
Outdoor Cultivation of Marine Diatoms for Year-Round Production of Biofuels


Abstract
:1. Introduction
2. Results
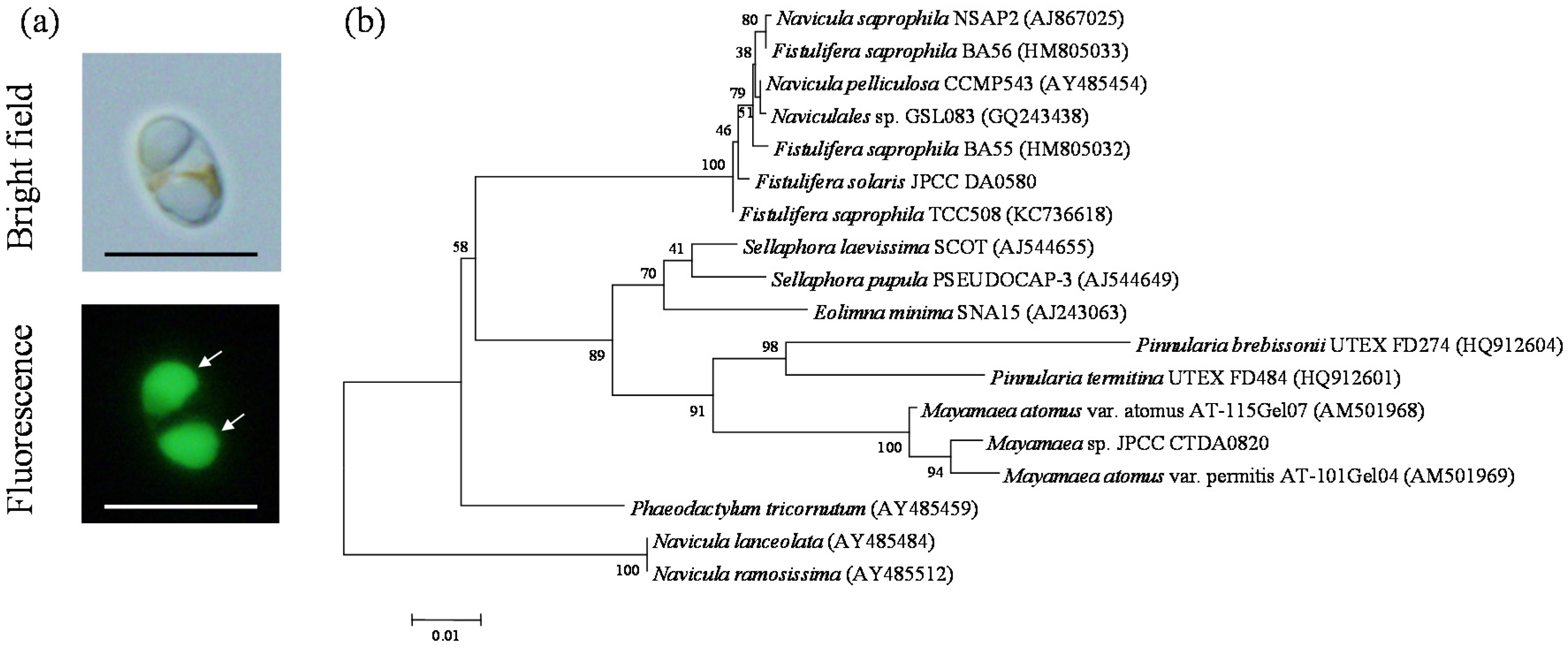
2.1. Molecular Phylogenetic Analysis of Mayamaea sp. JPCC CTDA0820
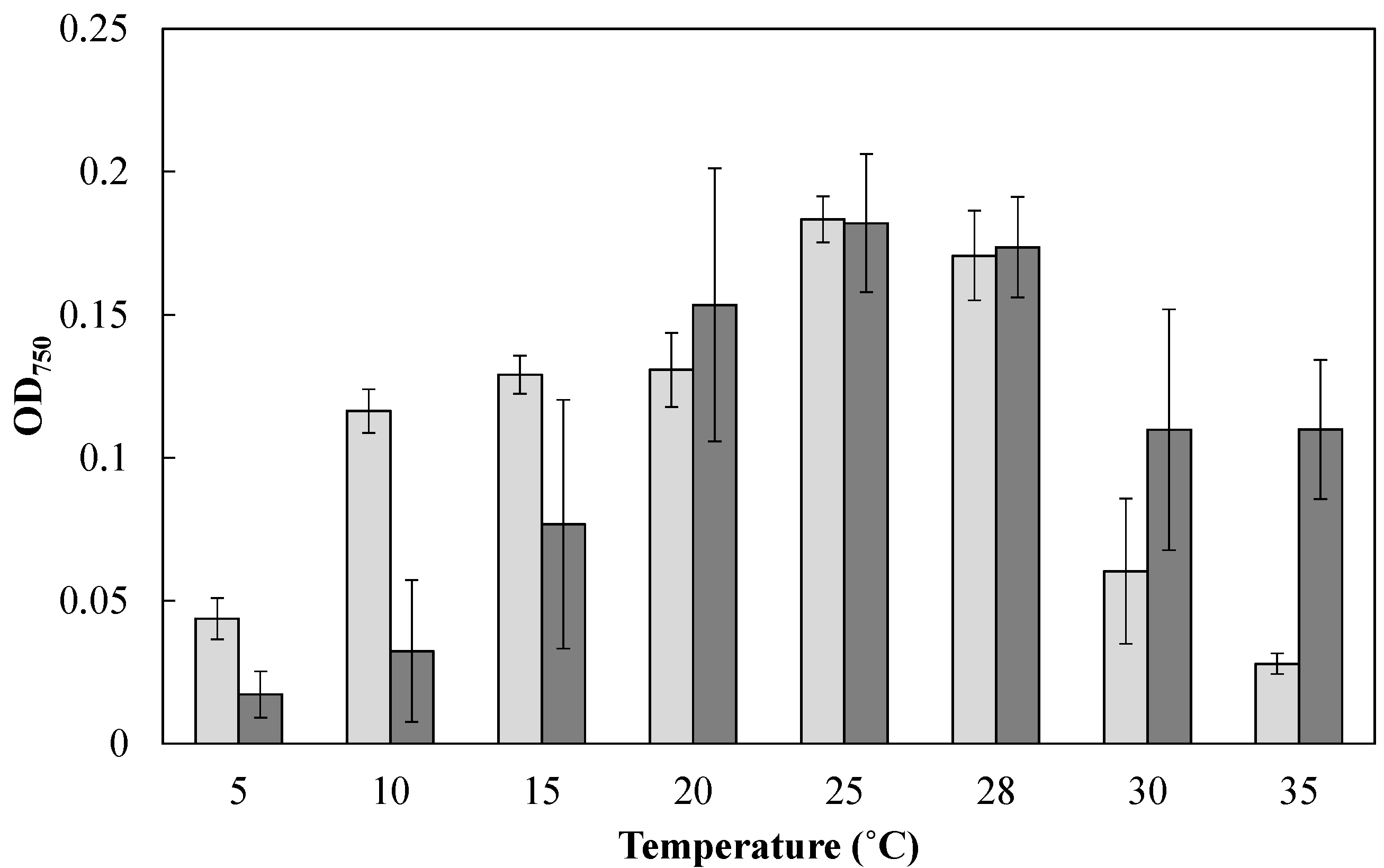
2.2. Characterization of Mayamaea sp. JPCC CTDA0820 by Laboratory-Scale Indoor Cultivation
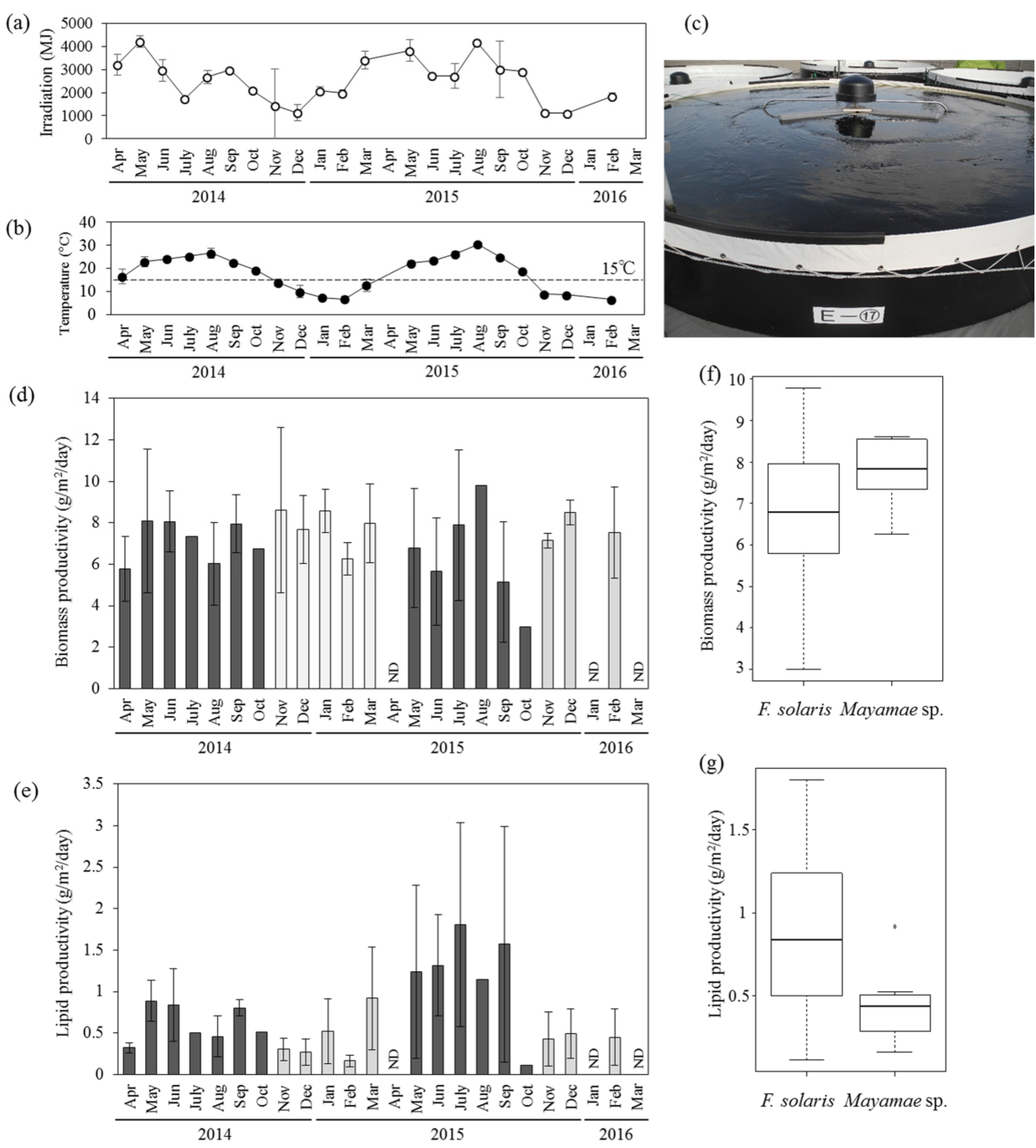
2.3. Year-Round Production of Diatom Lipids with Conbined Use of F. Solaris and Mayamaea sp. JPCC CTDA0820 in Open-Pond Bioreactors
2.4. Comparison of Biomass and Lipid Production in Different Types of Outdoor Bioreactors
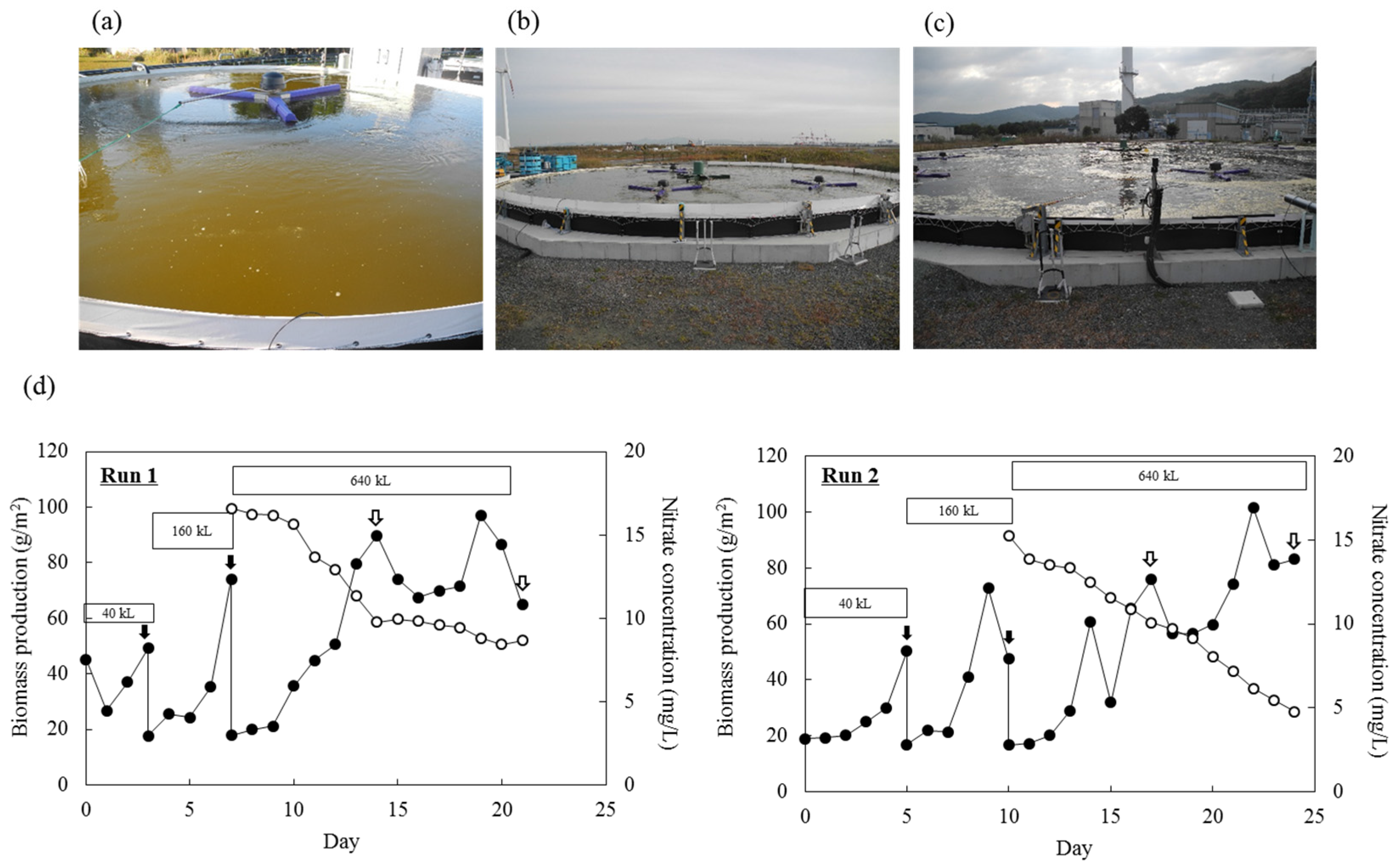
2.5. Pilot-Scale Outdoor Cultivation of Mayamaea sp. JPCC CTDA0820
3. Discussion
4. Materials and Methods
4.1. Isolation of Diatoms
4.2. Phylogenetic Analysis
4.3. Laboratory-Scale Indoor Cultivation
4.4. Quantification and Statistics Analysis of Biomass and Oil Productivity
4.5. Bioreactor Design and Culture Conditions for Outdoor Cultivation
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Koller, M.; Muhr, A.; Braunegg, G. Microalgae as versatile cellular factories for valued products. Algal Res. 2014, 6, 52–63. [Google Scholar] [CrossRef]
- Chisti, Y. Biodiesel from microalgae. Biotechnol. Adv. 2007, 25, 294–306. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.L.; Yasin, N.H.M.; Derek, C.J.C.; Lim, J.K. Microalgae as a sustainable energy source for biodiesel production: A review. Renew. Sustain. Energy Rev. 2011, 15, 584–593. [Google Scholar] [CrossRef]
- Schenk, P.M.; Thomas-Hall, S.R.; Stephens, E.; Marx, U.C.; Mussgnug, J.H.; Posten, C.; Kruse, O.; Hankamer, B. Second generation biofuels: High-efficiency microalgae for biodiesel production. Bioenerg. Res. 2008, 1, 20–43. [Google Scholar] [CrossRef]
- Smith, V.H.; Sturm, B.S.; Denoyelles, F.J.; Billings, S.A. The ecology of algal biodiesel production. Trends Ecol. Evol. 2010, 25, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Rodolfi, L.; Chini Zittelli, G.; Bassi, N.; Padovani, G.; Biondi, N.; Bonini, G.; Tredici, M.R. Microalgae for oil: Strain selection, induction of lipid synthesis and outdoor mass cultivation in a low-cost photobioreactor. Biotechnol. Bioeng. 2009, 102, 100–112. [Google Scholar] [CrossRef] [PubMed]
- Bechet, Q.; Munoz, R.; Shilton, A.; Guieysse, B. Outdoor cultivation of temperature-tolerant Chlorella sorokiniana in a column photobioreactor under low power-input. Biotechnol. Bioeng. 2013, 110, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Del Campo, J.A.; Rodriguez, H.; Moreno, J.; Vargas, M.A.; Rivas, J.; Guerrero, M.G. Lutein production by muriellopsis sp. In an outdoor tubular photobioreactor. J. Biotechnol. 2001, 85, 289–295. [Google Scholar] [CrossRef]
- Acien Fernandez, F.G.; Hall, D.O.; Canizares Guerrero, E.; Krishna Rao, K.; Molina Grima, E. Outdoor production of Phaeodactylum tricornutum biomass in a helical reactor. J. Biotechnol. 2003, 103, 137–152. [Google Scholar] [CrossRef]
- Zittelli, G.C.; Rodolfi, L.; Biondi, N.; Tredici, M.R. Productivity and photosynthetic efficiency of outdoor cultures of tetraselmis suecica in annular columns. Aquaculture 2006, 261, 932–943. [Google Scholar] [CrossRef]
- Raes, E.; Isdepsky, A.; Muylaert, K.; Borowitzka, M.; Moheimani, N. Comparison of growth of tetraselmis in a tubular photobioreactor (biocoil) and a raceway pond. J. Appl. Phycol. 2014, 26, 247–255. [Google Scholar] [CrossRef]
- Moheimani, N.R.; Borowitzka, M.A. The long-term culture of the coccolithophore Pleurochrysis carterae (haptophyta) in outdoor raceway ponds. J. Appl. Phycol. 2006, 18, 703–712. [Google Scholar] [CrossRef]
- Mehrabadi, A.; Farid, M.M.; Craggs, R. Variation of biomass energy yield in wastewater treatment high rate algal ponds. Algal Res. 2016, 15, 143–151. [Google Scholar] [CrossRef]
- Bondioli, P.; Della Bella, L.; Rivolta, G.; Chini Zittelli, G.; Bassi, N.; Rodolfi, L.; Casini, D.; Prussi, M.; Chiaramonti, D.; Tredici, M.R. Oil production by the marine microalgae Nannochloropsis sp. F&m-m24 and Tetraselmis suecica f&m-m33. Bioresour. Technol. 2012, 114, 567–572. [Google Scholar] [PubMed]
- Slegers, P.; Lösing, M.; Wijffels, R.; Van Straten, G.; Van Boxtel, A. Scenario evaluation of open pond microalgae production. Algal Res. 2013, 2, 358–368. [Google Scholar] [CrossRef]
- Sato, R.; Maeda, Y.; Yoshino, T.; Tanaka, T.; Matsumoto, M. Seasonal variation of biomass and oil production of the oleaginous diatom fistulifera sp. In outdoor vertical bubble column and raceway-type bioreactors. J. Biosci. Bioeng. 2014, 117, 720–724. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.W.; Hwangbo, K.; Lee, S.Y.; Choi, H.G.; Park, Y.I.; Liu, J.R.; Jeong, W.J. A new arctic chlorella species for biodiesel production. Bioresour. Technol. 2012, 125, 340–343. [Google Scholar] [CrossRef] [PubMed]
- Nelson, D.R.; Mengistu, S.; Ranum, P.; Celio, G.; Mashek, M.; Mashek, D.; Lefebvre, P.A. New lipid-producing, cold-tolerant yellow-green alga isolated from the rocky mountains of colorado. Biotechnol. Prog. 2013, 29, 853–861. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Hu, H. Screening high oleaginous chlorella strains from different climate zones. Bioresour. Technol. 2013, 144, 637–643. [Google Scholar] [CrossRef] [PubMed]
- Bidigare, R.; Ondrusek, M.E.; Kennicutt, M.C.; Iturriaga, R.; Harvey, H.R.; Hoham, R.W.; Macko, S. Evidence a photoprotective for secondary carotenoids of snow algae. J. Phycol. 1993, 29, 427–434. [Google Scholar] [CrossRef]
- Yoshimura, Y.; Kohshima, S.; Ohtani, S. A community of snow algae on a himalayan glacier: Change of algal biomass and community structure with altitude. Arct. Alp. Res. 1997, 126–137. [Google Scholar] [CrossRef]
- Liang, Y.; Maeda, Y.; Yoshino, T.; Matsumoto, M.; Tanaka, T. Profiling of fatty acid methyl esters from the oleaginous diatom fistulifera sp. strain JPCC DA0580 under nutrition-sufficient and -deficient conditions. J. Appl. Phycol. 2014, 26, 2295–2302. [Google Scholar] [CrossRef]
- Tanaka, T.; Maeda, Y.; Veluchamy, A.; Tanaka, M.; Abida, H.; Marechal, E.; Bowler, C.; Muto, M.; Sunaga, Y.; Tanaka, M.; et al. Oil accumulation by the oleaginous diatom Fistulifera solaris as revealed by the genome and transcriptome. Plant Cell 2015, 27, 162–176. [Google Scholar] [CrossRef] [PubMed]
- Nojima, D.; Yoshino, T.; Maeda, Y.; Tanaka, M.; Nemoto, M.; Tanaka, T. Proteomics analysis of oil body-associated proteins in the oleaginous diatom. J. Proteom. Res. 2013, 12, 5293–5301. [Google Scholar] [CrossRef] [PubMed]
- Maeda, Y.; Tateishi, T.; Niwa, Y.; Muto, M.; Yoshino, T.; Kisailus, D.; Tanaka, T. Peptide-mediated microalgae harvesting method for efficient biofuel production. Biotechnol. Biofuels 2016, 9, 10. [Google Scholar] [CrossRef] [PubMed]
- Muto, M.; Tanaka, M.; Liang, Y.; Yoshino, T.; Matsumoto, M.; Tanaka, T. Enhancement of glycerol metabolism in the oleaginous marine diatom Fistulifera solaris JPCC DA0580 to improve triacylglycerol productivity. Biotechnol. Biofuels 2015, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, M.; Nojima, D.; Ikeda, K.; Yoshino, T.; Tanaka, T. Application of cold-tolerant marine diatom, mayamaea sp. JPCC CTDA0820 to low-energy cultivation process for stable biodiesel production (in Japanese). J. Jpn. Inst. Energy 2015, 94, 1087–1091. [Google Scholar] [CrossRef]
- Guillard, R.R.; Ryther, J.H. Studies of marine planktonic diatoms. I. Cyclotella nana hustedt, and Detonula confervacea (cleve) gran. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Satoh, A.; Ichii, K.; Matsumoto, M.; Kubota, C.; Nemoto, M.; Tanaka, M.; Yoshino, T.; Matsunaga, T.; Tanaka, T. A process design and productivity evaluation for oil production by indoor mass cultivation of a marine diatom, fistulifera sp. JPCC DA0580. Bioresour.Technol. 2013, 137, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Souffreau, C.; Vanormelingen, P.; Verleyen, E.; Sabbe, K.; Vyverman, W. Tolerance of benthic diatoms from temperate aquatic and terrestrial habitats to experimental desiccation and temperature stress. Phycologia 2010, 49, 309–324. [Google Scholar] [CrossRef]
- Starks, T.L.; Shubert, L.E.; Trainor, F. Ecology of soil algae: A review. Phycologia 1981, 20, 65–80. [Google Scholar] [CrossRef]
- Maeda, K.; Owada, M.; Kimura, N.; Omata, K.; Karube, I. Co2 fixation from the flue gas on coal-fired thermal power plant by microalgae. Energy Convers. Manag. 1995, 36, 717–720. [Google Scholar] [CrossRef]
- Koller, M.; Salerno, A.; Tuffner, P.; Koinigg, M.; Böchzelt, H.; Schober, S.; Pieber, S.; Schnitzer, H.; Mittelbach, M.; Braunegg, G. Characteristics and potential of micro algal cultivation strategies: A review. J. Clean. Prod. 2012, 37, 377–388. [Google Scholar] [CrossRef]
- Rumin, J.; Bonnefond, H.; Saint-Jean, B.; Rouxel, C.; Sciandra, A.; Bernard, O.; Cadoret, J.-P.; Bougaran, G. The use of fluorescent nile red and bodipy for lipid measurement in microalgae. Biotechnol. Biofuels 2015, 8, 42. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal w and clustal x version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, M.; Mayama, S.; Nemoto, M.; Fukuda, Y.; Muto, M.; Yoshino, T.; Matsunaga, T.; Tanaka, T. Morphological and molecular phylogenetic analysis of the high triglyceride–producing marine diatom, Fistulifera solaris sp. Nov.(bacillariophyceae). Phycol. Res. 2014, 62, 257–268. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microalgae | Outdoor Bioreactors | Biomass Productivity [g/(m2 day)] | Lipid Productivity [g/(m2 day)] | Reference |
|---|---|---|---|---|
| Mayamaea sp. JPCC CTDA0820 | Open-pond | 8.62 | 0.92 | This study |
| Fistulifera solaris | Open-pond | 9.79 | 1.80 | This study |
| Chlorella sorokiniana | Column | 10 ± 2.2 | - | [7] |
| Nannochloropsis sp. F&M-M24 | Green Wall Panels | 9.9 | 6.5 | [14] |
| Tetraselmis suecica F&M-M33 | Green Wall Panels | 7.6 | 1.7 | [14] |
| Tetraselmis sp. MUR-233 | Open-pond | 7.2 ± 0.4 | 3.0 ± 0.6 | [11] |
| Tetraselmis sp. MUR-233 | Coiled tube | 7.8 ± 0.2 | 5.0±0.4 | [11] |
| Pleurochrysis carterae | Raceway | 3.20–33.68 | 1.54–11.41 | [12] |
| Microalgae in wastewater treatment high rate algal ponds | 5.9 | 1.42 | [13] | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matsumoto, M.; Nojima, D.; Nonoyama, T.; Ikeda, K.; Maeda, Y.; Yoshino, T.; Tanaka, T. Outdoor Cultivation of Marine Diatoms for Year-Round Production of Biofuels. Mar. Drugs 2017, 15, 94. https://doi.org/10.3390/md15040094
Matsumoto M, Nojima D, Nonoyama T, Ikeda K, Maeda Y, Yoshino T, Tanaka T. Outdoor Cultivation of Marine Diatoms for Year-Round Production of Biofuels. Marine Drugs. 2017; 15(4):94. https://doi.org/10.3390/md15040094
Chicago/Turabian StyleMatsumoto, Mitsufumi, Daisuke Nojima, Tomomi Nonoyama, Kiichi Ikeda, Yoshiaki Maeda, Tomoko Yoshino, and Tsuyoshi Tanaka. 2017. "Outdoor Cultivation of Marine Diatoms for Year-Round Production of Biofuels" Marine Drugs 15, no. 4: 94. https://doi.org/10.3390/md15040094
APA StyleMatsumoto, M., Nojima, D., Nonoyama, T., Ikeda, K., Maeda, Y., Yoshino, T., & Tanaka, T. (2017). Outdoor Cultivation of Marine Diatoms for Year-Round Production of Biofuels. Marine Drugs, 15(4), 94. https://doi.org/10.3390/md15040094