Degradation of Marine Algae-Derived Carbohydrates by Bacteroidetes Isolated from Human Gut Microbiota





Abstract
:1. Introduction
2. Results and Discussion
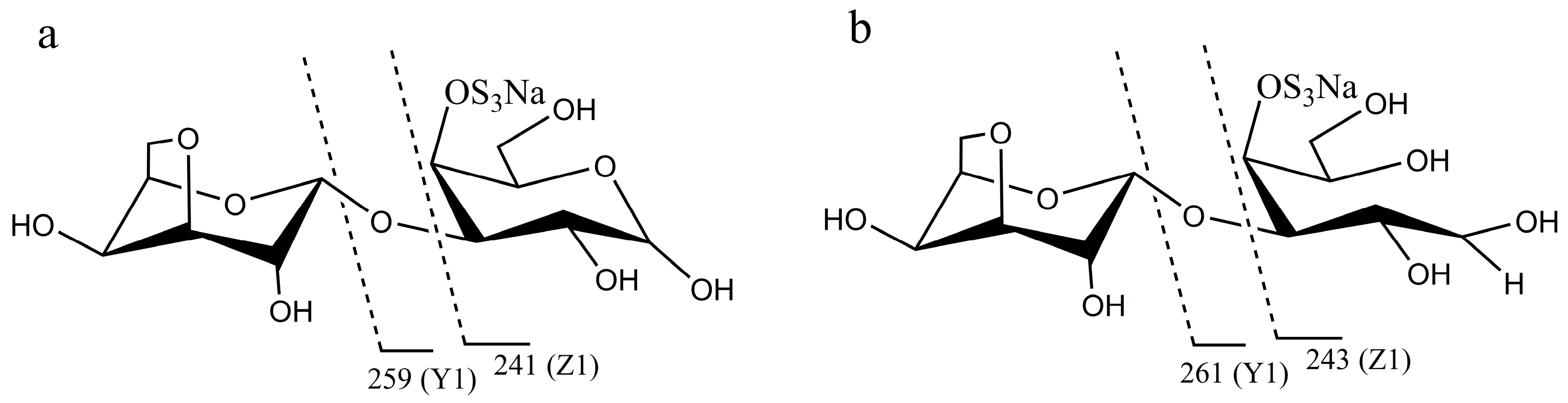
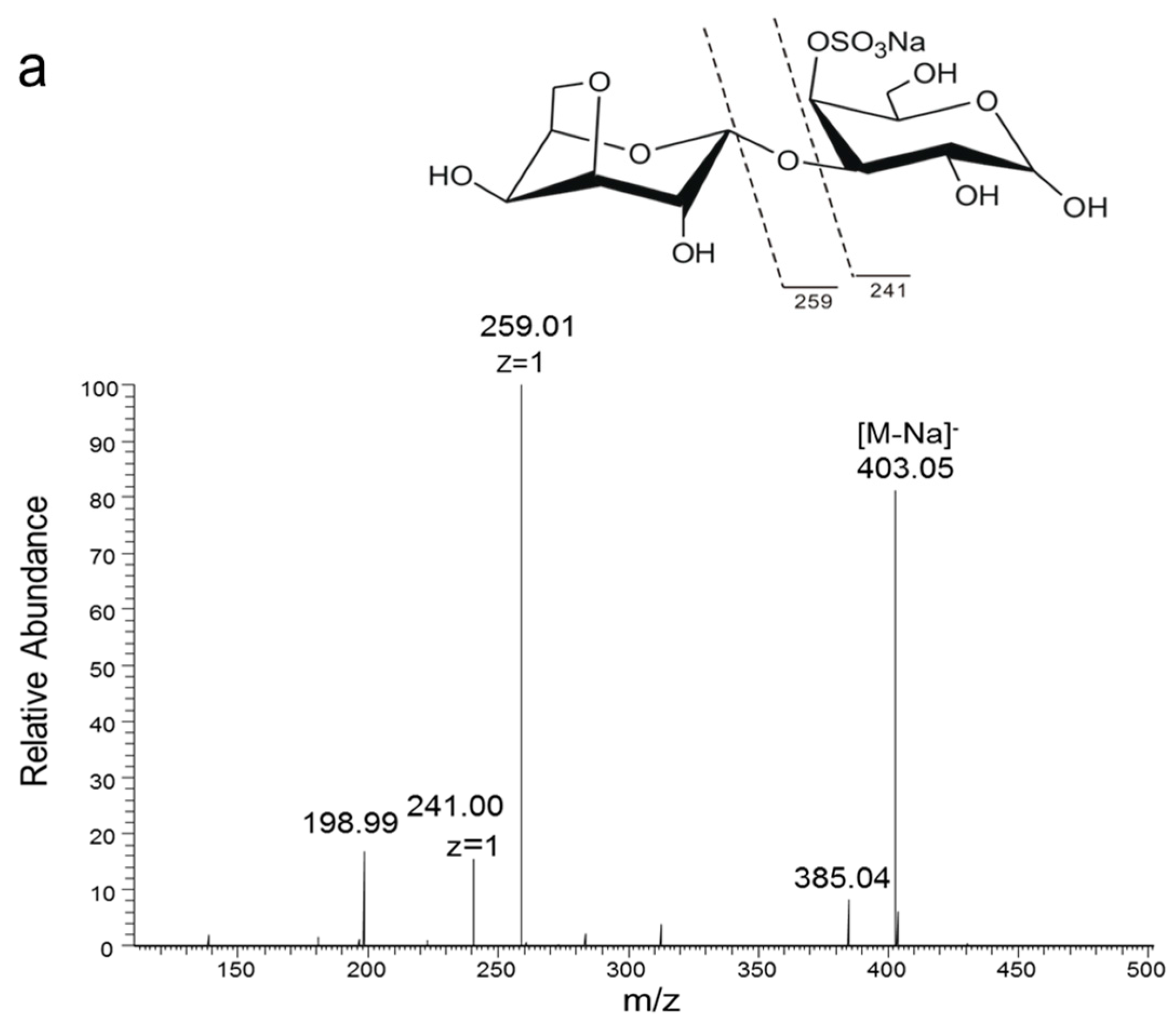
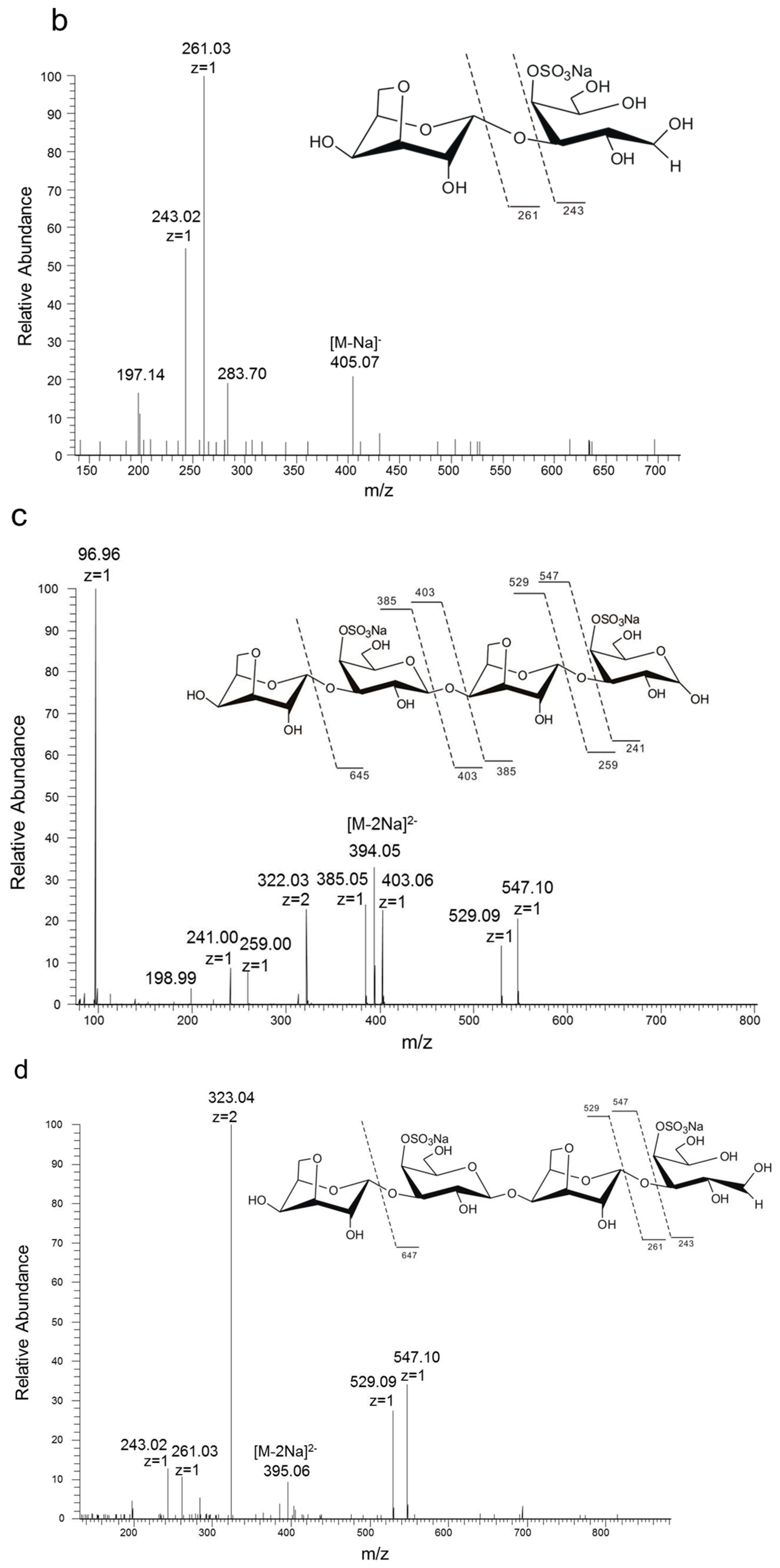
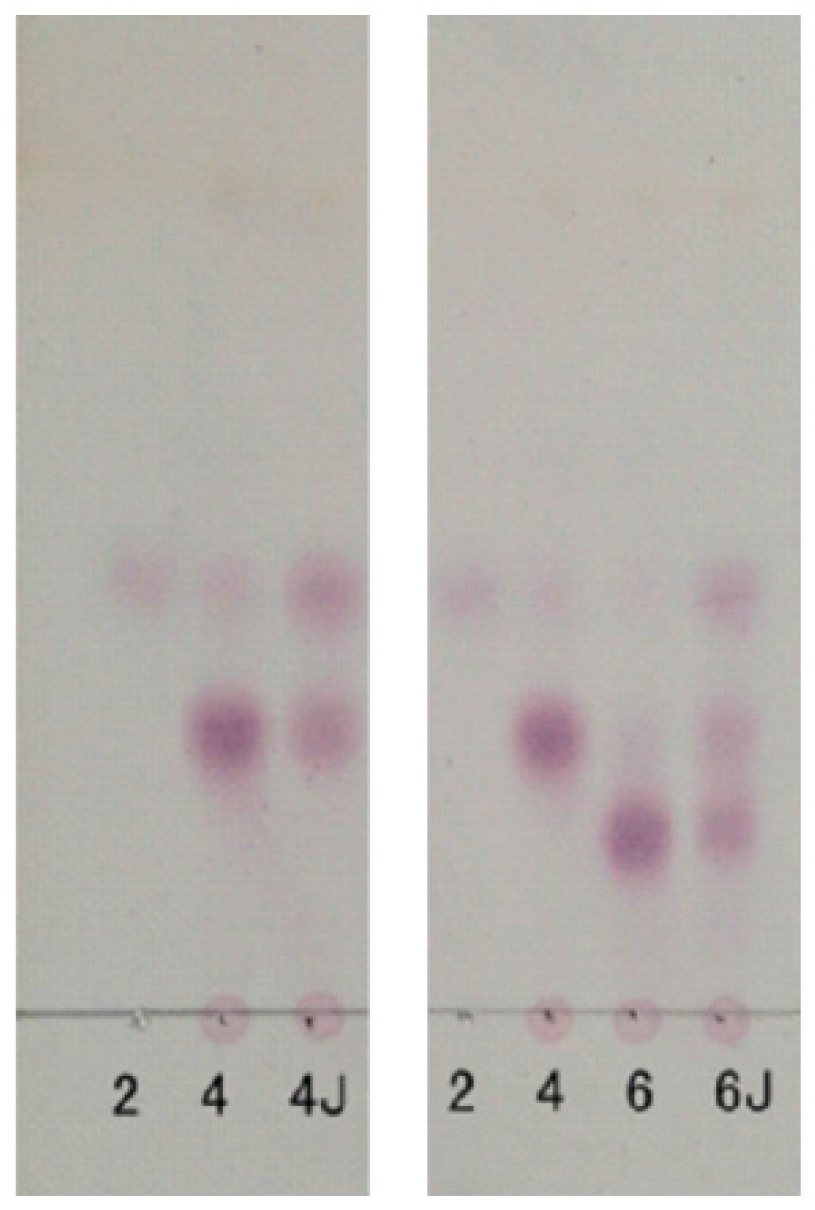
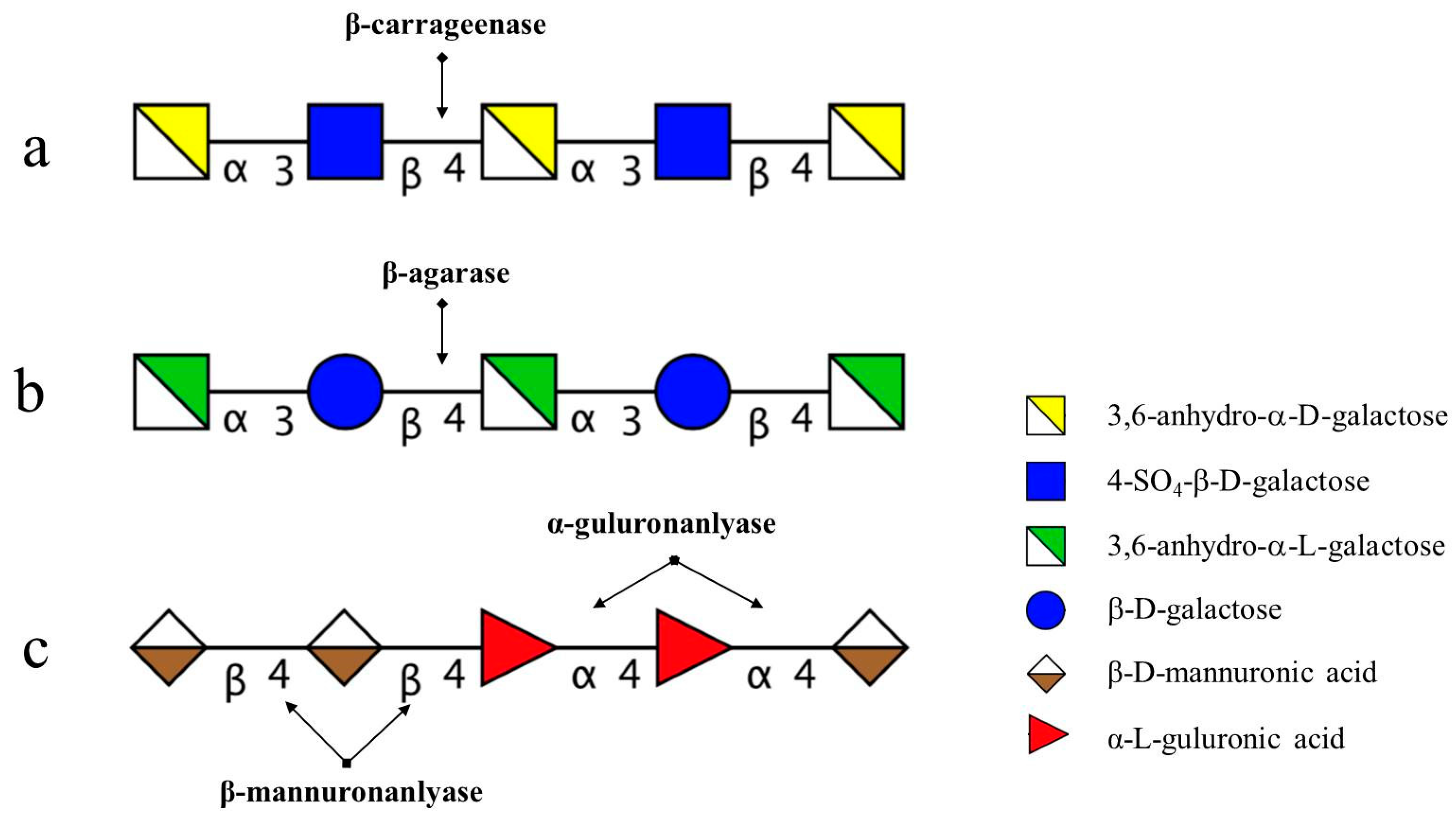
2.1. Chemical Structures of the Products Generated by the Degradation of κ-Carrageenan Oligosaccharides by B. xylanisolvens and E. coli
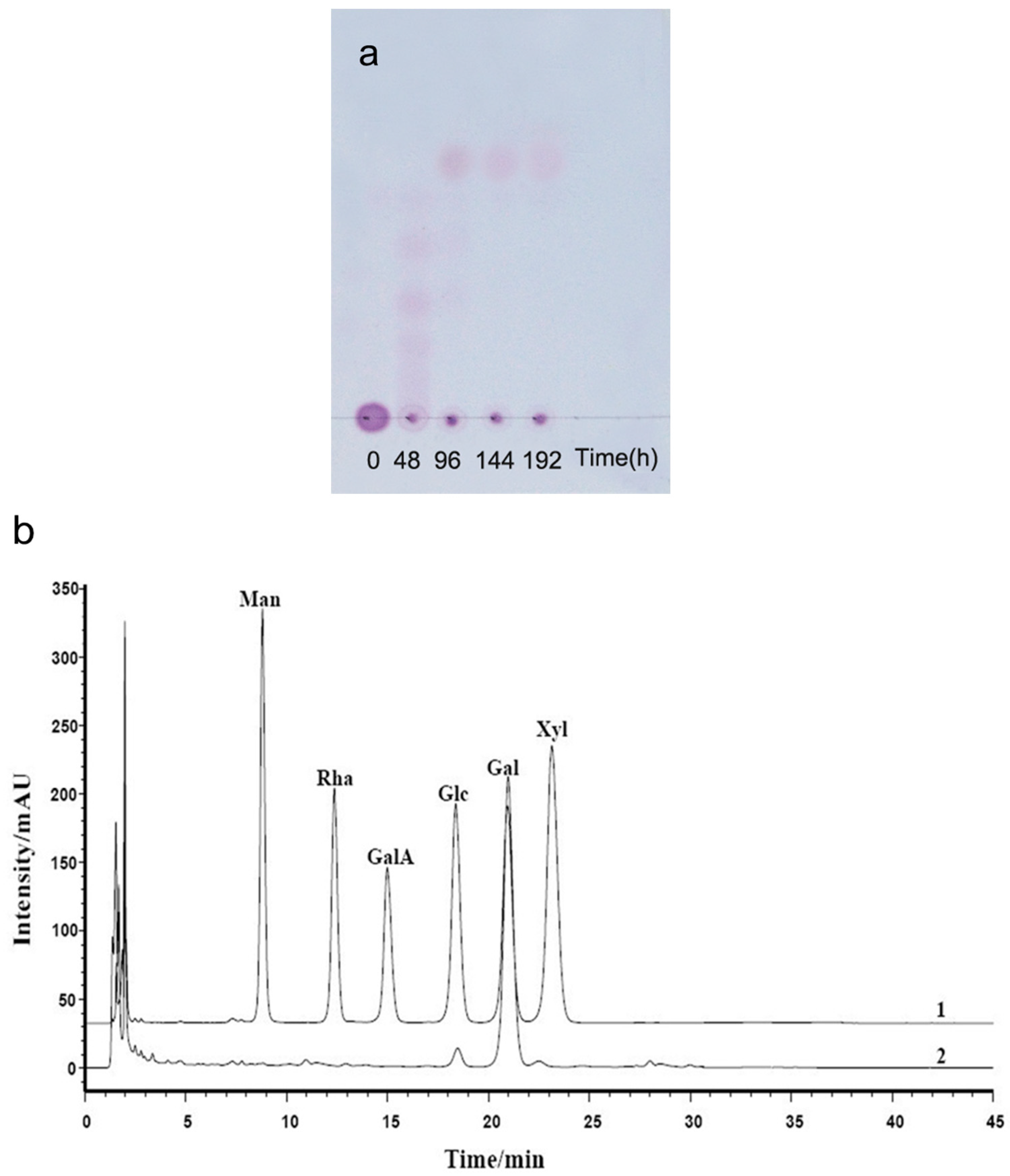
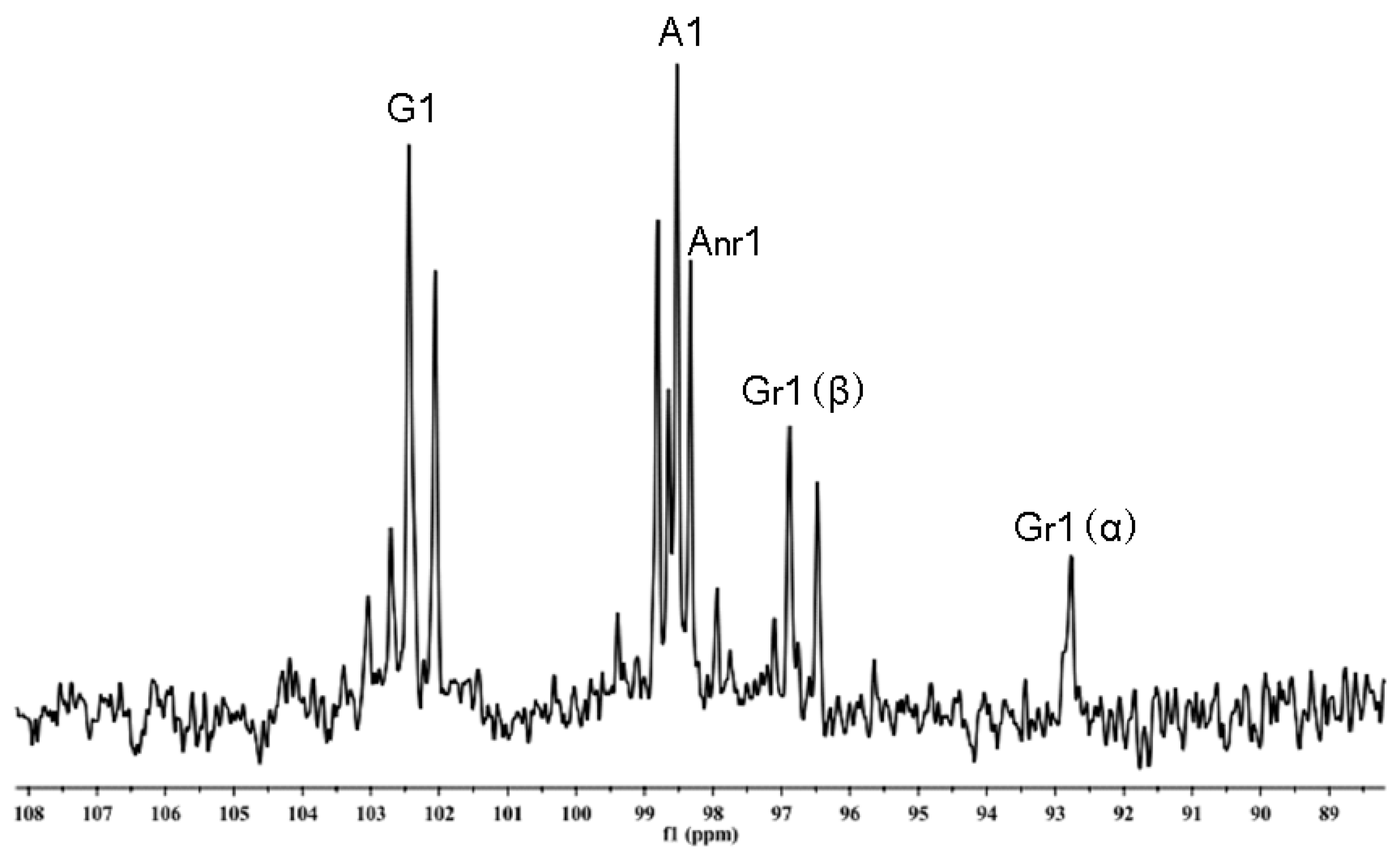
2.2. Chemical Structures of the Intermediates Produced by the Degradation of AP by B. uniformis L8
2.3. Chemical Structures of the Intermediates Produced by the Degradation of Alg, MO and GO by B. ovatus G19
3. Experimental Section
3.1. Polysaccharide and Oligosaccharide Materials
3.2. Bacteroidetes Material
3.3. Degradation of Marine Carbohydrates by Human Gut Bacteria Isolates
3.4. General Experimental Procedures
3.5. Oligosaccharide Reduction
4. Conclusions
Acknowledgments
Author Contribution
Conflicts of Interest
References
- Yagi, H.; Fujise, A.; Itabashi, N.; Ohshiro, T. Purification and characterization of a novel alginate lyase from the marine bacterium Cobetia sp. NAP1 isolated from brown algae. Biosci. Biotechnol. Biochem. 2016, 80, 2338–2346. [Google Scholar] [CrossRef] [PubMed]
- Shan, D.; Ying, J.; Li, X.; Gao, Z.; Wei, G.; Shao, Z. Draft genome sequence of the carrageenan-degrading bacterium Cellulophaga sp. strain KL-A, isolated from decaying marine algae. Genome Announc. 2014, 2. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Lin, B.K.; Xu, Y.; Zhong, M.Q.; Liu, G.M. Production and purification of agarase from a marine agarolytic bacterium Agarivorans sp. HZ105. J. Appl. Microbiol. 2009, 106, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Hehemann, J.-H.; Kelly, A.G.; Pudlo, N.A.; Martens, E.C.; Boraston, A.B. Bacteria of the human gut microbiome catabolize red seaweed glycans with carbohydrate-active enzyme updates from extrinsic microbes. Proc. Natl. Acad. Sci. USA 2012, 109, 19786–19791. [Google Scholar] [CrossRef] [PubMed]
- Hehemann, J.-H.; Correc, G.; Barbeyron, T.; Helbert, W.; Czjzek, M.; Michel, G. Transfer of carbohydrate-active enzymes from marine bacteria to Japanese gut microbiota. Nature 2010, 464, 908–912. [Google Scholar] [CrossRef] [PubMed]
- Kaoutari, A.E.; Armougom, F.; Gordon, J.I.; Raoult, D.; Henrissat, B. The abundance and variety of carbohydrate-active enzymes in the human gut microbiota. Nat. Rev. Microbiol. 2013, 11, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Li, R.; Raes, J.; Arumugam, M.; Burgdorf, K.S.; Manichanh, C.; Nielsen, T.; Pons, N.; Levenez, F.; Yamada, T.; et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature 2010, 464, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, G.; Zhu, L.; Yin, Y.; Zhao, X.; Xiang, C.; Yu, G.; Wang, X. Isolation and characterization of an agaro-oligosaccharide (AO)-hydrolyzing bacterium from the gut microflora of Chinese individuals. PLoS ONE 2014, 9, e91106. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Li, G.; Shang, Q.; Chen, X.; Liu, W.; Pi, X.; Zhu, L.; Yin, Y.; Yu, G.; Wang, X. In vitro fermentation of alginate and its derivatives by human gut microbiota. Anaerobe 2016, 39, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Li, M. The Degradation and Utilization Study of Agarose,κ-carrageenan and Their Oligosaccharides by Human Gut Microbiota. Ph.D. Thesis, Ocean University of China, Qingdao, China, 2014. [Google Scholar]
- Yang, B. Preparation and Sequence Analysis of Oligosaccharides and Neoglycolipids Probes from Marine-Derived Sulfated Galactan & Construction of Oligosaccharide-Chips. Ph.D. Thesis, Ocean University of China, Qingdao, China, 2009. [Google Scholar]
- Ji, M. Seaweed Chemistry; Science Press: Beijing, China, 1997. [Google Scholar]
- Yang, B.; Yu, G.; Zhao, X.; Jiao, G.; Ren, S.; Chai, W. Mechanism of mild acid hydrolysis of galactan polysaccharides with highly ordered disaccharide repeats leading to a complete series of exclusively odd-numbered oligosaccharides. FEBS J. 2009, 276, 2125–2137. [Google Scholar] [CrossRef] [PubMed]
- Ostgaard, K.; Knutsen, S.H.; Dyrset, N.; Aasen, I.M. Production and characterization of guluronate lyase from Klebsiella pneumoniae for applications in seaweed biotechnology. Enzyme Microb. Technol. 1993, 15, 756–763. [Google Scholar] [CrossRef]
- Linker, A.; Evans, L.R. Isolation and characterization of an alginase from mucoid strains of Pseudomonas aeruginosa. J. Bacteriol. 1984, 159, 958–964. [Google Scholar] [PubMed]
- Sawabe, T.; Ohtsuka, M.; Ezura, Y. Novel alginate lyases from marine bacterium Alteromonas sp. strain H-4. Carbohydr. Res. 1997, 304, 69–76. [Google Scholar] [CrossRef]
- Lei, F.; Yin, Y.; Wang, Y.; Deng, B.; Yu, H.D.; Li, L.; Xiang, C.; Wang, S.; Zhu, B.; Wang, X. Higher-level production of volatile fatty acids in vitro by chicken gut microbiotas than by human gut microbiotas as determined by functional analyses. Appl. Environ. Microbiol. 2012, 78, 5763–5772. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Zhao, X.; Lv, Y.; Li, M.; Liu, X.; Li, G.; Yu, G. Structural and compositional characteristics of hybrid carrageenans from red algae Chondracanthus chamissoi. Carbohydr. Polym. 2012, 89, 914–919. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yu, G.; Zhao, X.; Liu, H.; Guan, H.; Lawson, A.M.; Chai, W. Sequence analysis of alginate-derived oligosaccharides by negative-ion electrospray tandem mass spectrometry. J. Am. Soc. Mass Spectrom. 2006, 17, 621–630. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Zhao, X.; Yang, B.; Ren, S.; Guan, H.; Zhang, Y.; Lawson, A.M.; Chai, W. Sequence determination of sulfated carrageenan-derived oligosaccharides by high-sensitivity negative-ion electrospray tandem mass spectrometry. Anal. Chem. 2006, 78, 8499–8505. [Google Scholar] [CrossRef] [PubMed]
- Jiao, G.; Yu, G.; Wang, W.; Zhao, X.; Zhang, J.; Ewart, S.H. Properties of polysaccharides in several seaweeds from Atlantic Canada and their potential anti-influenza viral activities. J. Ocean Univ. China 2012, 11, 205–212. [Google Scholar] [CrossRef]
- Chai, W.; Luo, J.; Lim, C.K.; Lawson, A.M. Characterization of heparin oligosaccharide mixtures as ammonium salts using electrospray mass spectrometry. Anal. Chem. 1998, 70, 2060–2066. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhang, P.; Hao, C.; Zhang, X.-E.; Cui, Z.-Q.; Guan, H.-S. In vitro inhibitory effect of carrageenan oligosaccharide on influenza A H1N1 virus. Antivir. Res. 2011, 92, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Sun, Y.; Xin, H.; Zhang, Y.; Li, Z.; Xu, Z. In vivo antitumor and immunomodulation activities of different molecular weight lambda-carrageenans from Chondrus ocellatus. Pharmacol. Res. 2004, 50, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Hao, J.; Zhang, L.; Zhao, X.; He, X.; Li, M.; Zhao, X.; Wu, J.; Qiu, P.; Yu, G. Activated AMPK explains hypolipidemic effects of sulfated low molecular weight guluronate on HepG2 cells. Eur. J. Med. Chem. 2014, 85, 304–310. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fractions | Found Ions (Charge) | Calculated Mol Mass (Na Form) | Assignment | |
|---|---|---|---|---|
| DP | Sequences | |||
| A 1 | 403.06 (−1) | 426.05 | 2 | A-G4S |
| A 2 | 322.03 (−2) | 690.05 | 3 | G4S-A-G4S |
| A 3 | 394.05 (−2) | 834.10 | 4 | A-G4S-A-G4S |
| A 4 | 343.03 (−3) | 1098.09 | 5 | G4S-A-G4S-A-G4S |
| A 5 | 391.06 (−3) | 1245.18 | 6 | A-G4S-A-G4S-A-G4S |
| Fraction | Found Ions (Charge) | Calculated Mol Mass (H Form) | Assignment | |
|---|---|---|---|---|
| DP | Sequences | |||
| B1 | 325.11 * (+1) | 324.11 | 2 | A-G |
| B2 | 653.19 ** (+1) | 630.20 | 4 | A-G-A-G |
| B3 | 959.29 ** (+1) | 936.30 | 6 | A-G-A-G-A-G |
| B4 | 1265.38 ** (+1) | 1242.39 | 8 | A-G-A-G-A-G- A-G |
| Compound | G1 | Gr1 | Gnr1 | A1 | Ar1 | Anr1 |
|---|---|---|---|---|---|---|
| neoagaroligosaccharides a | 102.4 | 92.8(α) 96.8(β) | - | 98.5 | - | 98.3 |
| neoagarotetraose a Anr-Gnr-Ar-Gr | 102.4 | 92.8(α) 96.8(β) | - | 98.5 | - | 98.3 |
| agaroligosaccharides b | 102.6 | - | 102.6 | 98.4 | 90.4 | |
| agarotetraose b Gnr-Anr-Gr-Ar | 102.6 | - | 102.5 | 98.5 | 63.3 | |
| AP-L8 | 102.44 | 92.77(α) 96.88(β) | 98.53 | - | 98.33 |
| Fraction | Found Ions (Charge) | Calculated Mol Mass (H Form) | Assignment | |
|---|---|---|---|---|
| DP | Sequences | |||
| C1 | 351.05 (−1) | 351.06 | 2 | ∆NN |
| C2 | 527.08 (−1) | 527.09 | 3 | ∆NNN |
| C3 | 351.05 (−2), 703.11 (−1) | 704.13 | 4 | ∆NNNN |
| C4 | 439.06 (−2), 879.13 (−1) | 880.16 | 5 | ∆NNNNN |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, M.; Shang, Q.; Li, G.; Wang, X.; Yu, G. Degradation of Marine Algae-Derived Carbohydrates by Bacteroidetes Isolated from Human Gut Microbiota. Mar. Drugs 2017, 15, 92. https://doi.org/10.3390/md15040092
Li M, Shang Q, Li G, Wang X, Yu G. Degradation of Marine Algae-Derived Carbohydrates by Bacteroidetes Isolated from Human Gut Microbiota. Marine Drugs. 2017; 15(4):92. https://doi.org/10.3390/md15040092
Chicago/Turabian StyleLi, Miaomiao, Qingsen Shang, Guangsheng Li, Xin Wang, and Guangli Yu. 2017. "Degradation of Marine Algae-Derived Carbohydrates by Bacteroidetes Isolated from Human Gut Microbiota" Marine Drugs 15, no. 4: 92. https://doi.org/10.3390/md15040092
APA StyleLi, M., Shang, Q., Li, G., Wang, X., & Yu, G. (2017). Degradation of Marine Algae-Derived Carbohydrates by Bacteroidetes Isolated from Human Gut Microbiota. Marine Drugs, 15(4), 92. https://doi.org/10.3390/md15040092