
A Review Study on Macrolides Isolated from Cyanobacteria


Abstract
:1. Introduction
2. Anti-Neoplastic Property of Cyanobacterium-Derived Macrolides on Different Cell Lines
3. Antibacterial Activity
4. Effects of Cyanobacterium-Derived Macrolides on Animals
5. Other Bioactivity
6. Conclusions
Acknowledgments
Conflicts of Interest
References
- Capper, A.; Erickson, A.A.; Ritson-Williams, R.; Becerro, M.A.; Arthur, K.A.; Paul, V.J. Palatability and chemical defences of benthic cyanobacteria to a suite of herbivores. J. Exp. Mar. Biol. Ecol. 2016, 474, 100–108. [Google Scholar] [CrossRef]
- Tan, L.T. Bioactive natural products from marine cyanobacteria for drug discovery. Phytochemistry 2007, 68, 954–979. [Google Scholar] [CrossRef] [PubMed]
- Napolitano, J.G.; Daranas, A.H.; Norte, M.; Fernández, J.J. Marine macrolides, a promising source of antitumor compounds. Anti-Cancer Agent Med. Chem. 2009, 9, 122–137. [Google Scholar] [CrossRef]
- Kollár, P.; Rajchard, J.; Balounová, Z.; Pazourek, J. Marine natural products: bryostatins in preclinical and clinical studies. Pharm. Biol. 2014, 52, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Belakhov, V.V.; Garabadzhiu, A.V. Polyene macrolide antibiotics: mechanisms of inactivation, ways of stabilization, and methods of disposal of unusable drugs (review). Russ. J. Gen. Chem. 2015, 85, 2985–3001. [Google Scholar] [CrossRef]
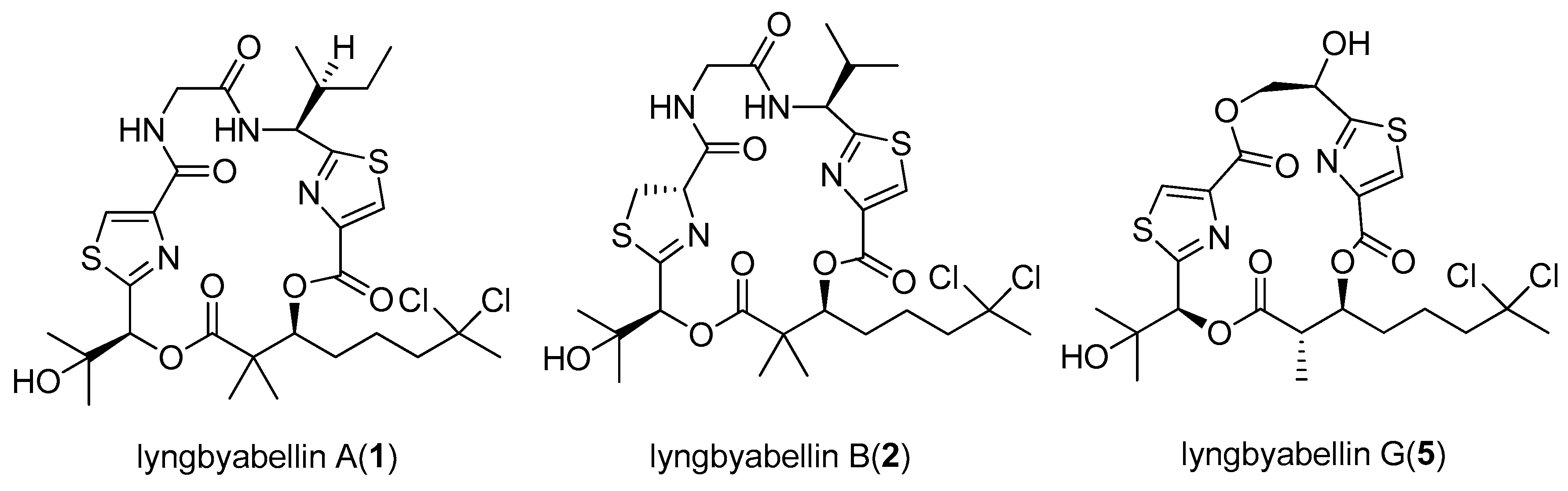
- Luesch, H.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J.; Mooberry, S.L. Isolation, structure determination, and biological activity of lyngbyabellin A from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2000, 63, 611–615. [Google Scholar] [CrossRef] [PubMed]
- Gunasekera, S.P.; Li, Y.; Ratnayake, R.; Luo, D.M.; Lo, J.; Reibenspies, J.H.; Xu, Z.S.; Clare-Salzler, M.J.; Ye, T.; Paul, V.J.; Luesch, H. Discovery, total synthesis and key structural elements for the immunosuppressive activity of cocosolide, a symmetrical glycosylated macrolide dimer from marine cyanobacteria. Chem. Eur. J. 2016, 22, 8158–8166. [Google Scholar] [CrossRef] [PubMed]
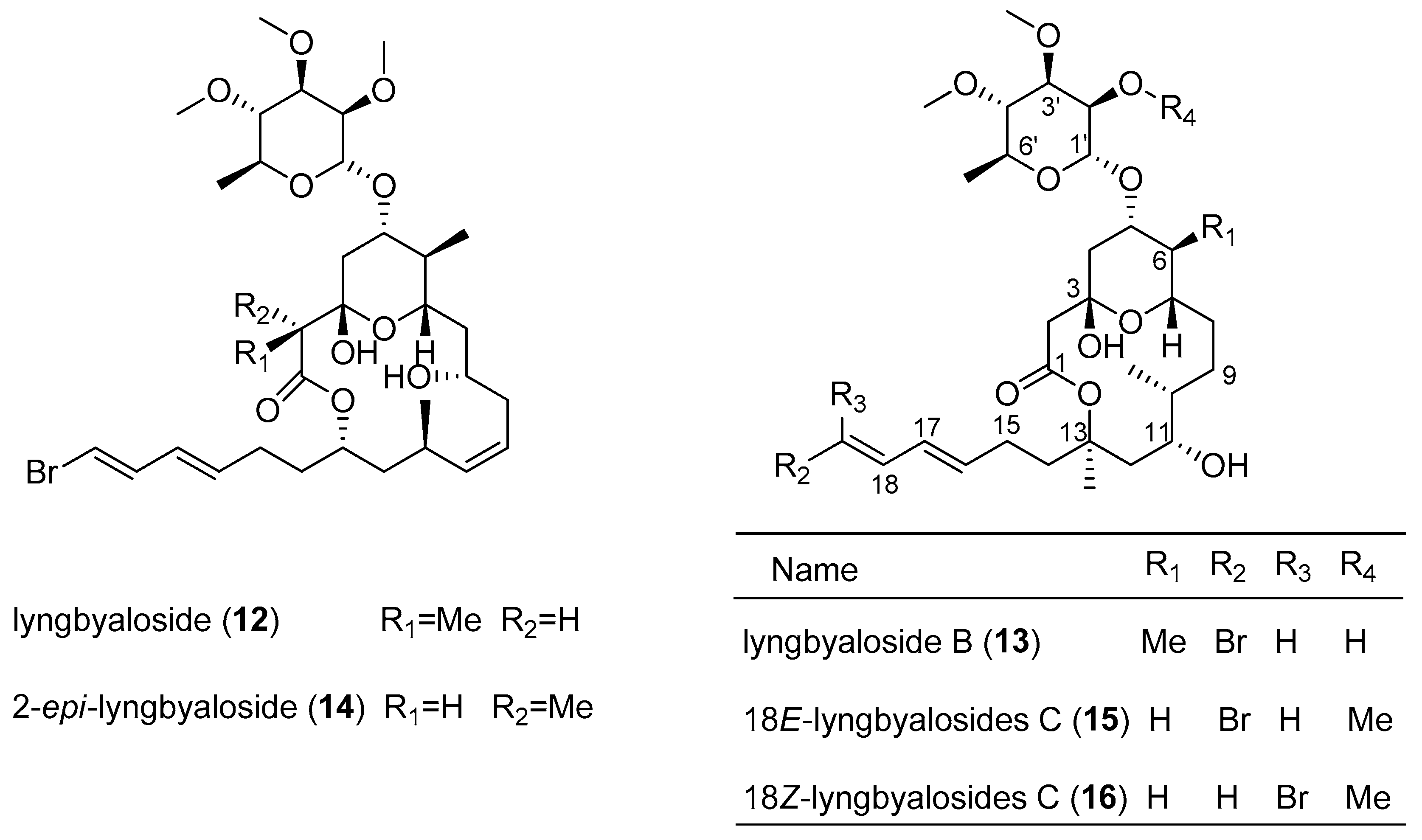
- Klein, D.; Braekman, J.C.; Daloze, D.; Hoffmann, L.; Demoulin, V. Lyngbyaloside, a novel 2,3,4-Tri-O-methyl-6-deoxy-r-mannopyranoside macrolide from Lyngbya bouillonii (Cyanobacteria). J. Nat. Prod. 1997, 60, 1057–1059. [Google Scholar] [CrossRef]
- DeVita, V.T.J.; Rosenberg, S.A. Two hundred years of cancer research. N. Engl. J. Med. 2012, 366, 2207–2214. [Google Scholar] [CrossRef] [PubMed]
- Luesch, H.; Yoshida, W.Y.; Moore, R.E.; Paul, V.J. Isolation and structure of the cytotoxin lyngbyabellin B and absolute configuration of lyngbyapeptin A from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2000, 63, 1437–1439. [Google Scholar] [CrossRef] [PubMed]
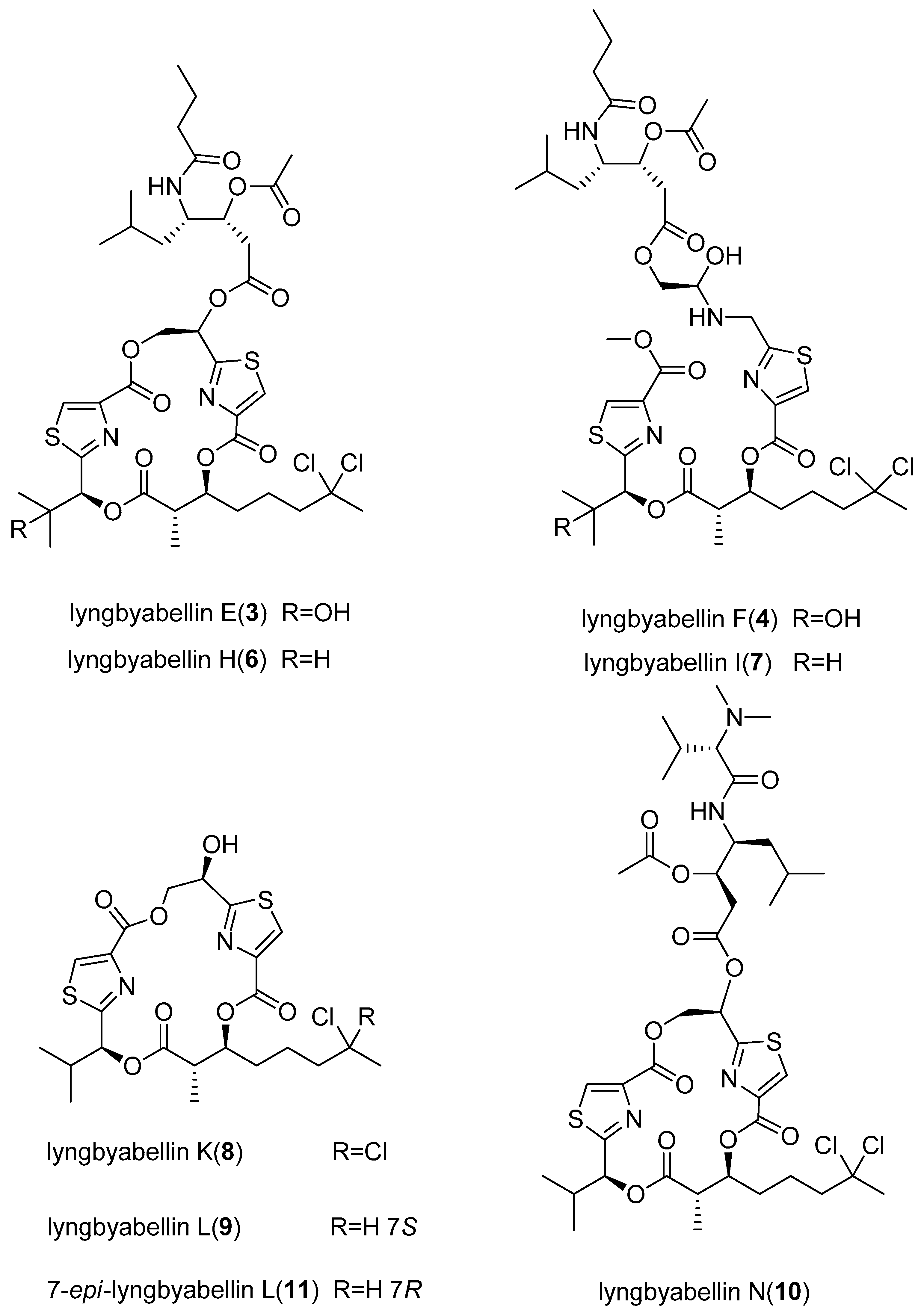
- Han, B.N.; McPhail, K.L.; Gross, H.; Goeger, D.E.; Mooberry, S.L.; Gerwick, W.H. Isolation and structure of five lyngbyabellin derivatives from a Papua New Guinea collection of the marine cyanobacterium Lyngbya majuscula. Tetrahedron 2005, 61, 11723–11729. [Google Scholar] [CrossRef]
- Choi, H.; Mevers, E.; Byrum, T.; Valeriote, F.A.; Gerwick, W.H. Lyngbyabellins K–N from Two Palmyra Atoll Collections of the marine cyanobacterium Moorea bouillonii. Eur. J. Org. Chem. 2012, 5141–5150. [Google Scholar] [CrossRef] [PubMed]
- Luesch, H.; Yoshida, W.Y.; Harrigan, G.G.; Doom, J.P.; Moore, R.E.; Paul, V.J. Lyngbyaloside B, a new glycoside macrolide from a palauan marine cyanobacterium, Lyngbya sp. J. Nat. Prod. 2002, 65, 1945–1948. [Google Scholar] [CrossRef] [PubMed]
- Matthew, S.; Salvador, L.A.; Schupp, P.J.; Paul, V.J.; Luesch, H. Cytotoxic halogenated macrolides and modified peptides from the apratoxin-producing marine cyanobacterium Lyngbya bouillonii from Guam. J. Nat. Prod. 2010, 73, 1544–1552. [Google Scholar] [CrossRef] [PubMed]
- Teruya, T.; Sasaki, H.; Kitamura, K.; Nakayama, T.; Suenaga, K. Biselyngbyaside, a macrolide glycoside from marine cyanobacterium Lyngbya sp. Org. Lett. 2009, 11, 2421–2424. [Google Scholar] [CrossRef] [PubMed]
- Morita, M.; Ohno, O.; Teruya, T.; Yamori, T.; Inuzuka, T.; Suenaga, K. Isolation and structures of biselyngbyasides B, C, and D from the marine cyanobacterium Lyngbya sp., and the biological activities of biselyngbyasides. Tetrahedron 2012, 68, 5984–5990. [Google Scholar] [CrossRef]
- Watanabe, A.; Ohno, O.; Morita, M.; Inuzuka, T.; Suenaga, K. Structures and biological activities of novel biselyngbyaside analogs isolated from the marine cyanobacterium Lyngbya sp. Bull. Chem. Soc. Jpn. 2015, 88, 1256–1264. [Google Scholar] [CrossRef]
- Morita, M.; Ohno, O.; Suenaga, K. Biselyngbyolide A, a novel cytotoxic macrolide from the marine cyanobacterium Lyngbya sp. Chem. Lett. 2012, 41, 165–167. [Google Scholar] [CrossRef]
- Ohno, O.; Watanabe, A.; Morita, M.; Suenaga, K. Biselyngbyolide B, a novel ER stress-inducer isolated from the marine cyanobacterium Lyngbya sp. Chem. Lett. 2014, 43, 287–289. [Google Scholar] [CrossRef]
- MacMillan, J.B.; Molinski, T.F. Caylobolide A, a unique 36-membered macrolactone from a Bahamian Lyngbya majuscula. Org. Lett. 2002, 4, 1535–1538. [Google Scholar] [CrossRef] [PubMed]
- Salvador, L.A.; Paul, V.J.; Luesch, H. Caylobolide B, a macrolactone from symplostatin 1-producing marine cyanobacteria phormidium spp. from Florida. J. Nat. Prod. 2010, 73, 1606–1609. [Google Scholar] [CrossRef] [PubMed]
- Andrianasolo, E.H.; Gross, H.; Goeger, D.; Musafija-Girt, M.; McPhail, K.; Leal, R.M.; Mooberry, S.L.; Gerwick, W.H. Isolation of swinholide A and related glycosylated derivatives from two field collections of marine cyanobacteria. Org. Lett. 2005, 7, 1375–1378. [Google Scholar] [CrossRef] [PubMed]
- Carmeli, S.; Moore, R.E.; Patterson, G.M.L. Tolytoxin and new scytophycins from three species of Scytonema. J. Nat. Prod. 1990, 53, 1533–1542. [Google Scholar] [CrossRef] [PubMed]
- Patterson, G.M.L.; Carmeli, S. Biological effects of tolytoxin (6-hydroxy-7-O-methyl-scytophycin b), a potent bioactive metabolite from cyanobacteria. Arch. Microbiol. 1992, 157, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Mynderse, J.S.; Moore, R.E.; Kashiwagi, M.; Norton, T.R. Antileukemia activity in the Osciliatoriaceae: Isolation of debromoaplysiatoxin from Lyngbya. Science 1977, 196, 538–540. [Google Scholar] [CrossRef] [PubMed]
- Tan, L.T.; Márquez, B.L.; Gerwick, W.H. Lyngbouilloside, a novel glycosidic macrolide from the marine cyanobacterium Lyngbya bouillonii. J. Nat. Prod. 2002, 65, 925–928. [Google Scholar] [CrossRef] [PubMed]
- Iwasaki, A.; Teruya, T.; Suenaga, K. Isolation and structure of koshikalide, a 14-membered macrolide from the marine cyanobacterium Lyngbya sp. Tetrahedron Lett. 2010, 51, 959–960. [Google Scholar] [CrossRef]
- Kang, H.S.; Krunic, A.; Orjala, J. Sanctolide A, a 14-membered PK-NRP hybrid macrolide from the cultured cyanobacterium Oscillatoria sancta (SAG 74.79). Tetrahedron Lett. 2012, 53, 3563–3567. [Google Scholar] [CrossRef] [PubMed]
- Barchi, J.J.; Moore, R.E.; Patterson, G.M.L. Acutiphycin and 20,21-didehydroacutiphycin, new antineoplastic agents from the cyanophyte Oscillatoria acutissima. J. Am. Chem. Soc. 1984, 106, 8193–8197. [Google Scholar] [CrossRef]
- Navarro, G.; Cummings, S.; Lee, J.; Moss, N.; Glukhov, E.; Valeriote, F.A.; Gerwick, L.; Gerwick, W.H. Isolation of polycavernoside D from a marine cyanobacterium. Environ. Sci. Technol. Lett. 2015, 2, 166–170. [Google Scholar] [CrossRef]
- Shao, C.L.; Linington, R.G.; Balunas, M.J.; Centeno, A.; Boudreau, P.; Zhang, C.; Engene, N.; Spadafora, C.; Mutka, T.S.; Kyle, D.E.; Gerwick, L.; Wang, C.Y.; Gerwick, W.H. Bastimolide A, a potent antimalarial polyhydroxy macrolide from the marine cyanobacterium Okeania hirsute. J. Org. Chem. 2015, 80, 7849–7855. [Google Scholar] [CrossRef] [PubMed]
- Mori, S.; Williams, H.; Cagle, D.; Karanovich, K.F.; Horgen, D.; Smith, R.; Watanabe, C.M.H. Macrolactone nuiapolide, isolated from a Hawaiian marine cyanobacterium, exhibits anti-chemotactic activity. Mar. Drugs 2015, 13, 6274–6290. [Google Scholar] [CrossRef] [PubMed]
- Fuwa, H.; Yamagata, N.Y.; Okuaki, Y.T.; Ogata, Y.Y.; Saito, A.; Sasaki, M. total synthesis and complete stereostructure of a marine macrolide glycoside, (-)-lyngbyaloside B. Chem. Eur. J. 2016, 22, 6815–6829. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.; Stefan, E.; Taylo, R.E. Total synthesis and structural reassignment of lyngbyaloside C highlighted by intermolecular ketene esterification. Chem. Eur. J. 2015, 21, 10681–10686. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekhar, S.; Rajesh, G.; Naresh, T. Enantioselective synthesis of the C5–C23 segment of biselyngbyaside. Tetrahedron Lett. 2013, 54, 252–255. [Google Scholar] [CrossRef]
- Yadav, J.S.; Swapnil, N.; Venkatesh, M.; Prasad, A.R. Studies directed toward the synthesis of caylobolide A: convergent synthesis of C21–C40 subunit. Tetrahedron Lett. 2014, 55, 1164–1167. [Google Scholar] [CrossRef]
- Ishibashi, M.; Moore, R.E.; Patterson, G.M.L. Scytophycins, cytotoxic and antimycotic agents from the cyanophyte Scytonema pseudohofmanni. J. Org. Chem. 1986, 51, 5300–5306. [Google Scholar] [CrossRef]
- Cui, J.; Watanabe, T.; Shibasaki, M. Catalytic asymmetric synthesis of key intermediate for scytophycin C. Tetrahedron Lett. 2016, 57, 446–448. [Google Scholar] [CrossRef]
- Moore, R.E. Constituents of blue-green algae. In Marine Natural Products; Scheuer, P.J., Ed.; Academic Press: New York, NY, USA, 1981; Volume 4, pp. 1–52. [Google Scholar]
- Solomon, A.E.; Stoughton, R.B. Dermatitis from purified sea algae toxin (debromoaplysiatoxin). Arch. Dermatol. 1978, 114, 1333–1335. [Google Scholar] [CrossRef] [PubMed]
- Mynderse, J.S.; Moore, R.E. Toxins from Blue-Green algae: Structures ofoscillatoxin A and three related bromine-containing toxins. J. Org. Chem. 1978, 43, 2301–2303. [Google Scholar] [CrossRef]
- Kunifuda, K.; Iwasaki, A.; Nagamoto, M.; Suenaga, K. Total synthesis and absolute configuration of koshikalide. Tetrahedron Lett. 2016, 57, 3121–3123. [Google Scholar] [CrossRef]
- Yadav, J.S.; Suresh, B.; Srihari, P. Stereoselective total synthesis of the marine macrolide sanctolide A. Eur. J. Org. Chem. 2015, 5856–5863. [Google Scholar] [CrossRef]
- Frayman, K.; Robinson, P. Macrolide therapy in cystic fibrosis: New developments in clinical use. Clin. Investig. 2013, 3, 1179–1186. [Google Scholar] [CrossRef]
- Shishido, T.K.; Humisto, A.; Jokela, J.; Liu, L.W.; Wahlsten, M.; Tamrakar, A.; Fewer, D.P.; Permi, P.; Andreote, A.P.D.; Fiore, M.F.; Sivonen, K. Antifungal compounds from cyanobacteria. Mar. Drugs 2015, 13, 2124–2140. [Google Scholar] [CrossRef] [PubMed]
- Salvador-Reyes, L.A.; Sneed, J.; Paul, V.J.; Luesch, H. Amantelides A and B, polyhydroxylated macrolides with differential broad-spectrum cytotoxicity from a guamanian marine cyanobacterium. J. Nat. Prod. 2015, 78, 1957–1962. [Google Scholar] [CrossRef] [PubMed]
- Wood, R. Acute animal and human poisonings from cyanotoxin exposure—A review of the literature. Environ. Int. 2016, 9, 276–282. [Google Scholar] [CrossRef] [PubMed]
- Stierle, D.B.; Stierle, A.A.; Bugni, T.; Loewen, G. Gloeolactone, a new epoxy lactone from a blue-green alga. J. Nat. Prod. 1998, 61, 251–252. [Google Scholar] [CrossRef] [PubMed]
- Williamson, R.T.; Boulanger, A.; Vulpanovici, A.; Roberts, M.A.; Gerwick, W.H. Structure and absolute stereochemistry of phormidolide, a new toxic metabolite from the marine cyanobacterium phormidium sp. J. Org. Chem. 2002, 67, 7927–7936. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.R.; McCue, C.F.; Gerwick, W.H. Cyanolide A, a glycosidic macrolide with potent molluscicidal activity from the Papua New Guinea cyanobacterium Lyngbya bouillonii. J. Nat. Prod. 2010, 73, 217–220. [Google Scholar] [CrossRef] [PubMed]
- Bates, R.W.; Lek, T.G. A synthesis of cyanolide A by intramolecular oxa-michael addition. Synthesis 2014, 46, 1731–1738. [Google Scholar] [CrossRef]
- Yonezawa, T.; Mase, N.; Sasaki, H.; Teruya, T.; Hasegawa, S.; Cha, B.Y.; Yagasaki, K.; Suenaga, K.; Nagai, K.; Woo, J.T. Biselyngbyaside, isolated from marine cyanobacteria, inhibits osteoclastogenesis and induces apoptosis in mature osteoclasts. J. Cell. Biochem. 2012, 113, 440–448. [Google Scholar] [CrossRef] [PubMed]
- Gutiérrez, M.; Tidgewell, K.; Capson, T.L.; Engene, N.; Almanza, A.; Schemies, J.; Jung, M.; Gerwick, W.H. Malyngolide Dimer, a bioactive symmetric cyclodepside from the panamanian marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 2010, 73, 709–711. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, M.; Andrianasolo, E.H.; Shin, W.K.; Goeger, D.E.; Yokochi, A.; Schemies, J.; Jung, M.; France, D.; Cornell-Kennon, S.; Lee, E. Structural and synthetic investigations of tanikolide dimer, a SIRT2 selective inhibitor, and tanikolideseco-acid from the Madagascar marine cyanobacterium Lyngbya majuscula. J. Org. Chem. 2009, 74, 5267–5275. [Google Scholar] [CrossRef] [PubMed]
- Pereira, A.R.; Cao, Z.Y.; Engene, N.; Soria-Mercado, I.E.; Murray, T.F.; Gerwick, W.H. Palmyrolide A, an Unusually Stabilized Neuroactive Macrolide from Palmyra Atoll Cyanobacteria. Org. Lett. 2010, 12, 4490–4493. [Google Scholar] [CrossRef] [PubMed]
- Sudhakar, G.; Reddy, K.J.; Nanubolu, J.B. Total synthesis of palmyrolide A and its 5,7-epi isomers. Org. Lett. 2010, 12, 4490–4493. [Google Scholar] [CrossRef]
- Klein, D.; Braekman, J.C.; Daloze, D. Laingolide, a novel 15-membered macrolide from Lyngbya bouillonii (cyanophyceae). Tetrahedron Lett. 1996, 37, 7519–7520. [Google Scholar] [CrossRef]
- Klein, D.; Braekman, J.C.; Daloze, D.; Hoffmann, L.; Castillo, G.; Demoulin, V. Madangolide and laingolide A, two novel macrolides from Lyngbya bouillonii (Cyanobacteria). J. Nat. Prod. 1999, 62, 934–936. [Google Scholar] [CrossRef] [PubMed]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolite | Source | Location | Target Cell Lines | Concentration/Effect | Reference |
|---|---|---|---|---|---|
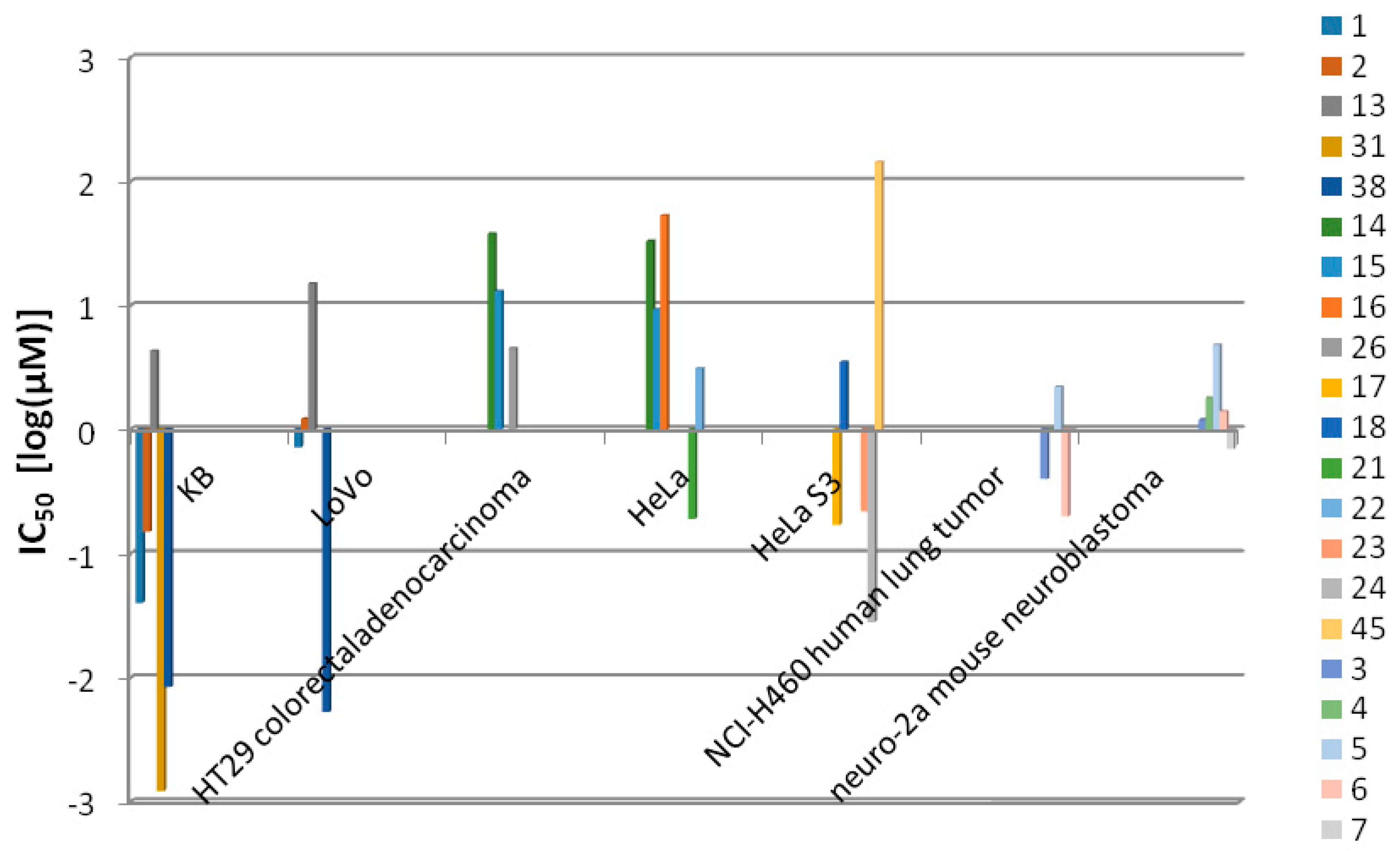
| lyngbyabellin A (1) | Lyngbya majuscula | Guam | KB cells and LoVo cells | IC50 value of 0.03 and 0.50 µg/mL respectively | [6] |
| lyngbyabellin B (2) | Lyngbya majuscula | Guam | KB cells and LoVo cells | IC50 value of 0.10 and 0.83 µg/mL respectively | [10] |
| lyngbyabellin E (3) | Lyngbya majuscula | Papua New Guinea | NCI-H460 human lung tumor and neuro-2a mouse neuroblastoma cells | LC50 value of 0.4 and 1.2 µM respectively | [11] |
| lyngbyabellin F (4) | Lyngbya majuscula | Papua New Guinea | NCI-H460 human lung tumor and neuro-2a mouse neuroblastoma cells | LC50 value of 1.0 and 1.8 µM respectively | [11] |
| lyngbyabellin G (5) | Lyngbya majuscula | Papua New Guinea | NCI-H460 human lung tumor and neuro-2a mouse neuroblastoma cells | LC50 value of 2.2 and 4.8 µM respectively | [11] |
| lyngbyabellin H (6) | Lyngbya majuscula | Papua New Guinea | NCI-H460 human lung tumor and neuro-2a mouse neuroblastoma cells | LC50 value of 0.2 and 1.4 µM respectively | [11] |
| lyngbyabellin I (7) | Lyngbya majuscula | Papua New Guinea | NCI-H460 human lung tumor and neuro-2a mouse neuroblastoma cells | LC50 value of 1.0 and 0.7 µM respectively | [11] |
| lyngbyabellin N (10) | Moorea bouillonii | Palmyra Atoll, USA | H-460 human lung carcinoma and HCT116 colon cancer cell lines | IC50 value of 0.0048–1.8 µM and 15 µM respectively | [12] |
| lyngbyaloside B (13) | Lyngbya sp. | Palau | KB cells and LoVo cells | IC50 value of 4.3 and 15 µM respectively | [13] |
| 2-epi-lyngbyalosid (14) | Lyngbya bouillonii | Apra Harbor, Guam | HT29 colorectal adenocarcinoma and HeLa cells | IC50 value of 38 and 33 µM respectively | [14] |
| 18E-lyngbyaloside C (15) | Lyngbya bouillonii | Apra Harbor, Guam | HT29 colorectal adenocarcinoma and HeLa cells | IC50 value of 13 and 9.3 µM respectively | [14] |
| 18Z-lyngbyaloside C (16) | Lyngbya bouillonii | Apra Harbor, Guam | HT29 colorectal adenocarcinoma and HeLa cells | IC50 value of >100 µM and 53 µM respectively | [14] |
| biselyngbyaside (17) | Lyngbya sp. | Tokunoshima Island, Japan | HeLa S3 cells | IC50 value of 0.1 μg/mL | [15] |
| biselyngbyaside B (18) | Lyngbya sp. | Tokunoshima Island, Japan | HeLa S3 cells and HL60 cells | IC50 value of 3.5 and 0.82 µM respectively | [16] |
| biselyngbyaside E (21) | Lyngbya sp. | Ishigaki Island, Japan | HeLa and HL60 cells | IC50 value of 0.19 and 0.071 µM respectively | [17] |
| biselyngbyaside F (22) | Lyngbya sp. | Ishigaki Island, Japan | HeLa and HL60 cells | IC50 value of 3.1 and 0.66 µM respectively | [17] |
| biselyngbyolide A (23) | Lyngbya sp. | Tokunoshima Island, Japan | HeLa S3 cells and HL60 cells | IC50 value of 0.22 and 0.027 µM respectively | [18] |
| biselyngbyolide B (24) | Lyngbya sp. | Ishigaki Island, Japan | HeLa S3 cells and HL60 cells | IC50 value of 0.028 and 0.0027 µM respectively | [19] |
| caylobolide A (25) | Lyngbya majuscula | Bahamian | human colon tumor cells HCT 116 | IC50 values of 9.9 µM | [20] |
| caylobolide B (26) | Phormidium spp. | Florida USA | HT29 colorectal adenocarcinoma and HeLa cervical carcinoma cells | IC50 value of 4.5 and 12.2 µM respectively | [21] |
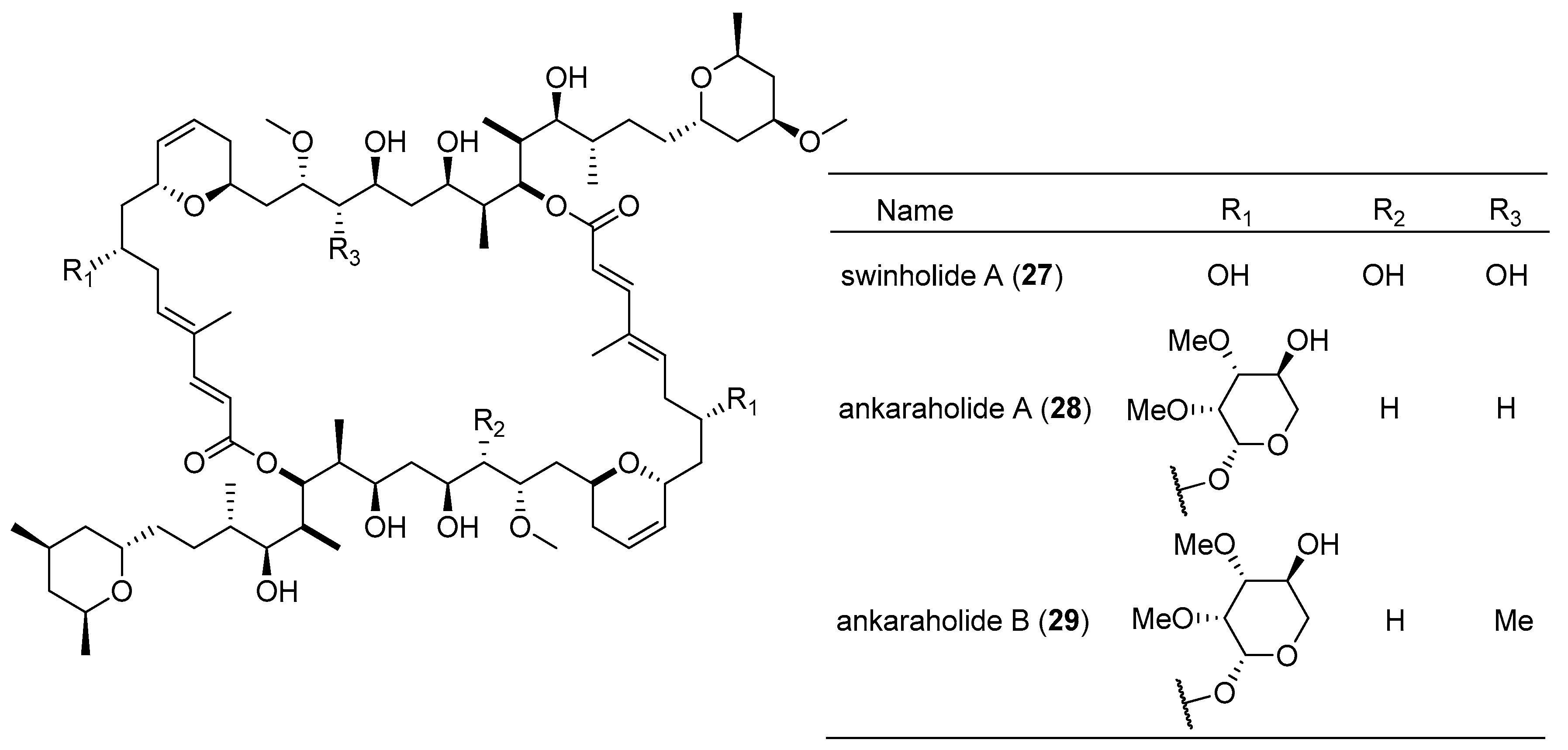
| swinholide A (27) | Symploca cf. sp. | Fiji | several cancer cell lines | IC50 values of 0.37 nM–1.0 µM | [22] |
| ankaraholide A (28) | Geitlerinema sp. | Madagascar | NCI-H460, Neuro-2a cells and MDA-MB-435 cells | IC50 value of 119, 262 and 8.9 nM respectively | [22] |
| scytophycin A (30) | Scytonema pseudohofmanni | Oahu, Hawaii | human carcinoma of nasopharynx Cell (KB cells) | IC50 value of 1 ng/mL | [23] |
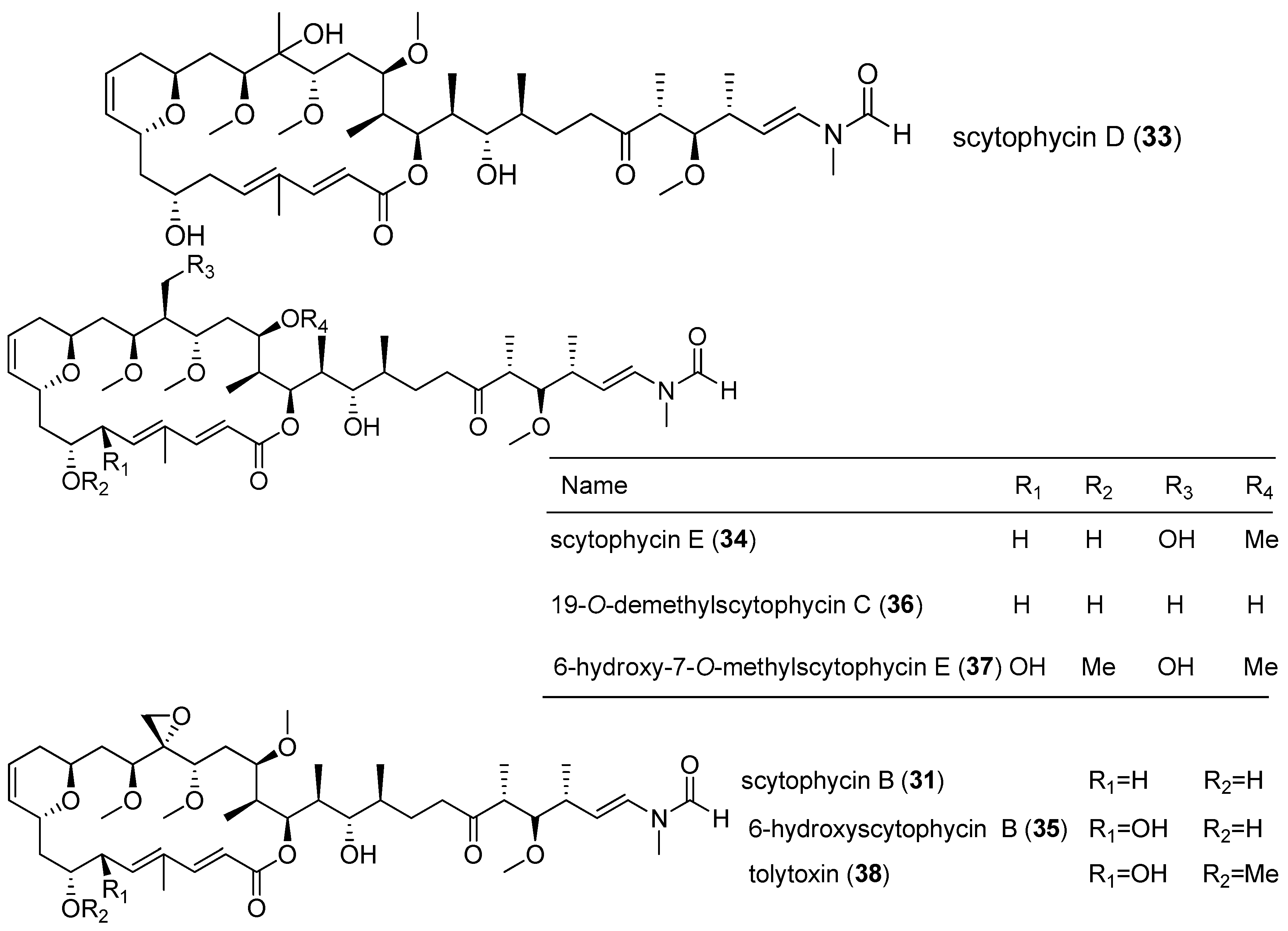
| scytophycin B (31) | Scytonema pseudohofmanni | Oahu, Hawaii | KB cells | IC50 value of 1 ng/mL | [23] |
| scytophycins C-E (32–34) | Scytonema pseudohofmanni | Oahu, Hawaii | KB cells | IC50 value of 10–100 ng/mL | [23] |
| 6-hydroxyscytophycin B (35) | Scytonema mirabile | cultured | KB cells and LoVo cells | MICs of 1–5 and 10–50 ng/mL respectively | [23] |
| 19-O-demethylscytophycin C (36) | Scytonema burmanicurn | cultured | KB cells and LoVo cells | MICs of 1–5 and 10–50 ng/mL respectively | [23] |
| 6-hydroxy-7-O-methylscytophycin E (37) | Scytonema ocellatum | cultured | KB cells and LoVo cells | MICs of 1–5 and 10–50 ng/mL respectively | [23] |
| tolytoxin (38) | Tolypothrix conglutinata var. colorata | Fanning Island | KB cells and LoVo cells | IC50 value of 8.4 and 5.3 nM respectively | [24] |
| debromoaplysiatoxin (39) | Lyngbya majuscula | Marshall Islands | P-388 lymphocytic mouse leukemia | weak | [25] |
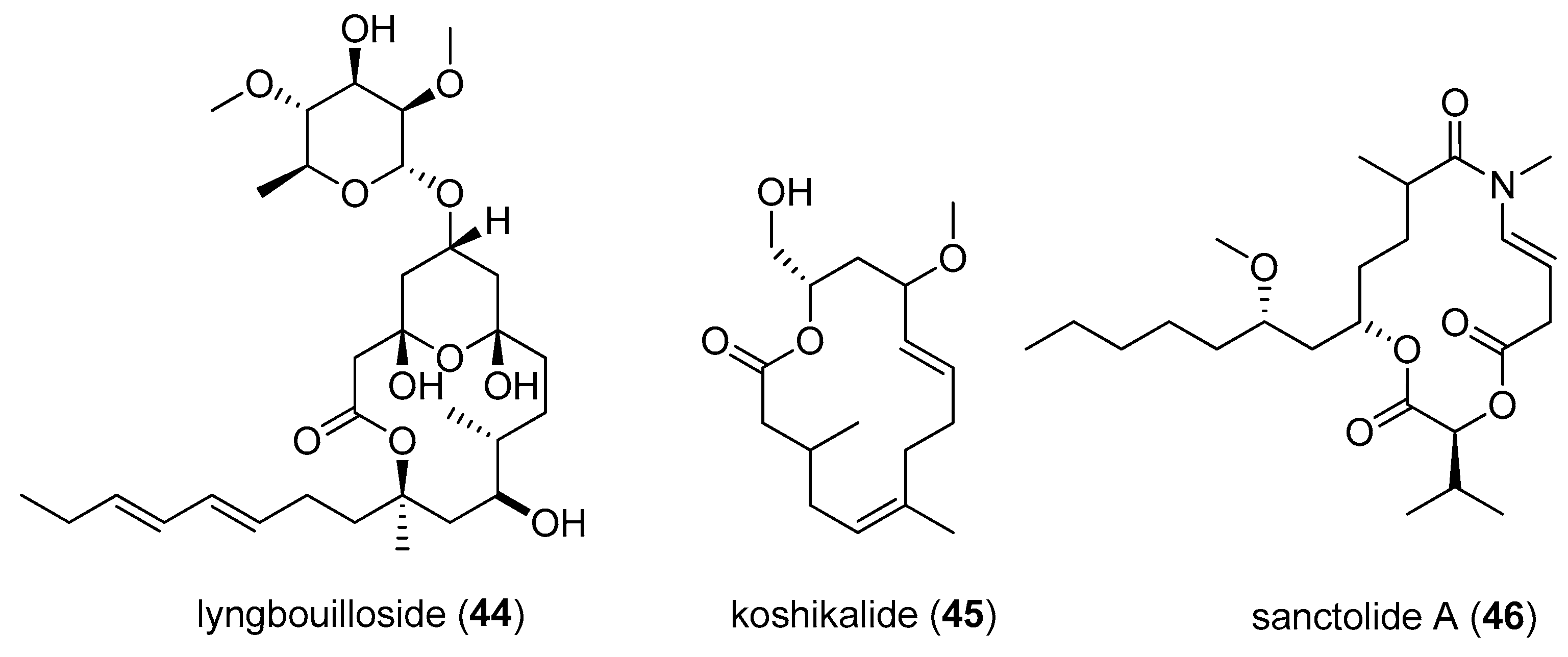
| lyngbouilloside (44) | Lyngbya bouillonii | Papua New Guinea | neuroblastoma cells | IC50 value of 17 µM | [26] |
| koshikalide (45) | Lyngbya sp. | Mie Prefecture | HeLa S3 cells | IC50 value of 42 µg/mL | [27] |
| sanctolide A (46) | Oscillatoria sancta | cultured | HT-29 and MDA-MB-435 cell lines | nd a | [28] |
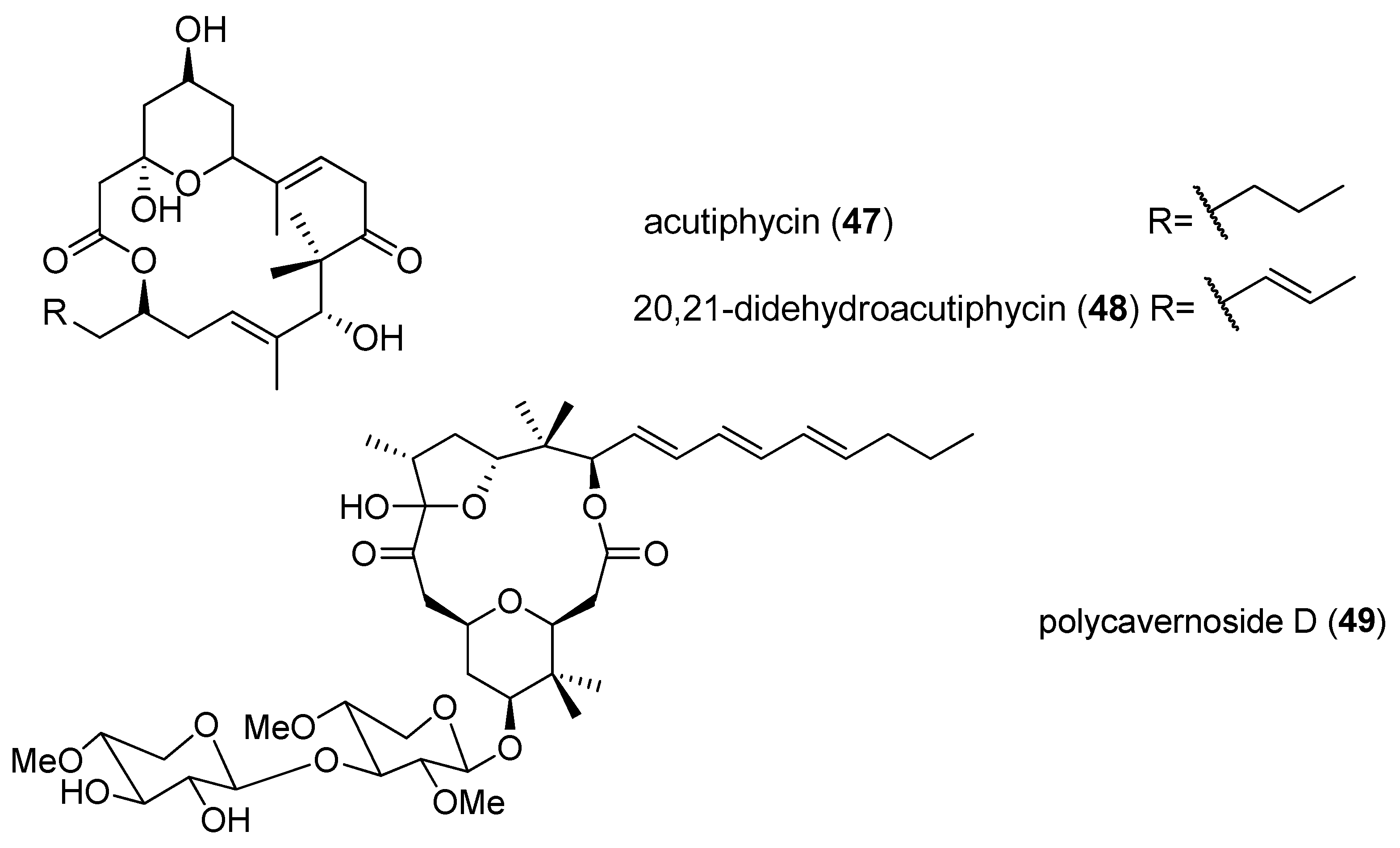
| acutiphycin (47) | Oscillatoria acutissima | Manoa Valley Oahu, Hawaii | KB cells and NIH/3T3 cells | ED50 < 1 µg/mL | [29] |
| 20,21-didehydroacutiphycin (48) | Oscillatoria acutissima | Manoa Valley Oahu, Hawaii | KB cells and NIH/3T3 cells | ED50 < 1 µg/mL | [29] |
| polycavernoside D (49) | Okeania sp. | Puerto Rican | H-460 human lung cancer cell lines | EC50 value of 2.5 µM | [30] |
| bastimolide A (50) | Okeania hirsuta | Panama | Vero cells | IC50 value of 2.1 µM | [31] |
| nuiapolide (51) | colonial cyanobacterium (071905-NII-01) | Hawaii | Jurkat cells and cancerous T lymphocytes | anti-chemotactic activity | [32] |
| Metabolite | Source | Location | Target | Concentration/Effect | Reference |
|---|---|---|---|---|---|
| 6-hydroxyscytophycin B (35) | Scytonema mirabile | cultured | Fungus (1) Aspergillus oryzae (2) Candida albicans (3) Penicillium notatum (4) Saccharomyces cerevisiae | nd a | [23] |
| 19-O-demethylscytophycin C (36) | Scytonema burmanicurn | cultured | Fungus (1) Aspergillus oryzae (2) Candida albicans (3) Penicillium notatum (4) Saccharomyces cerevisiae | nd a | [23] |
| 6-hydroxy-7-O-methylscytophycin E (37) | Scytonema ocellatum | cultured | Fungus (1) Aspergillus oryzae (2) Candida albicans (3) Penicillium notatum (4) Saccharomyces cerevisiae | nd a | [23] |
| tolytoxin (38) | Tolypothrix conglutinata var. colorata | Fanning Island | Fungi Penicillium notatum and Rhizoctonia solani 1165 | MIC value of 0.25 nM respectively | [24] |
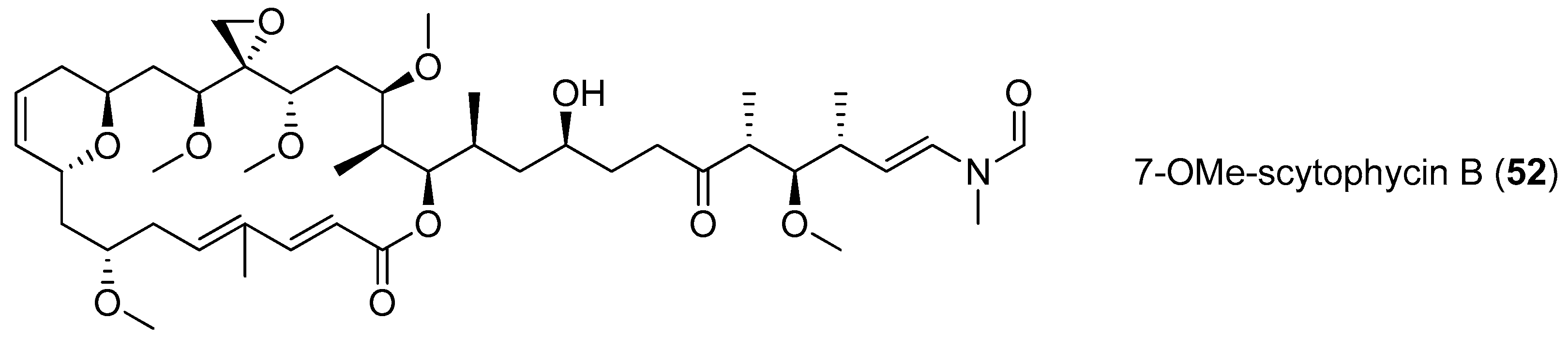
| 7-OMe-scytophycin B (52) | Anabaena sp. | cultured | Fungus Candida albicans HAMBI 484 and Candida guilliermondii HAMBI 257 | MIC values of 0.40 and 0.80 mM respectively; IC50 value of 0.19 and 0.23 µM respectively | [45] |
| amantelide A (53) | Oscillatoriales | Guam | Fungi Lindra thalassiae and Fusarium sp. | totally inhibited of 62.5 μg/mL | [46] |
| amantelide B (54) | Oscillatoriales | Guam | Fungus Dendryphiella salina | totally inhibited of 6.25 μg/mL | [46] |
| Metabolite | Source | Location | Target Fauna | Impacts | Reference |
|---|---|---|---|---|---|
| lyngbyabellin A (1) | Lyngbya majuscula | Guam | mice | LD50 value of 1.2–1.5 mg/kg | [6] |
| tolytoxin (38) | Scytonema pseudohofmanni | cultured | mice | LD50 value of 1.5 mg/kg | [24] |
| sanctolide A (48) | Oscillatoria sancta | cultured | brine shrimp | LD50 value of 23.5 μM | [28] |
| gloeolactone (55) | Gloeotrichia sp. | Clark Canyon Reservoir | brine shrimp | 100% killed at 125 µg/mL | [48] |
| phormidolide (56) | Phormidium sp. | Sulawesi, Indonesia | brine shrimp | LD50 value of 1.5 μM | [49] |
| cyanolide A (57) | Lyngbya bouillonii | Papua New Guinea | snail vector Biomphalaria glabrata | LD50 value of 1.2 μM | [50] |
| Metabolite | Source | Location | Biological Activity | Reference |
|---|---|---|---|---|
| biselyngbyaside (17) | Lyngbya sp. | Okinawa Prefecture Japan | osteoclast differentiation and function | [52] |
| debromoaplysiatoxin (39) | Lyngbya majuscula | Enewetak Atoll, Marshall Islands | produce an irritant pustular folliculitis in humans and cause a severe cutaneous inflammatory reaction in the rabbit and in hairless mice | [25] |
| bastimolide A (50) | Okeania hirsuta | Caribbean coast of Panama | Plasmodium falciparum TM90-C2A, TM90-C2B, W2, TM91-C235 (IC50 values of 80, 90, 140 and 270 nM respectively) | [31] |
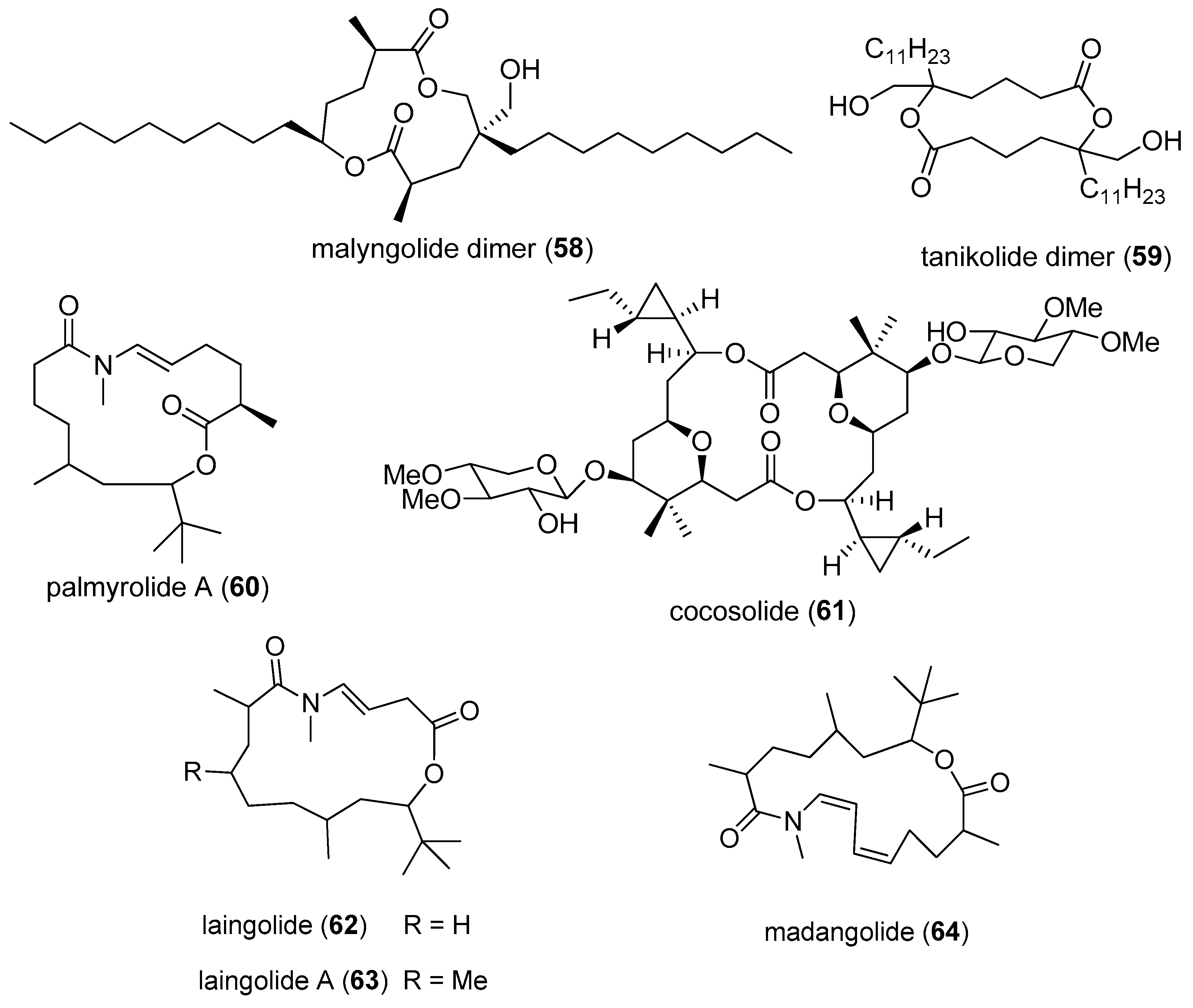
| malyngolide dimer (58) | Lyngbya majuscula | Panama | Plasmodium falciparum (IC50 values of 19 µM) | [53] |
| tanikolide dimer (59) | Lyngbya majuscula | Madagascar | SIRT2 (IC50 = 176 nM to 2.4 µM) | [54] |
| palmyrolide A (60) | Leptolyngbya cf. Oscillatoria sp. | Palmyra Atoll | suppression of calcium influx in cerebrocortical neurons (IC50 value of 3.70 µM) sodium channel blocking activity in neuro-2a cells (IC50 value of 5.2 µM) | [55] |
| cocosolide (61) | Symploca sp. | Guam | inhibition of IL-2 production and T-cell proliferation | [7] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Zhang, J.; He, S.; Yan, X. A Review Study on Macrolides Isolated from Cyanobacteria. Mar. Drugs 2017, 15, 126. https://doi.org/10.3390/md15050126
Wang M, Zhang J, He S, Yan X. A Review Study on Macrolides Isolated from Cyanobacteria. Marine Drugs. 2017; 15(5):126. https://doi.org/10.3390/md15050126
Chicago/Turabian StyleWang, Mengchuan, Jinrong Zhang, Shan He, and Xiaojun Yan. 2017. "A Review Study on Macrolides Isolated from Cyanobacteria" Marine Drugs 15, no. 5: 126. https://doi.org/10.3390/md15050126
APA StyleWang, M., Zhang, J., He, S., & Yan, X. (2017). A Review Study on Macrolides Isolated from Cyanobacteria. Marine Drugs, 15(5), 126. https://doi.org/10.3390/md15050126