The Venom Repertoire of Conus gloriamaris (Chemnitz, 1777), the Glory of the Sea

 ,
,
Abstract
:1. Introduction
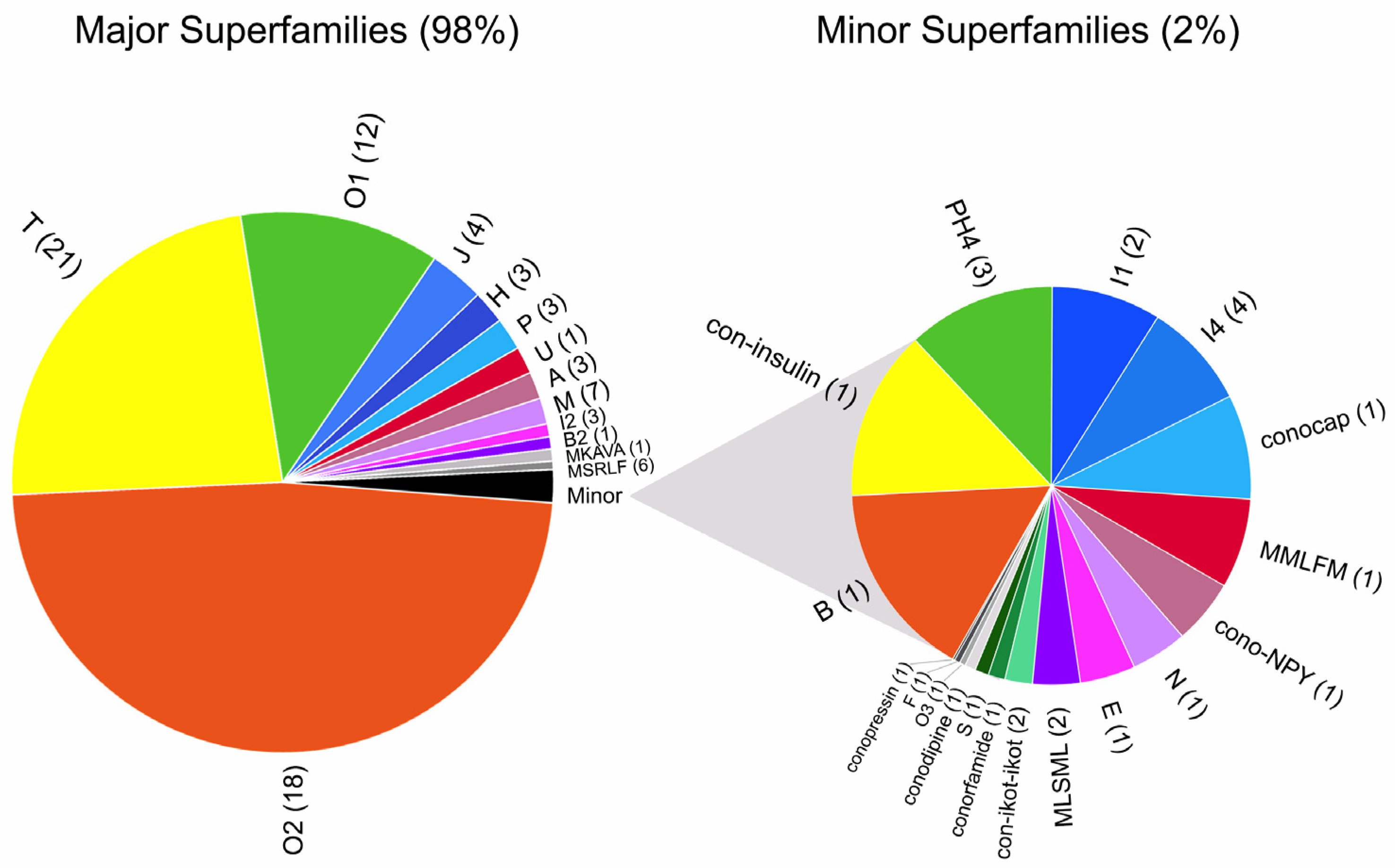
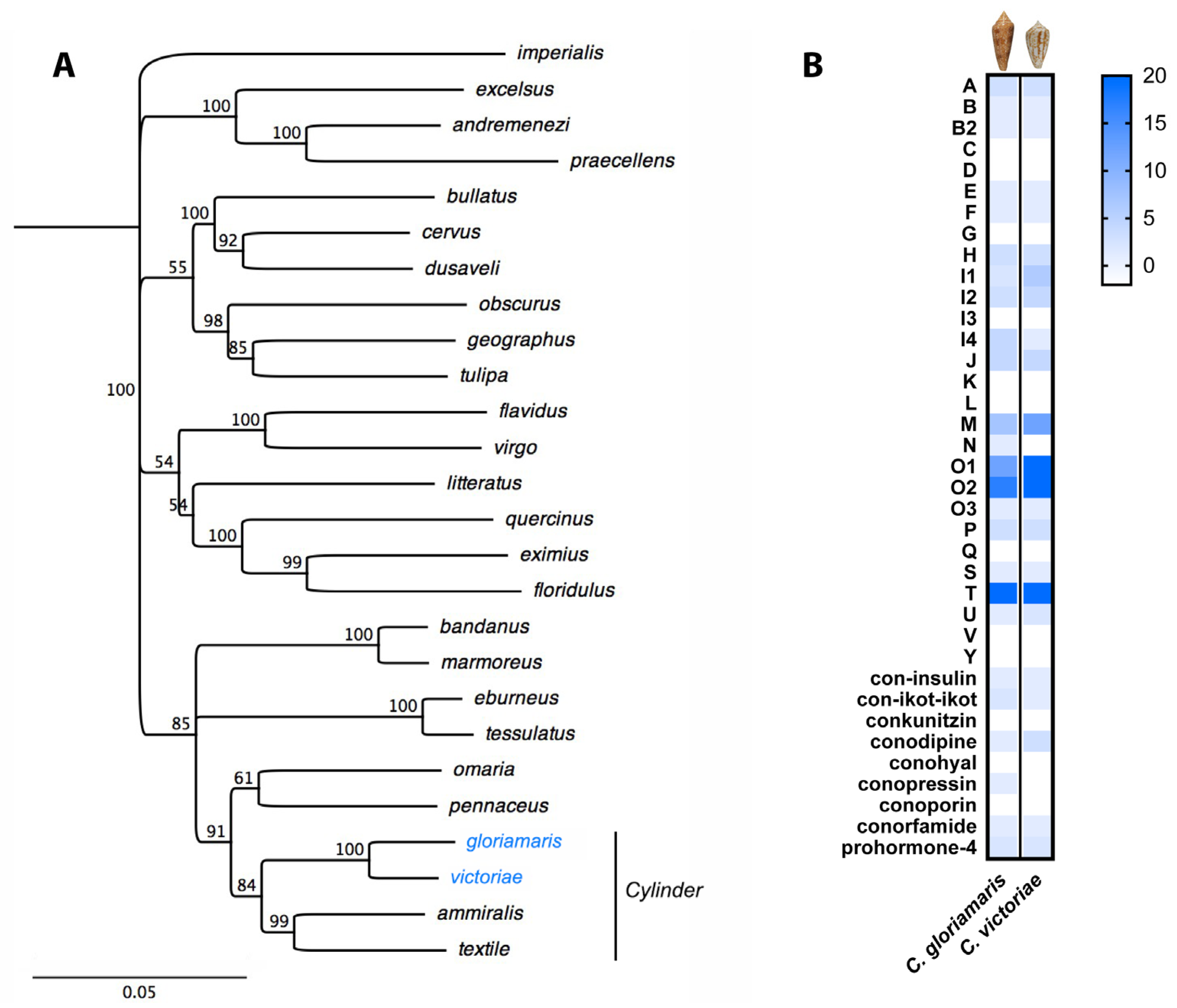
2. Results
2.1. O2-Superfamily
2.2. T-Superfamily
2.3. O1-Superfamily
2.4. J-Superfamily
2.5. H-Superfamily
2.6. P-Superfamily
2.7. U-Superfamily
2.8. A-Superfamily
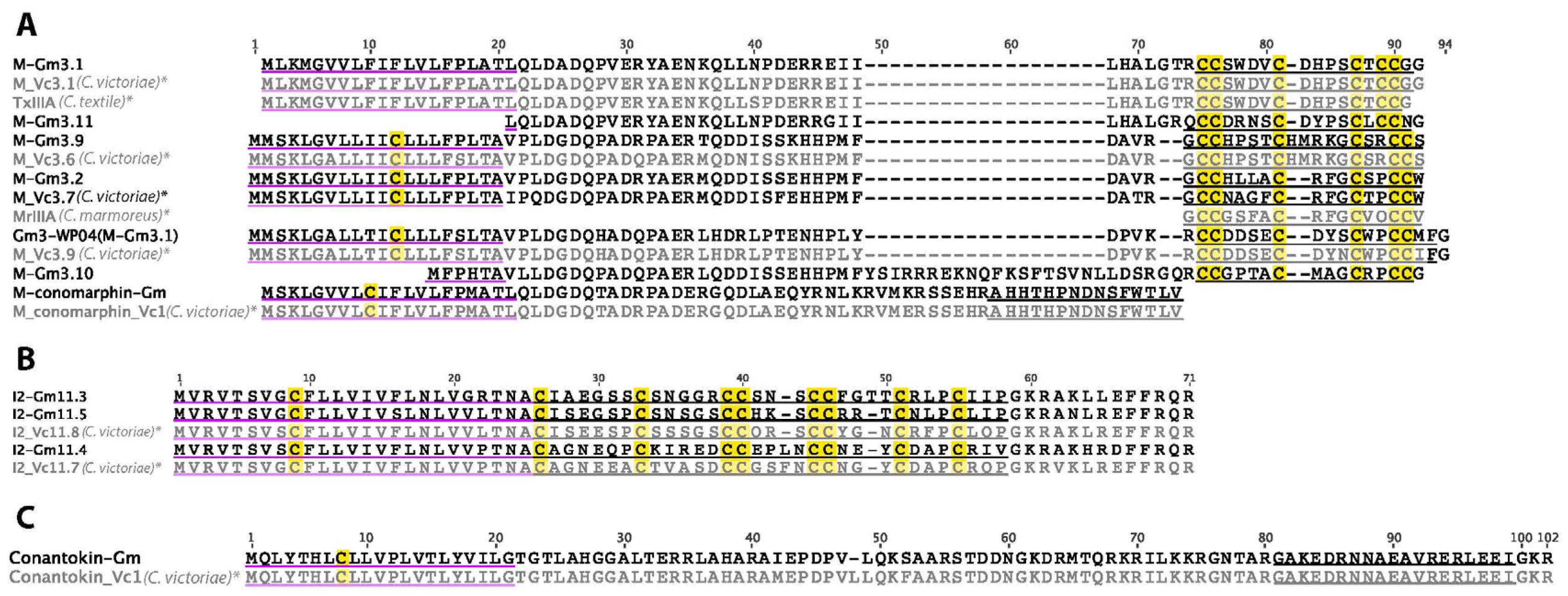
2.9. M-Superfamily
2.10. I2-Superfamily
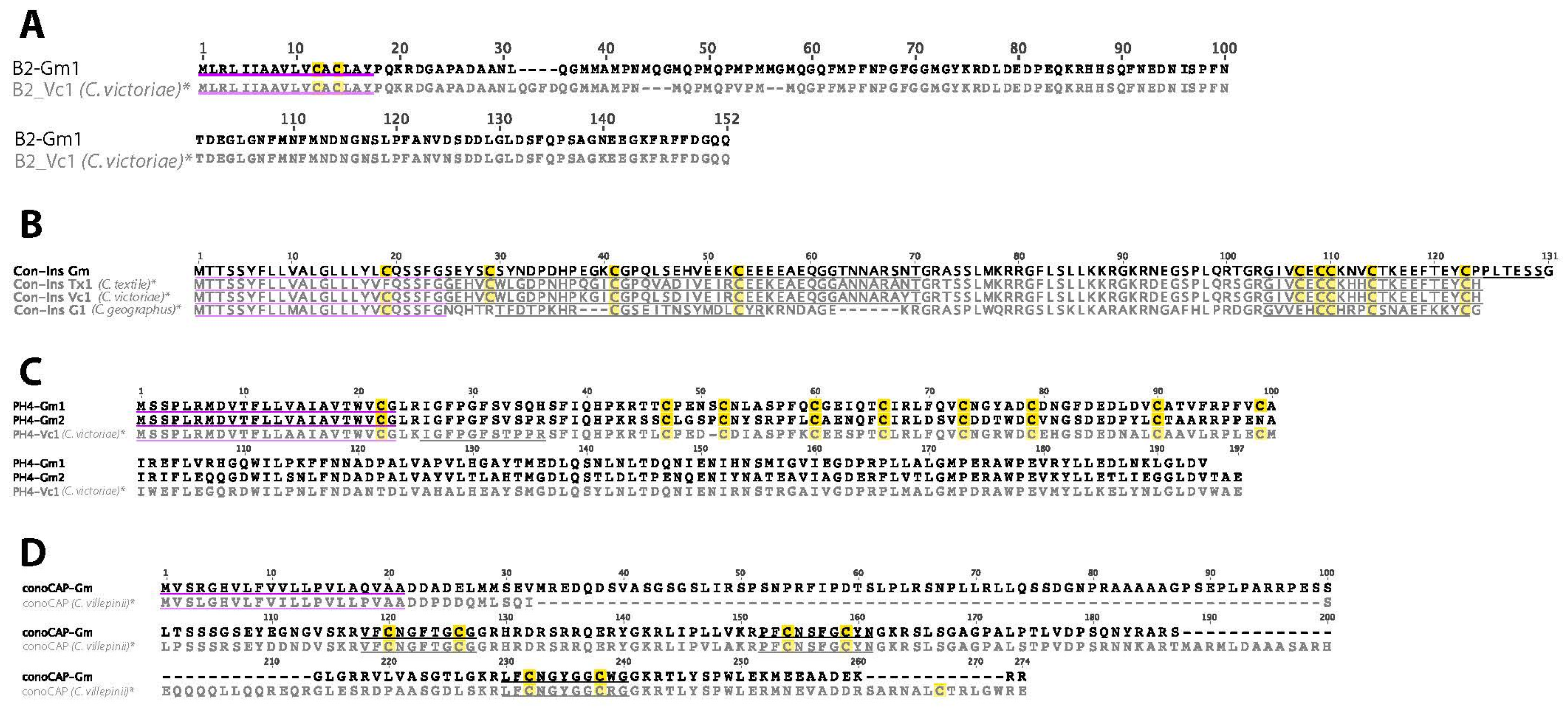
2.11. B2-Superfamily
2.12. B-Superfamily (Conantokins)
2.13. Con-Insulin
2.14. Prohormone-4
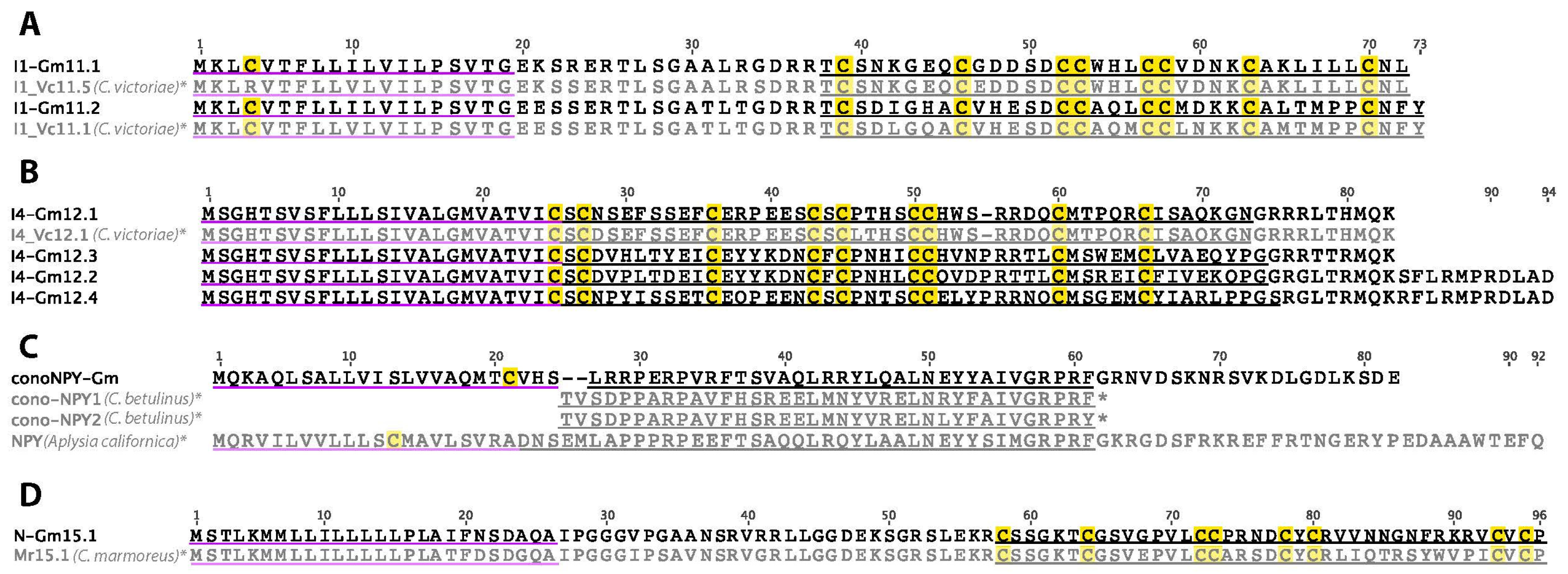
2.15. I1-Superfamily
2.16. I4-Superfamily
2.17. ConoCAP
2.18. Cono-NPY
2.19. N-Superfamily
2.20. E-Superfamily
2.21. Con-ikot-ikot
2.22. Conorfamide
2.23. S-Superfamily
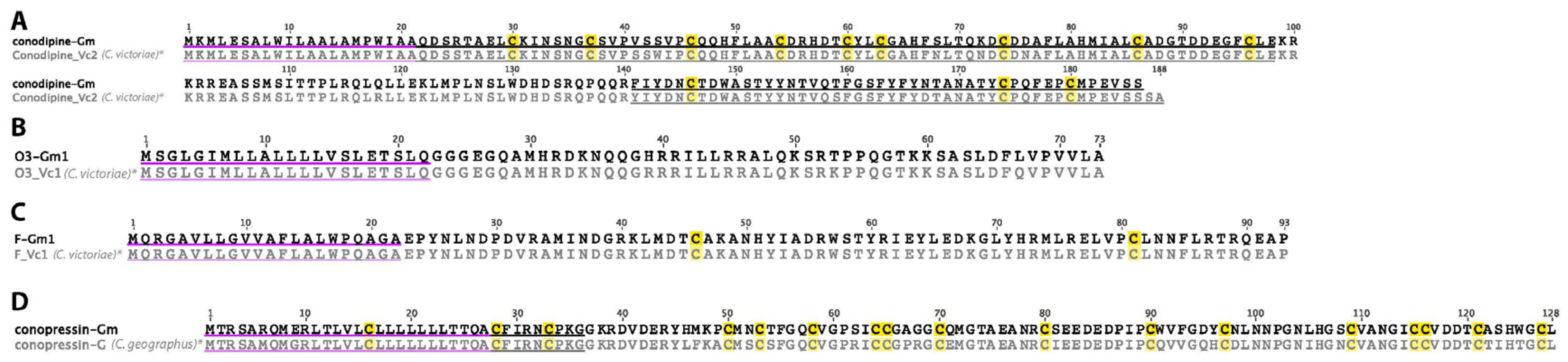
2.24. Conodipine
2.25. O3-Superfamily
2.26. F-Superfamily
2.27. Conopressin
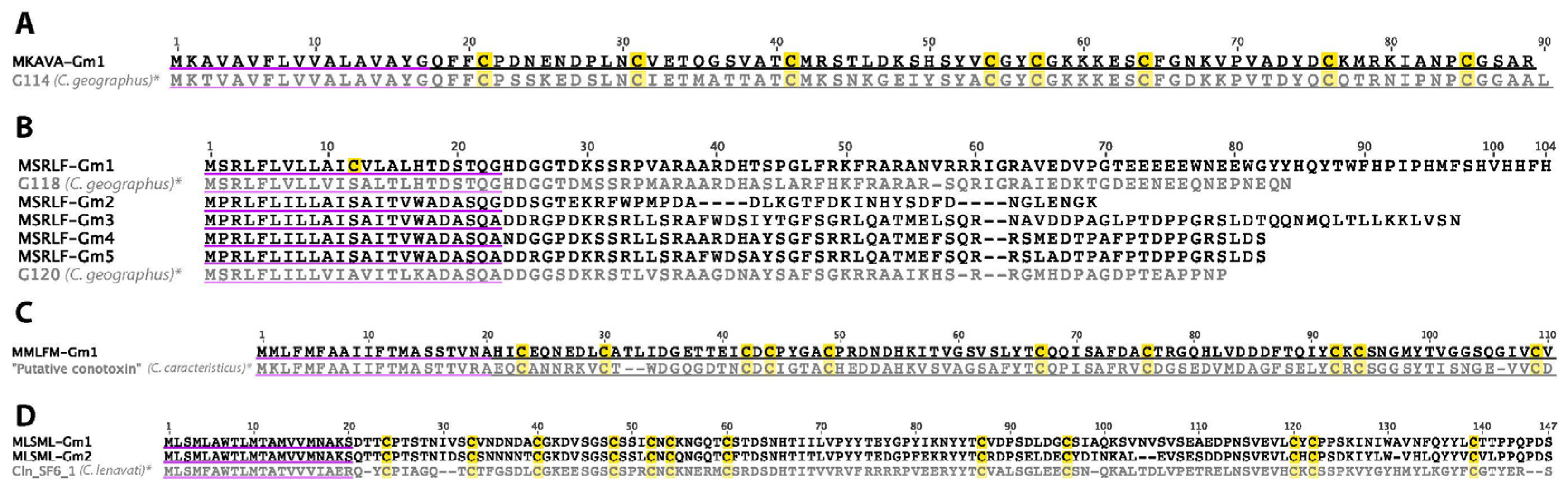
2.28. Putative Conotoxins (MKAVA, MSRLF, MMLFM, MLSML)
3. Discussion
4. Materials and Methods
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Dance, S.P. Rare Shells; Faber and Faber Limited: London, UK, 1969. [Google Scholar]
- Dance, S.P. Shell Collecting: An Illustrated History; University of California Press: Berkeley, California, USA, 1966. [Google Scholar]
- Dance, S.P. A History of Shell Collecting; Brill: Leiden, The Netherlands, 1986. [Google Scholar]
- Röckel, D.; Korn, W.; Kohn, A.J. Manual of the Living Conidae; Verlag Christa Hemmen: Wiesbaden, Germany, 1995. [Google Scholar]
- Olivera, B.M. Conus venom peptides: Correlating chemistry and behavior. J. Comp. Physiol. A Neuroethol. Sens. Neural. Behav. Physiol. 1999, 185, 353–359. [Google Scholar] [CrossRef]
- Terlau, H.; Olivera, B.M. Conus venoms: A rich source of novel ion channel-targeted peptides. Physiol. Rev. 2004, 84, 41–68. [Google Scholar] [CrossRef] [PubMed]
- Miljanich, G.P. Ziconotide: Neuronal calcium channel blocker for treating severe chronic pain. Curr. Med. Chem. 2004, 11, 3029–3040. [Google Scholar] [CrossRef] [PubMed]
- King, G.F. Venoms as a platform for human drugs: translating toxins into therapeutics. Expert Opin. Biol. Ther. 2011, 11, 1469–1484. [Google Scholar] [CrossRef] [PubMed]
- Woodward, S.R.; Cruz, L.J.; Olivera, B.M.; Hillyard, D.R. Constant and hypervariable regions in conotoxin propeptides. EMBO J. 1990, 9, 1015–1020. [Google Scholar] [PubMed]
- Dutertre, S.; Jin, A.-h.; Kaas, Q.; Jones, A.; Alewood, P.F.; Lewis, R.J. Deep venomics reveals the mechanism for expanded peptide diversity in cone snail venom. Mol. Cell. Proteom. 2012, 12, 312–329. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Bandyopadhyay, P.; Olivera, B.; Yandell, M. Elucidation of the molecular envenomation strategy of the cone snail Conus geographus through transcriptome sequencing of its venom duct. BMC Genom. 2012, 13, 284. [Google Scholar] [CrossRef] [PubMed]
- Lluisma, A.O.; Milash, B.A.; Moore, B.; Olivera, B.M.; Bandyopadhyay, P.K. Novel venom peptides from the cone snail Conus pulicarius discovered through next-generation sequencing of its venom duct transcriptome. Mar. Genom. 2012, 5, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.D.; Safavi-Hemami, H.; McIntosh, L.D.; Purcell, A.W.; Norton, R.S.; Papenfuss, A.T. Diversity of conotoxin gene superfamilies in the venomous snail, Conus victoriae. PLoS ONE 2014, 9, e87648. [Google Scholar] [CrossRef] [PubMed]
- Kaas, Q.; Westermann, J.C.; Craik, D.J. Conopeptide characterization and classifications: An analysis using ConoServer. Toxicon 2010, 55, 1491–1509. [Google Scholar] [CrossRef] [PubMed]
- Miles, L.A.; Dy, C.Y.; Nielsen, J.; Barnham, K.J.; Hinds, M.G.; Olivera, B.M.; Bulaj, G.; Norton, R.S. Structure of a novel P-superfamily spasmodic conotoxin reveals an inhibitory cystine knot motif. J. Biol. Chem. 2002, 277, 43033–43040. [Google Scholar] [CrossRef] [PubMed]
- Hasson, A.; Shon, K.J.; Olivera, B.M.; Spira, M.E. Alterations of voltage-activated sodium current by a novel conotoxin from the venom of Conus gloriamaris. J. Neurophysiol. 1995, 73, 1295. [Google Scholar] [PubMed]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De novo transcript sequence reconstruction from RNA-seq using the Trinity platform for reference generation and analysis. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.D.; Norton, R.S. Conotoxin gene superfamilies. Mar. Drugs 2014, 12, 6058–6101. [Google Scholar] [CrossRef] [PubMed]
- Fainzilber, M.; Gordon, D.; Hasson, A.; Spira, M.E.; Zlotkin, E. Mollusc-specific toxins from the venom of Conus textile neovicarius. Eur. J. Biochem. 1991, 202, 589–595. [Google Scholar] [CrossRef] [PubMed]
- Fainzilber, M.; Nakamura, T.; Lodder, J.C.; Zlotkin, E.; Kits, K.S.; Burlingame, A.L. γ-conotoxin-PnVIIA, a γ-carboxyglutamate-containing peptide agonist of neuronal pacemaker cation currents. Biochemistry 1998, 37, 1470–1477. [Google Scholar] [CrossRef] [PubMed]
- Walker, C.S.; Steel, D.; Jacobsen, R.B.; Lirazan, M.B.; Cruz, L.J.; Hooper, D.; Shetty, R.; DelaCruz, R.C.; Nielsen, J.S.; Zhou, L.M.; et al. The T-superfamily of conotoxins. J. Biol. Chem. 1999, 274, 30664–30671. [Google Scholar] [CrossRef] [PubMed]
- Rigby, A.C.; Lucas-Meunier, E.; Kalume, D.E.; Czerwiec, E.; Hambe, B.; Dahlqvist, I.; Fossier, P.; Baux, G.; Roepstorff, P.; Baleja, J.D.; et al. A conotoxin from Conus textile with unusual posttranslational modifications reduces presynaptic Ca2+ influx. Proc. Natl. Acad. Sci. USA 1999, 96, 5758–5763. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wu, Q.; Pi, C.; Zhao, Y.; Zhou, M.; Wang, L.; Chen, S.; Xu, A. Isolation and characterization of a T-superfamily conotoxin from Conus litteratus with targeting tetrodotoxin-sensitive sodium channels. Peptides 2007, 28, 2313–2319. [Google Scholar] [CrossRef] [PubMed]
- Petrel, C.; Hocking, H.G.; Reynaud, M.; Upert, G.; Favreau, P.; Biass, D.; Paolini-Bertrand, M.; Peigneur, S.; Tytgat, J.; Gilles, N.; et al. Identification, structural and pharmacological characterization of τ-CnVA, a conopeptide that selectively interacts with somatostatin sst3 receptor. Biochem. Pharmacol. 2013, 85, 1663–1671. [Google Scholar] [CrossRef] [PubMed]
- Sharpe, I.A.; Gehrmann, J.; Loughnan, M.L.; Thomas, L.; Adams, D.A.; Atkins, A.; Palant, E.; Craik, D.J.; Adams, D.J.; Alewood, P.F.; et al. Two new classes of conopeptides inhibit the α1-adrenoceptor and noradrenaline transporter. Nat. Neurosci. 2001, 4, 902–907. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, J.M.; Corpuz, G.O.; Layer, R.T.; Garrett, J.E.; Wagstaff, J.D.; Bulaj, G.; Vyazovkina, A.; Yoshikami, D.; Cruz, L.J.; Olivera, B.M. Isolation and characterization of a novel Conus peptide with apparent antinociceptive activity. J. Biol. Chem. 2000, 275, 32391–32397. [Google Scholar] [CrossRef] [PubMed]
- Olivera, B.M.; Cartier, G.E.; Watkins, M.; Hillyard, D.R.; McIntosh, J.M.; Layer, R.T.; Jones, R.M. O-Superfamily Conotoxin Peptides. Google Patents US6,762,165 B2, 13 July 2004. [Google Scholar]
- Shon, K.J.; Hasson, A.; Spira, M.E.; Cruz, L.J.; Gray, W.R.; Olivera, B.M. Delta-conotoxin GmVIA, a novel peptide from the venom of Conus gloriamaris. Biochemistry 1994, 33, 11420–11425. [Google Scholar] [CrossRef] [PubMed]
- Imperial, J.S.; Bansal, P.S.; Alewood, P.F.; Daly, N.L.; Craik, D.J.; Sporning, A.; Terlau, H.; López-Vera, E.; Bandyopadhyay, P.K.; Olivera, B.M. A novel conotoxin inhibitor of Kv1.6 channel and nAChR subtypes defines a new superfamily of conotoxins. Biochemistry 2006, 45, 8331–8340. [Google Scholar] [CrossRef] [PubMed]
- Lu, B.-S.; Yu, F.; Zhao, D.; Huang, P.-T.; Huang, C.-F. Conopeptides from Conus striatus and Conus textile by cDNA cloning. Peptides 1999, 20, 1139–1144. [Google Scholar] [PubMed]
- Sandall, D.W.; Satkunanathan, N.; Keays, D.A.; Polidano, M.A.; Liping, X.; Pham, V.; Down, J.G.; Khalil, Z.; Livett, B.G.; Gayler, K.R. A novel α-conotoxin identified by gene sequencing is active in suppressing the vascular response to selective stimulation of sensory nerves in vivo. Biochemistry 2003, 42, 6904–6911. [Google Scholar] [CrossRef] [PubMed]
- Olivera, B.; Rivier, J.; Clark, C.; Ramilo, C.A.; Corpuz, G.P.; Abogadie, F.C.; Edward Mena, E.; Woodward, S.R.; Hillyard, D.R.; Cruz, L.J. Diversity of Conus neuropeptides. Science 1990, 249, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Jacob, R.B.; McDougal, O.M. The M-superfamily of conotoxins: A review. Cell. Mol. Life Sci. 2010, 67, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Conticello, S.G.; Gilad, Y.; Avidan, N.; Ben-Asher, E.; Levy, Z.; Fainzilber, M. Mechanisms for evolving hypervariability: The case of conopeptides. Mol. Biol. Evol. 2001, 18, 120–131. [Google Scholar] [CrossRef] [PubMed]
- Corpuz, G.P.; Jacobsen, R.B.; Jimenez, E.C.; Watkins, M.; Walker, C.; Colledge, C.; Garrett, J.E.; McDougal, O.; Li, W.; Gray, W.R.; et al. Definition of the M-conotoxin superfamily: Characterization of novel peptides from molluscivorous Conus venoms. Biochemistry 2005, 44, 8176–8186. [Google Scholar] [CrossRef] [PubMed]
- Aguilar, M.B.; Pérez-Reyes, L.I.; López, Z.; de la Cotera, E.P.H.; Falcón, A.; Ayala, C.; Galván, M.; Salvador, C.; Escobar, L.I. Peptide sr11a from Conus spurius is a novel peptide blocker for Kv1 potassium channels. Peptides 2010, 31, 1287–1291. [Google Scholar] [CrossRef] [PubMed]
- Fan, C.-X.; Chen, X.-K.; Zhang, C.; Wang, L.-X.; Duan, K.-L.; He, L.-L.; Cao, Y.; Liu, S.-Y.; Zhong, M.-N.; Ulens, C.; et al. A novel conotoxin from Conus betulinus, κ-BtX, unique in cysteine pattern and in function as a specific BK channel modulator. J. Biol. Chem. 2003, 278, 12624–12633. [Google Scholar] [CrossRef] [PubMed]
- Kauferstein, S.; Huys, I.; Lamthanh, H.; Stöcklin, R.; Sotto, F.; Menez, A.; Tytgat, J.; Mebs, D. A novel conotoxin inhibiting vertebrate voltage-sensitive potassium channels. Toxicon 2003, 42, 43–52. [Google Scholar] [CrossRef]
- Safavi-Hemami, H.; Gajewiak, J.; Karanth, S.; Robinson, S.D.; Ueberheide, B.; Douglass, A.D.; Schlegel, A.; Imperial, J.S.; Watkins, M.; Bandyopadhyay, P.K.; et al. Specialized insulin is used for chemical warfare by fish-hunting cone snails. Proc. Natl. Acad. Sci. USA 2015, 112, 1743–1748. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.D.; Li, Q.; Bandyopadhyay, P.K.; Gajewiak, J.; Yandell, M.; Papenfuss, A.T.; Purcell, A.W.; Norton, R.S.; Safavi-Hemami, H. Hormone-like peptides in the venoms of marine cone snails. Gen. Comp. Endocrinol. 2015. [Google Scholar] [CrossRef] [PubMed]
- Möller, C.; Melaun, C.; Castillo, C.; Díaz, M.E.; Renzelman, C.M.; Estrada, O.; Kuch, U.; Lokey, S.; Marí, F. Functional hypervariability and gene diversity of cardioactive neuropeptides. J. Biol. Chem. 2010, 285, 40673–40680. [Google Scholar] [CrossRef] [PubMed]
- Mena, E.E.; Gullak, M.F.; Pagnozzi, M.J.; Richter, K.E.; Rivier, J.; Cruz, L.J.; Olivera, B.M. Conantokin-G: A novel peptide antagonist to the N-methyl-D-aspartic acid (NMDA) receptor. Neurosci. Lett. 1990, 118, 241–244. [Google Scholar] [CrossRef]
- Safavi-Hemami, H.; Lu, A.; Li, Q.; Fedosov, A.E.; Biggs, J.; Showers Corneli, P.; Seger, J.; Yandell, M.; Olivera, B.M. Venom insulins of cone snails diversify rapidly and track prey taxa. Mol. Biol. Evol. 2016, 33, 2924–2934. [Google Scholar] [CrossRef] [PubMed]
- Fiedler, B.; Zhang, M.-M.; Buczek, O.; Azam, L.; Bulaj, G.; Norton, R.S.; Olivera, B.M.; Yoshikami, D. Specificity, affinity and efficacy of iota-conotoxin RXIA, an agonist of voltage-gated sodium channels NaV1.2, 1.6 and 1.7. Biochem. Pharmacol. 2008, 75, 2334–2344. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Shao, X.; Guo, Z.Y.; Chi, C.W. Identification of neuropeptide Y-like conopeptides from the venom of Conus betulinus. Acta Biochim. Biophys. Sin. (Shanghai) 2010, 42, 502–505. [Google Scholar] [CrossRef] [PubMed]
- Walker, C.S.; Jensen, S.; Ellison, M.; Matta, J.A.; Lee, W.Y.; Imperial, J.S.; Duclos, N.; Brockie, P.J.; Madsen, D.M.; Isaac, J.T.R.; et al. A novel Conus snail polypeptide causes excitotoxicity by blocking desensitization of AMPA receptors. Curr. Biol. 2009, 19, 900–908. [Google Scholar] [CrossRef] [PubMed]
- Robinson, S.D.; Safavi-Hemami, H.; Raghuraman, S.; Imperial, J.S.; Papenfuss, A.T.; Teichert, R.W.; Purcell, A.W.; Olivera, B.M.; Norton, R.S. Discovery by proteogenomics and characterization of an RF-amide neuropeptide from cone snail venom. J. Proteom. 2015, 114, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Dürr, K.L.; Gouaux, E. X-ray structures of AMPA receptor–cone snail toxin complexes illuminate activation mechanism. Science 2014, 345, 1021–1026. [Google Scholar] [CrossRef] [PubMed]
- England, L.J.; Imperial, J.; Jacobsen, R.; Craig, A.G.; Gulyas, J.; Akhtar, M.; Rivier, J.; Julius, D.; Olivera, B.M. Inactivation of a serotonin-gated ion channel by a polypeptide toxin from marine snails. Science 1998, 281, 575–578. [Google Scholar] [CrossRef] [PubMed]
- Teichert, R.W.; Jimenez, E.C.; Olivera, B.M. αS-Conotoxin RVIIIA: A structurally unique conotoxin that broadly targets nicotinic acetylcholine receptors. Biochemistry 2005, 44, 7897–7902. [Google Scholar] [CrossRef] [PubMed]
- Cruz, L.J.; de Santos, V.; Zafaralla, G.C.; Ramilo, C.A.; Zeikus, R.; Gray, W.R.; Olivera, B.M. Invertebrate vasopressin/oxytocin homologs. Characterization of peptides from Conus geographus and Conus straitus venoms. J. Biol. Chem. 1987, 262, 15821–15824. [Google Scholar] [PubMed]
- Dutertre, S.; Jin, A.H.; Vetter, I.; Hamilton, B.; Sunagar, K.; Lavergne, V.; Dutertre, V.; Fry, B.G.; Antunes, A.; Venter, D.J.; et al. Evolution of separate predation- and defence-evoked venoms in carnivorous cone snails. Nat. Commun. 2014, 5, 3521. [Google Scholar] [CrossRef] [PubMed]
- Phuong, M.A.; Mahardika, G.N.; Alfaro, M.E. Dietary breadth is positively correlated with venom complexity in cone snails. BMC Genom. 2016, 17, 401. [Google Scholar] [CrossRef] [PubMed]
- Barghi, N.; Concepcion, G.P.; Olivera, B.M.; Lluisma, A.O. Comparison of the venom peptides and their expression in closely related Conus species: Insights into adaptive post-speciation evolution of conus exogenomes. Genome Biol. Evol. 2015, 7, 1797–1814. [Google Scholar] [CrossRef] [PubMed]
- Hillyard, D.R.; Olivera, B.M.; Woodward, S.; Corpuz, G.P.; Gray, W.R.; Ramilo, C.A.; Cruz, L.J. A molluscivorous Conus toxin: Conserved frameworks in conotoxins. Biochemistry 1989, 28, 358–361. [Google Scholar] [CrossRef] [PubMed]
- Lavergne, V.; Harliwong, I.; Jones, A.; Miller, D.; Taft, R.J.; Alewood, P.F. Optimized deep-targeted proteotranscriptomic profiling reveals unexplored Conus toxin diversity and novel cysteine frameworks. Proc. Natl. Acad. Sci. USA 2015, 112, E3782–E3791. [Google Scholar] [CrossRef] [PubMed]
- Chang, D.; Olenzek, A.M.; Duda, T.F. Effects of geographical heterogeneity in species interactions on the evolution of venom genes. Proc. R. Soc. Lond. B Biol. Sci. 2015, 282. [Google Scholar] [CrossRef] [PubMed]
- Azam, L.; McIntosh, J.M. Alpha-conotoxins as pharmacological probes of nicotinic acetylcholine receptors. Acta Pharmacol. Sin. 2009, 30, 771–783. [Google Scholar] [CrossRef] [PubMed]
- Callaghan, B.; Haythornthwaite, A.; Berecki, G.; Clark, R.J.; Craik, D.J.; Adams, D.J. Analgesic α-conotoxins Vc1.1 and RgIA inhibit N-type calcium channels in rat sensory neurons via GABBA receptor activation. J. Neurosci. 2008, 28, 10943–10951. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, J.M.; Ghomashchi, F.; Gelb, M.H.; Dooley, D.J.; Stoehr, S.J.; Giordani, A.B.; Naisbitt, S.R.; Olivera, B.M. Conodipine-M, a novel phospholipase A isolated from the venom of the marine snail Conus magus. J. Biol. Chem. 1995, 270, 3518–3526. [Google Scholar] [PubMed]
- McDougal, O.M.; Turner, M.W.; Ormond, A.J.; Poulter, C.D. Three-dimensional structure of conotoxin tx3a: An M-1 branch peptide of the M-superfamily. Biochemistry 2008, 47, 2826–2832. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Liu, J.; Pi, C.; Zeng, X.; Zhou, M.; Jiang, X.; Chen, S.; Ren, Z.; Xu, A. Identification of a novel M-superfamily conotoxin with the ability to enhance tetrodotoxin sensitive sodium currents. Arch. Toxicol. 2009, 83, 925–932. [Google Scholar] [CrossRef] [PubMed]
- Bulaj, G.; Zhang, M.-M.; Green, B.R.; Fiedler, B.; Layer, R.T.; Wei, S.; Nielsen, J.S.; Low, S.J.; Klein, B.D.; Wagstaff, J.D.; et al. Synthetic μO-conotoxin MrVIB blocks TTX-resistant sodium channel NaV1.8 and has a long-lasting analgesic activity. Biochemistry 2006, 45, 7404–7414. [Google Scholar] [CrossRef] [PubMed]
- Shon, K.-J.; Grilley, M.M.; Marsh, M.; Yoshikami, D.; Hall, A.R.; Kurz, B.; Gray, W.R.; Imperial, J.S.; Hillyard, D.R.; Olivera, B.M. Purification, characterization, synthesis, and cloning of the lockjaw peptide from Conus purpurascens venom. Biochemistry 1995, 34, 4913–4918. [Google Scholar] [CrossRef] [PubMed]
- Terlau, H.; Stocker, M.; Shon, K.J.; McIntosh, J.M.; Olivera, B.M. MicroO-conotoxin MrVIA inhibits mammalian sodium channels, but not through site I. J. Neurophysiol. 1996, 76, 1423–1429. [Google Scholar] [PubMed]
- Olivera, B.; Gray, W.; Zeikus, R.; McIntosh, J.; Varga, J.; Rivier, J.; de Santos, V.; Cruz, L. Peptide neurotoxins from fish-hunting cone snails. Science 1985, 230, 1338–1343. [Google Scholar] [CrossRef] [PubMed]
- Sabareesh, V.; Gowd, K.H.; Ramasamy, P.; Sudarslal, S.; Krishnan, K.S.; Sikdar, S.K.; Balaram, P. Characterization of contryphans from Conus loroisii and Conus amadis that target calcium channels. Peptides 2006, 27, 2647–2654. [Google Scholar] [CrossRef] [PubMed]
- Massilia, G.R.; Eliseo, T.; Grolleau, F.; Lapied, B.; Barbier, J.; Bournaud, R.; Molgó, J.; Cicero, D.O.; Paci, M.; Eugenia Schininà, M.; et al. Contryphan-Vn: a modulator of Ca2+-dependent K+ channels. Biochem. Biophys. Res. Commun. 2003, 303, 238–246. [Google Scholar] [CrossRef]
- Hansson, K.; Ma, X.; Eliasson, L.; Czerwiec, E.; Furie, B.; Furie, B.C.; Rorsman, P.; Stenflo, J. The first γ-carboxyglutamic acid-containing contryphan. J. Biol. Chem. 2004, 279, 32453–32463. [Google Scholar] [CrossRef] [PubMed]
- Cruz, L.J.; Ramilo, C.A.; Corpuz, G.P.; Olivera, B.M. Conus peptides: Phylogenetic range of biological activity. Biol. Bull. 1992, 183, 159–164. [Google Scholar] [CrossRef]
- fqtrim, v0.9.4 release. Available online: http://doi.org/10.5281/zenodo.20552 (accessed on 20 May 2017).
- Schmieder, R.; Edwards, R. Quality control and preprocessing of metagenomic datasets. Bioinformatics 2011, 27, 863–864. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Li, B.; Dewey, C.N. RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| δ-Conotoxins | Primary Structure | |
| GmVIA | VKPCRKEGQLCDPIFQNCCRGWNCVLFCV | |
| Vc6.36 (C. victoriae) [13] | GKPCHKEGQLCDPFLQNCCLGWNCVFVCI | |
| Tx6.1 (C. textile) | QVKPCRKEHQLCDLIFQNCCRGWYCVVLSCT | |
| Gm6.1 | CRLGAESCDVISQNCCQGTCVFFCLP | |
| Vc6.41 (C. victoriae) [13] | CRLGAESCDVISQNCCQGTCVFFCLP | |
| TxVIA (C. textile) [55] | WCKQSGEMCNLLDQNCCDGYCIVLVCT | |
| TxVIB (C. textile) [19] | WCKQSGEMCNVLDQNCCDGYCIVFVCT | |
| Venom Insulins | Primary Structure | |
| Con-Ins Gm | A-chain | GIVCECCKNVCTKEEFTEYCPPLTESS * |
| B-chain | SEYSCSYNDPDHPEGKCGPQLSEHVEEKCEEEEAEQGGTNNARSNT * | |
| Con-Ins Vc1 (C. victoriae) [40] | A-chain | GIVCECCKHHCTKEELTEYCH |
| B-chain | HVCWLGDPNHPKGICGPQLSDIVEIRCEEKEAEQGGANNARAYT * | |
| Con-Ins Tx1 (C. textile) [39] | A-chain | GIVCECCKHHCTKEEFTEYCH |
| B-chain | GEHVCWLGDPNHPQGICGPQVADIVEIRCEEKEAEQGGANNARANT * | |
| Superfamily | Cysteine Framework | # Identified in C. gloriamaris | Associated Activities | Reference |
|---|---|---|---|---|
| A | I | 1 | nAChRs inhibitors, GABAB receptor agonists, α1-adrenoceptor inhibitor | [25,58,59] |
| - | XXII | 1 | N.D. | - |
| - | XXXVI | 1 | N.D. | - |
| conantokin (B) | Cysteine-free | 1 | NMDA receptor inhibitors | [42] |
| Con-insulin | - | 1 | Insulin receptor agonists | [39] |
| conodipine | - | 1 | Phospholipase-A2 | [60] |
| cono-NPY | Cysteine-free | 1 | [45] | |
| conorfamide | Cysteine-free | 1 | Convulsions in mice (IC), sensory neuron depolarization | [47] |
| conopressin | Single disulfide | 1 | Vasopressin receptor agonists | [51] |
| I1 | XI | 2 | NaV agonists | [44] |
| I2 | XI | 3 | K+ channel modulators | [37,38] |
| J | XIV | 4 | nAChR inhibitor and KV inhibitor | [29] |
| M | III | 7 | Excitatory symptoms in mice (IC) | [35,61,62] |
| conomarphin (M) | Cysteine-free | 1 | N.D. | - |
| (M) | Single disulfide | 1 | N.D. | - |
| O1 | VI/VII | 12 | NaV agonists, KV blockers, NaV blockers or CaV blockers | [63,64,65,66] |
| O2 | VI/VII | 17 | Neuronal pacemaker modulators | [19] |
| contryphan (O2) | Single disulfide | 1 | Ca2+ channel modulators | [67,68,69] |
| O3 | Cysteine-free | 1 | N.D. | - |
| P | IX | 2 | Hyperactivity and spasticity in mice (IC) | [15] |
| - | XIV | 1 | N.D. | - |
| S | VIII | 1 | 5-HT3 receptor inhibitor, nAChR inhibitor | [49,50] |
| T | V | 19 | NaV inhibitor, presynaptic Ca2+ channel inhibitor (or GPCR modulator), sst3 GPCR antagonist | [22,23,24] |
| - | XIII | 1 | N.D. | - |
| - | X | 1 | Noradrenaline transporter inhibitors | [25] |
| U | VI/VII | 1 | Convulsions in mice (IC) | [70] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robinson, S.D.; Li, Q.; Lu, A.; Bandyopadhyay, P.K.; Yandell, M.; Olivera, B.M.; Safavi-Hemami, H. The Venom Repertoire of Conus gloriamaris (Chemnitz, 1777), the Glory of the Sea. Mar. Drugs 2017, 15, 145. https://doi.org/10.3390/md15050145
Robinson SD, Li Q, Lu A, Bandyopadhyay PK, Yandell M, Olivera BM, Safavi-Hemami H. The Venom Repertoire of Conus gloriamaris (Chemnitz, 1777), the Glory of the Sea. Marine Drugs. 2017; 15(5):145. https://doi.org/10.3390/md15050145
Chicago/Turabian StyleRobinson, Samuel D., Qing Li, Aiping Lu, Pradip K. Bandyopadhyay, Mark Yandell, Baldomero M. Olivera, and Helena Safavi-Hemami. 2017. "The Venom Repertoire of Conus gloriamaris (Chemnitz, 1777), the Glory of the Sea" Marine Drugs 15, no. 5: 145. https://doi.org/10.3390/md15050145
APA StyleRobinson, S. D., Li, Q., Lu, A., Bandyopadhyay, P. K., Yandell, M., Olivera, B. M., & Safavi-Hemami, H. (2017). The Venom Repertoire of Conus gloriamaris (Chemnitz, 1777), the Glory of the Sea. Marine Drugs, 15(5), 145. https://doi.org/10.3390/md15050145