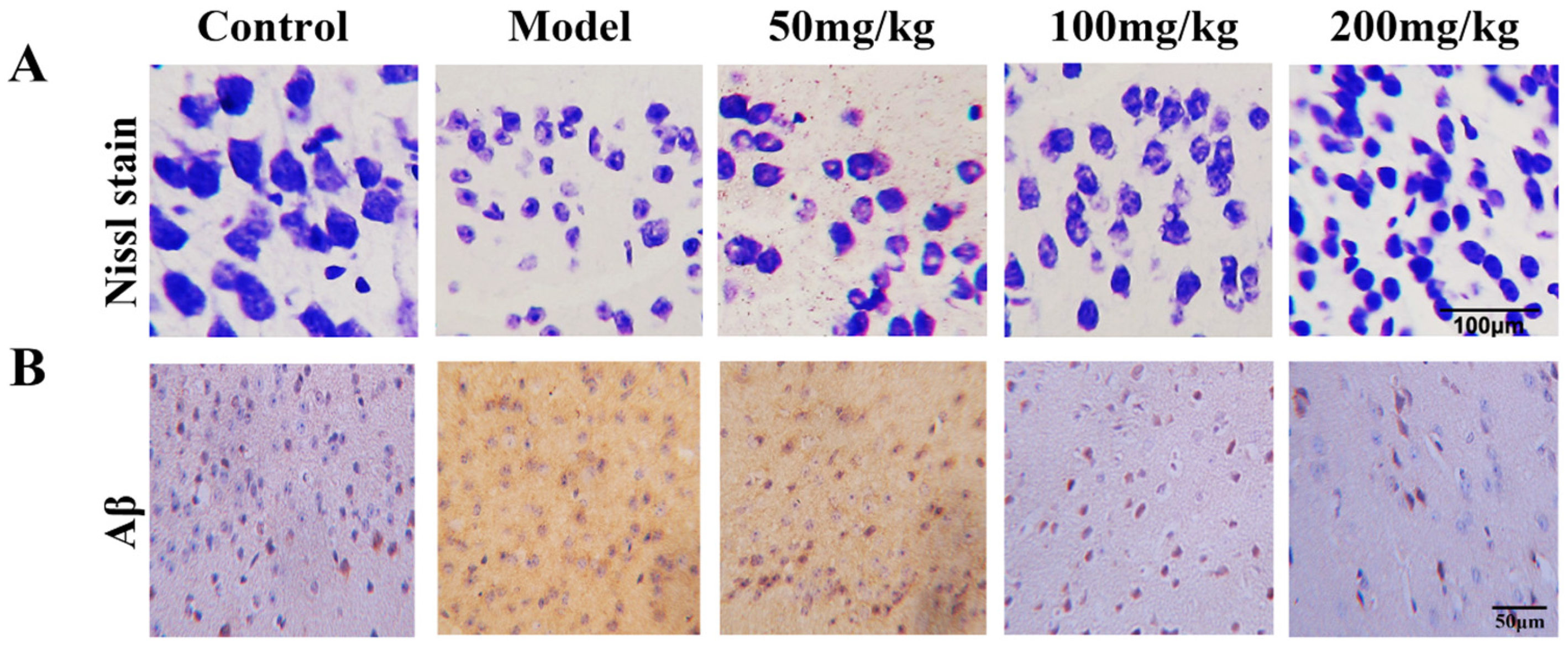
2.7. Fucoidan Protects Hippocampal Neurons Impaired by d-Gal In Vivo
2.8. Discussion
3. Materials and Methods
3.1. Reagents and Antibodies
3.2. Purification and Analyses of Fucoidan
3.3. Aβ25–35 and d-Gal Preparation
3.4. Animals
3.5. Cell Culture and Treatment
3.6. Cell Viability Assay
3.7. Light Microscope Observation
3.8. Hoechst 33258 Staining
3.9. Apoptosis Assay
3.10. Western Blot Analysis
3.11. Biochemistry Index of PC12 Cells
3.12. Treatments in Animals
3.13. Morris Water Maze
3.14. Cholinergic System and Biochemistry Estimations
3.15. Nissl and Immunohistochemistry Staining
3.16. Statistical Analysis
4. Conclusion
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| AD | Alzheimer’s disease; |
| Aβ25–35 | beta-amyloid 25–35; |
| d-Gal | d-galactose; |
| ACh | acetylcholine; |
| AChE | acetylcholine esterase; |
| ChAT | choline acetyl transferase; |
| Cyt C | cytochrome C; |
| GSH | glutathione; |
| FCM | flow cytometry; |
| SOD | superoxide dismutase; |
| XIAP | X-linked IAP; |
| IAP | inhibitor of apoptosis protein. |
References
- Kumar, A.; Nisha, C.M.; Silakari, C.; Sharma, I.; Anusha, K.; Gupta, N.; Nair, P.; Tripathi, T.; Kumar, A. Current and novel therapeutic molecules and targets in Alzheimer’s disease. J. Formos. Med. Assoc. 2015, 115, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.C.H.; Wang, B.; Ng, S.Y.; Lee, T.F. Effects of ginseng saponins on β-amyloid-induced amnesia in rats. J. Ethnopharmacol. 2006, 103, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Cuenco, K.T.; Green, R.C.; Zhang, J.; Lunetta, K.; Erlich, P.M.; Cupples, L.A.; Farrer, A.; DeCarli, C. Magnetic resonance imaging traits in siblings discordant for Alzheimer disease. J. Neuroimaging 2008, 18, 268–275. [Google Scholar] [CrossRef] [PubMed]
- Hardy, J.; Selkoe, D.J. The amyloid hypothesis of Alzheimer’s disease: Progress and problems on the road to therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.H.; Surh, Y.J. Protective effect of resveratrol on beta-amyloid induced oxidative PC12 cell death. Free Radic. Biol. Med. 2003, 34, 1100–1110. [Google Scholar] [CrossRef]
- Kayed, R.; Head, E.; Thompson, J.L.; McIntire, T.M.; Milton, S.C.; Cotman, C.W.; Glabe, C.C. Common structure of soluble amyloid oligomers implies common mechanism of pathogenesis. Science 2003, 300, 486–489. [Google Scholar] [CrossRef] [PubMed]
- Cappai, R.; Barnham, K.J. Delineating the mechanism of Alzheimer’s disease A beta peptide neurotoxicity. Neurochem. Res. 2008, 33, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Castellani, R.J.; Lee, H.G.; Zhu, X.W.; Perry, G.; Smith, M.A. Alzheimer disease pathology as a host response. J. Neuropathol. Exp. Neurol. 2008, 67, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Aisa, Y.; Miyakawa, Y.; Nakazato, T.; Shibata, H.; Saito, K.; Ikeda, Y.; Kizaki, M. Fucoidan induces apoptosis of human HS-sultan cells accompanied by activation of caspase-3 and down-regulation of ERK pathways. Am. J. Hematol. 2005, 78, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Xian, Y.F.; Lin, Z.X.; Zhao, M.; Mao, Q.Q.; Ip, S.P.; Che, C.T. Uncaria rhynchophylla ameliorates cognitive deficits induced by d-galactose in mice. Planta Med. 2011, 77, 1977–1983. [Google Scholar] [CrossRef] [PubMed]
- Jayaprakasam, B.; Padmanabhan, K.; Nair, M.G. Withanamides in Withania somniferafruit protect PC-12 cells from beta-amyloid responsible for Alzheimer’s disease. Phytother. Res. 2010, 24, 859–863. [Google Scholar] [PubMed]
- Fu, Z.; Yang, J.; Wei, Y.; Li, J. Effects of piceatannol and pterostilbene against β-amyloid-induced apoptosis on the PI3K/Akt/Bad signaling pathway in PC12 cells. Food Funct. 2016, 7, 1014–1023. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.L. Alzheimer’s disease: Channeling APP to non-amyloidogenic processing. Biochem. Biophys. Res. Commun. 2005, 331, 375–378. [Google Scholar] [CrossRef] [PubMed]
- Hardingham, G.E.; Fukunaga, Y.; Bading, H. Extrasynaptic NMDARs oppose synaptic NMDARs by triggering CREB shut-off and cell death pathways. Nat. Neurosci. 2002, 5, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, M. Plasticity, hippocampal place cells, and cognitive maps. Arch. Neurol. 2001, 58, 875–881. [Google Scholar] [CrossRef]
- Léveillé, F.; Papadia, S.; Fricker, M.; Bell, K.F.; Soriano, F.X.; Martel, M.A.; Puddifoot, C.; Habel, M.; Wyllie, D.J.; Ikonomidou, C.; et al. Suppression of the intrinsic apoptosis pathway by synaptic activity. J. Neurosci. 2010, 30, 2623–2635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lau, D.; Bading, H. Synaptic activity-mediated surpression of p53 and induction of nuclear calcium-regulated neuroprotective genes promote survival through inhibitor of mitochondrial permeability transition. J. Neurosci. 2009, 29, 4420–4429. [Google Scholar] [CrossRef] [PubMed]
- Sultan, A.; Nesslany, F.; Violet, M.; Bégard, S.; Loyens, A.; Talahari, S.; Mansuroglu, Z.; Marzin, D.; Sergeant, N.; Humez, S.; et al. Nuclear Tau, a key player in neuronal DNA protection. J. Biol. Chem. 2011, 286, 4566–4575. [Google Scholar] [CrossRef] [PubMed]
- Simon, H.U.; Haj-Yehia, A.; Levi-Schaffer, F. Role of reactive oxygen species (ROS) in apoptosis induction. Apoptosis 2000, 5, 415–418. [Google Scholar] [CrossRef] [PubMed]
- Budihardjo, I.; Oliver, H.; Lutter, M.; Luo, X.; Wang, X. Biochemical pathways of caspase activationduring apoptosis. Annu. Rev. Cell Dev. Biol. 1999, 15, 269–290. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.W.; Won, T.J.; Kim, H.R.; Lee, D.H.; Hwang, K.W.; Park, S.Y. Protective effect of chlorogenic acid against Aβ-induced neurotoxicity. Biomol. Ther. 2011, 19, 181–186. [Google Scholar] [CrossRef]
- Lu, J.H.; Guo, J.; Yang, W.H. Effects of green tea polyphenol on the behaviour of Alzheimer’s disease like mice induced by d-galactose and Aβ 25-35. J. Chin. Med. Mater. 2006, 29, 352–354. [Google Scholar]
- Pompl, P.N.; Yemul, S.; Xiang, Z.; Ho, L.; Haroutunian, V.; Purohit, D.; Mohs, R.; Pasinetti, G.M. Caspase gene expression in the brain as a function of the clinical progression of Alzheimer disease. Arch. Neurol. 2003, 60, 369–376. [Google Scholar] [CrossRef] [PubMed]
- Christine, S.; Thomas, L.D.; Anu, S.; Christian, B.; Wolfgang, B.; Kurt, J.; Hans, L. Activation ofcaspase-3 in single neurons and autophagic granules ofgranulovacuolar degeneration in Alzheimer’s disease. Am. J. Pathol. 1999, 155, 1459–1466. [Google Scholar]
- Lu, D.C.; Rabizadeh, S.; Chandra, S.; Shayya, R.F.; Ellerby, L.M.; Ye, X.; Salvesen, G.S.; Koo, E.H.; Bredesen, D.E. A second cytotoxic proteolyticpeptide derived from amyloid beta-protein precursor. Nat. Med. 2000, 6, 397–404. [Google Scholar] [PubMed]
- Crack, P.J.; Cimdins, K.; Ali, U.; Hertzog, P.J.; Iannello, R.C. Lack of glutathione peroxidase-1 exacerbates A[beta]-mediated neurotoxicity in cortical neurons. J. Neural Transm. 2006, 113, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Reddy, P.H. Amyloid precursor protein-mediated free radicals and oxidative damage: Implications for the development and progression of Alzheimer’s disease. J. Neurochem. 2006, 96, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, Q.; Zhang, Z.; Li, Z. Antioxidant activity of sulfated polysaccharide fractions extracted from Laminaria japonica. Int. J. Biol. Macromol. 2008, 42, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Freitas, R.M. Lipoic acid increases hippocampal choline acetyltransferase and acetylcholinesterase activities and improvement memory in epileptic rats. Neurochem. Res. 2010, 35, 162–170. [Google Scholar] [CrossRef] [PubMed]
- You, S.G.; Yang, C.; Lee, H.; Lee, B.Y. Molecular characteristics of partially hydrolyzed fucoidans from sporophyll of Undaria pinnatifidaand theirin vitroanticancer activity. Food Chem. 2010, 119, 554–559. [Google Scholar] [CrossRef]
- Yang, L.L.; Wang, P.S.; Wang, H.X. Fucoidan derived from Undaria pinnatifida induces apoptosis in human hepatocellular carcinoma SMMC-7721 cells via the ROS-Mediated mitochondrial pathway. Mar. Drugs 2013, 11, 1961–1976. [Google Scholar] [CrossRef] [PubMed]
- Hao, L.N.; Zhang, Q.Z.; Yu, T.G.; Cheng, Y.N.; Ji, S.L. Antagonistic effects of ultra-low-molecular-weight heparin on Aβ-induced apoptosis in cultured rat cortical neurons. Brain Res. 2011, 1368, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Morris, R. Developments of a water-maze procedure for studying spatial learning in the rat. J. Neurosci. Methods 1984, 11, 47–60. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Dose (mg/kg) | Escape Latency Time (s) | |||||
|---|---|---|---|---|---|---|---|
| 1 Day | 2 Day | 3 Day | 4 Day | 5 Day | 6 Day | ||
| Control | 27.10 ± 8.66 | 23.07 ± 10.87 | 12.53 ± 7.03 | 11.93 ± 4.37 | 7.08 ± 1.53 | 7.36 ± 5.25 | |
| Model | 0 | 73.51 ± 19.99 # | 63.82 ± 23.19 | 54.09 ± 33.65 | 49.95 ± 29.40 | 47.19 ± 18.17 # | 38.62 ± 19.51 # |
| Fucoidan | 50 | 46.54 ± 9.56 | 41.54 ± 10.65 | 41.06 ± 15.55 | 36.24 ± 18.35 * | 39.74 ± 17.68 * | 28.52 ± 16.51 * |
| 100 | 39.46 ± 15.27 * | 23.89 ± 13.83 ** | 15.95 ± 11.02 ** | 14.78 ± 11.98 ** | 14.24 ± 8.37 ** | 11.1 ± 7.09 ** | |
| 200 | 41.60 ± 16.70 * | 21.61 ± 10.47 ** | 21.16 ± 12.28 ** | 18.12 ± 9.09 ** | 11.54 ± 4.58 ** | 9.87 ± 3.76 ** | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, H.; Gao, Z.; Zheng, L.; Zhang, C.; Liu, Z.; Yang, Y.; Teng, H.; Hou, L.; Yin, Y.; Zou, X. Protective Effects of Fucoidan on Aβ25–35 and d-Gal-Induced Neurotoxicity in PC12 Cells and d-Gal-Induced Cognitive Dysfunction in Mice. Mar. Drugs 2017, 15, 77. https://doi.org/10.3390/md15030077
Wei H, Gao Z, Zheng L, Zhang C, Liu Z, Yang Y, Teng H, Hou L, Yin Y, Zou X. Protective Effects of Fucoidan on Aβ25–35 and d-Gal-Induced Neurotoxicity in PC12 Cells and d-Gal-Induced Cognitive Dysfunction in Mice. Marine Drugs. 2017; 15(3):77. https://doi.org/10.3390/md15030077
Chicago/Turabian StyleWei, Hengyun, Zixiang Gao, Luping Zheng, Cuili Zhang, Zundong Liu, Yazong Yang, Hongming Teng, Lin Hou, Yuling Yin, and Xiangyang Zou. 2017. "Protective Effects of Fucoidan on Aβ25–35 and d-Gal-Induced Neurotoxicity in PC12 Cells and d-Gal-Induced Cognitive Dysfunction in Mice" Marine Drugs 15, no. 3: 77. https://doi.org/10.3390/md15030077
APA StyleWei, H., Gao, Z., Zheng, L., Zhang, C., Liu, Z., Yang, Y., Teng, H., Hou, L., Yin, Y., & Zou, X. (2017). Protective Effects of Fucoidan on Aβ25–35 and d-Gal-Induced Neurotoxicity in PC12 Cells and d-Gal-Induced Cognitive Dysfunction in Mice. Marine Drugs, 15(3), 77. https://doi.org/10.3390/md15030077