Marine Sponges and Bacteria as Challenging Sources of Enzyme Inhibitors for Pharmacological Applications




Abstract
:1. An Introduction to Enzyme Inhibitors in Marine Environments
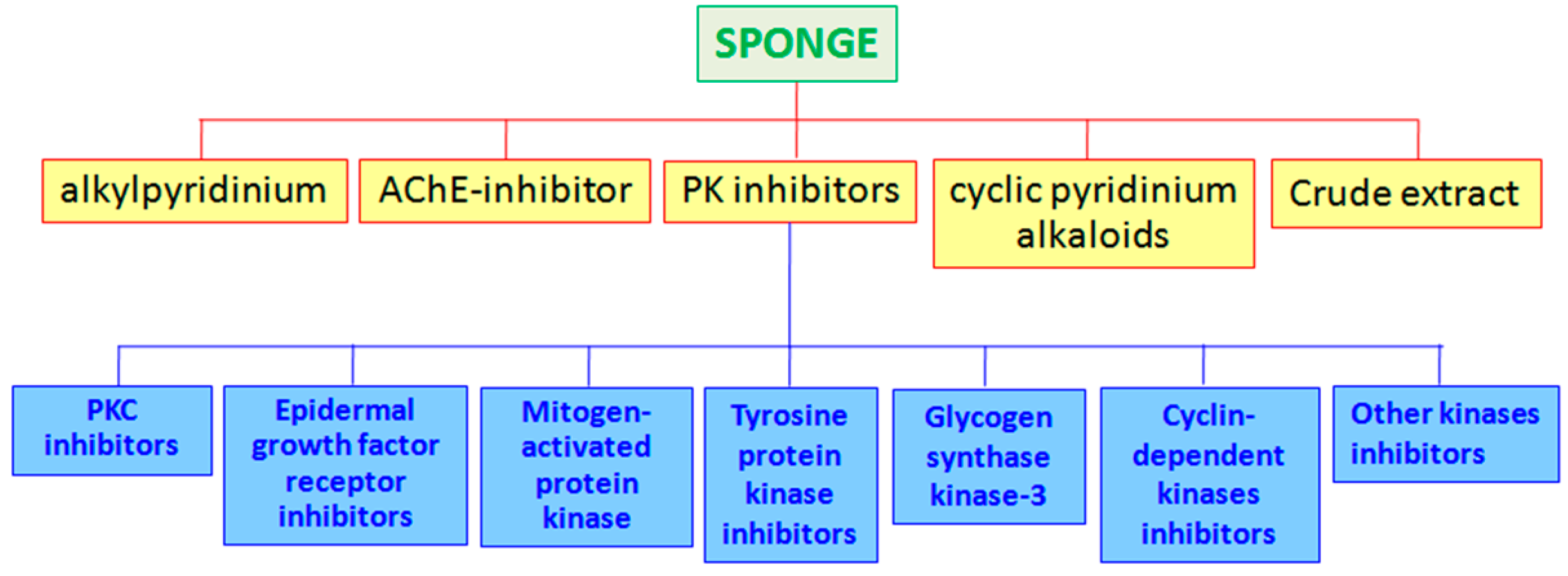
2. Marine Sponges
- -
- -
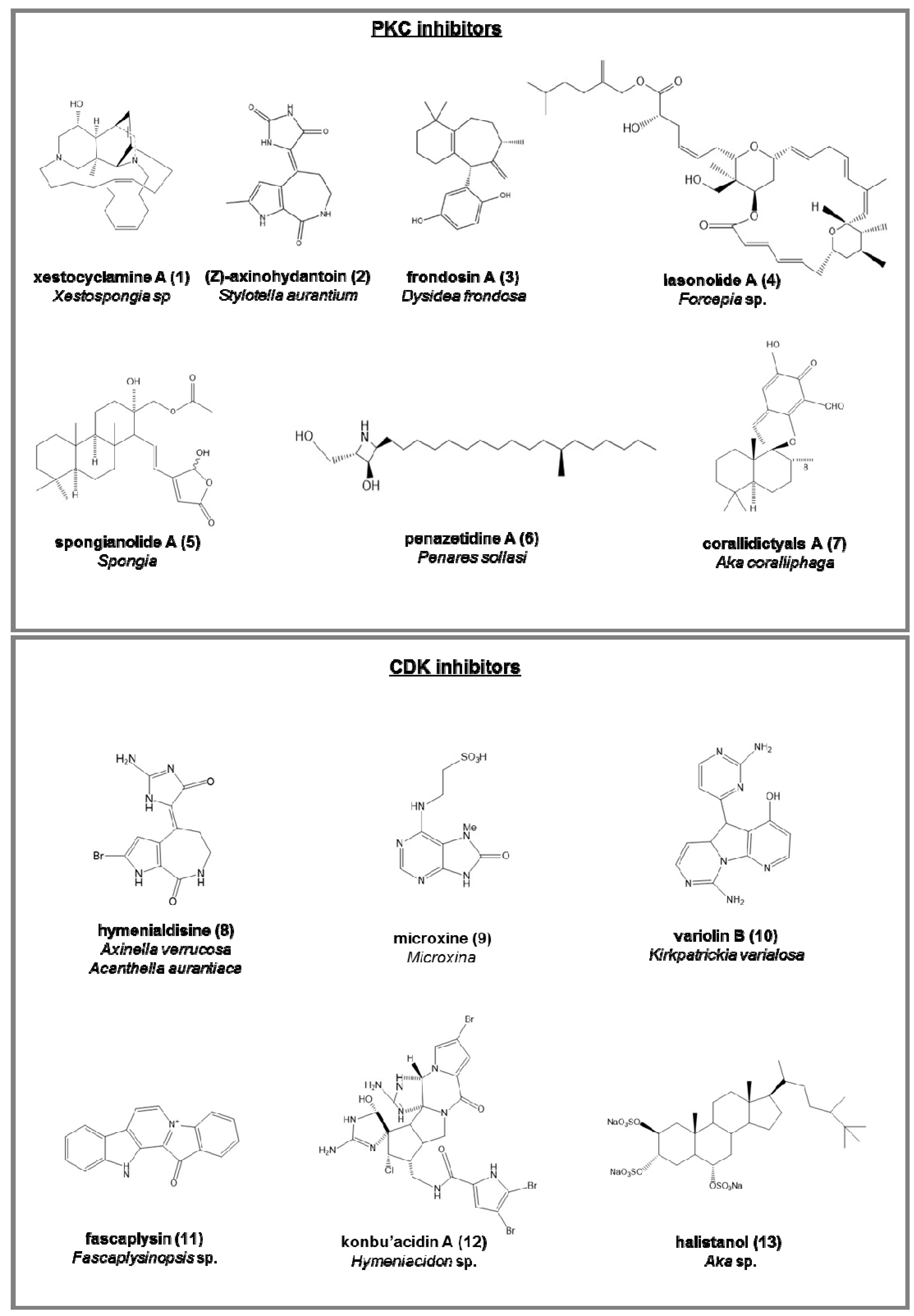
- (Z)-Axinohydantoin (2) and debromo-Z-axinohydantoin, from Stylotella aurantium [45];
- -
- -
- BRS1, a lipid from a calcarea sponge [47];
- -
- -
- -
- -
- penazetidine A (6), new azetidine compound from Penares sollasi, cytotoxic against human and murine cancer cell lines (A549, HT-29, B16/F10 and P388) [55];
- -
- corallidictyals A (7) and B, two diastereomeric spirosesquiterpene aldehydes, from Aka coralliphaga [56], selectively inhibiting the α-PKC isoform.
- -
- hymenialdisine (8), from Axinella verrucosa and Acanthella aurantiaca, able to act on NF-kappa B signaling process and indicated as possible pharmaceuticals in treating rheumatoid arthritis, multiple sclerosis, and Alzheimer’s diseases by some recent patents [57];
- -
- microxine (9), a purine derivative from genus Microxina [58];
- -
- -
- fascaplysin (11), a red pigment isolated from Fascaplysinopsis sp. It is a potent inhibitor of angiogenesis, inhibiting the proliferation of endothelial cells through VEGF [61].
- -
- konbu’acidin A (12), a bromopyrrole alkaloid from the Okinawan Hymeniacidon sp. [62];
- -
- two quinols and halistanol sulfate (13), novel sesquiterpene, from Aka sp. [63];
- -
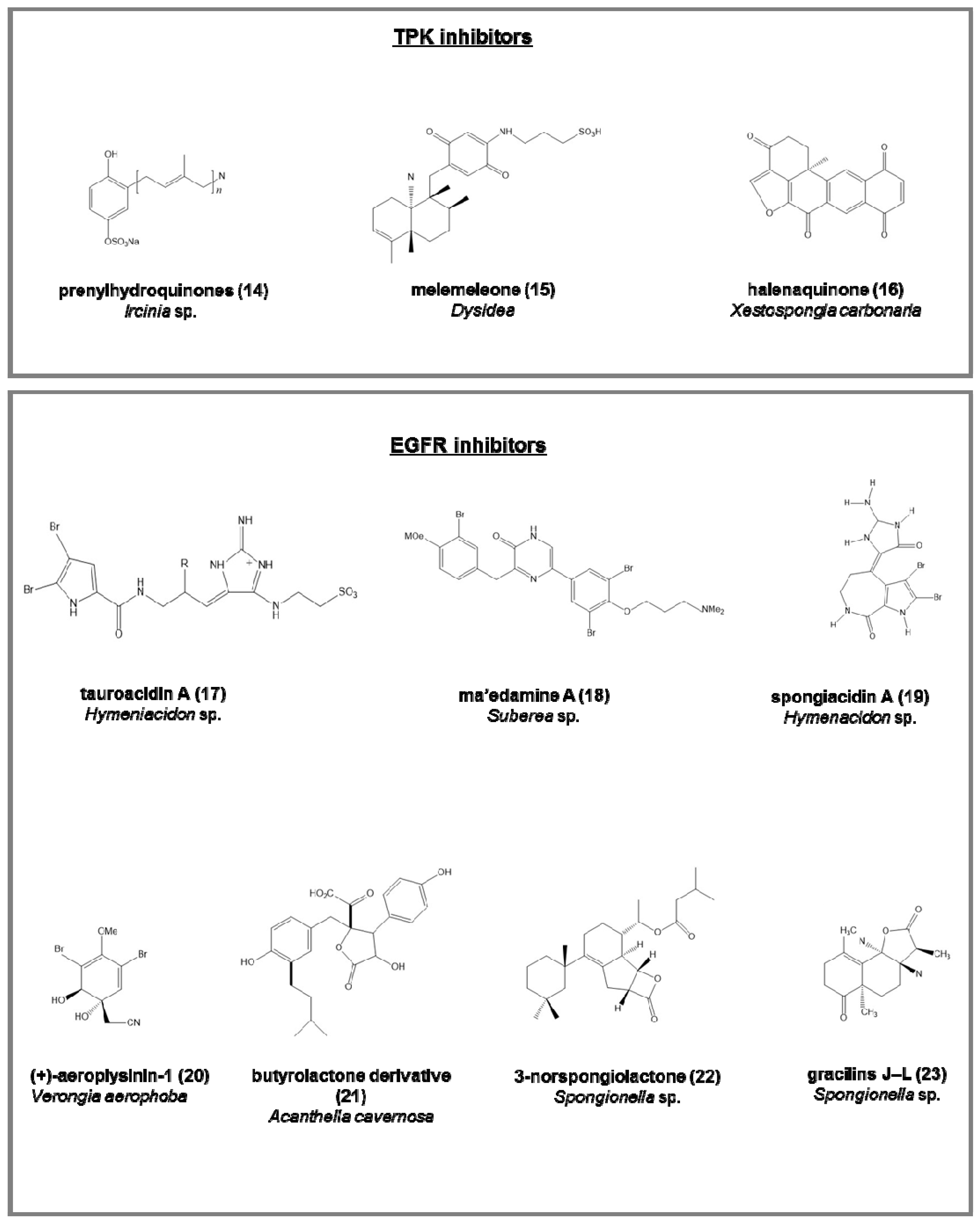
- the penta-, hexa-, and hepta-prenylhydroquinone 4-sulfates (14), from the deep-sea Ircinia sp. [64], with cytotoxic activity against HIV-1 integrase enzyme and epidermal carcinoma cell line;
- -
- melemeleone (15), a novel sesquiterpene, from two species of Dysidea, with activity against tyrosine kinase pp60V-SRC [65];
- -
- halenaquinone (16), halenaquinol, halenaquinol sulfate, and xestoquinone from Xestospongia carbonaria [66].
- -
- tauroacidins A (17) and B, bromopyrrole alkaloids from Hymeniacidon sp. [67];
- -
- ma’edamine A (18), bromotyrosine alkaloid, from Suberea sp. [68];
- -
- spongiacidins A (19) and B, bromopyrrole alkaloids from Hymenacidon sp. [69];
- -
- (+)-aeroplysinin-1 (20), from Verongia aerophoba, antitumoral on EGFR-dependent human breast cancer cell lines MCF-7 and ZR-75-1 [70];
- -
- butyrolactone derivative (21), from Acanthella cavernosa [71];
- -
- 3-norspongiolactone (22) and gracilins J–L (23), bioactive diterpenes from Spongionella sp. [72].
- -
- -
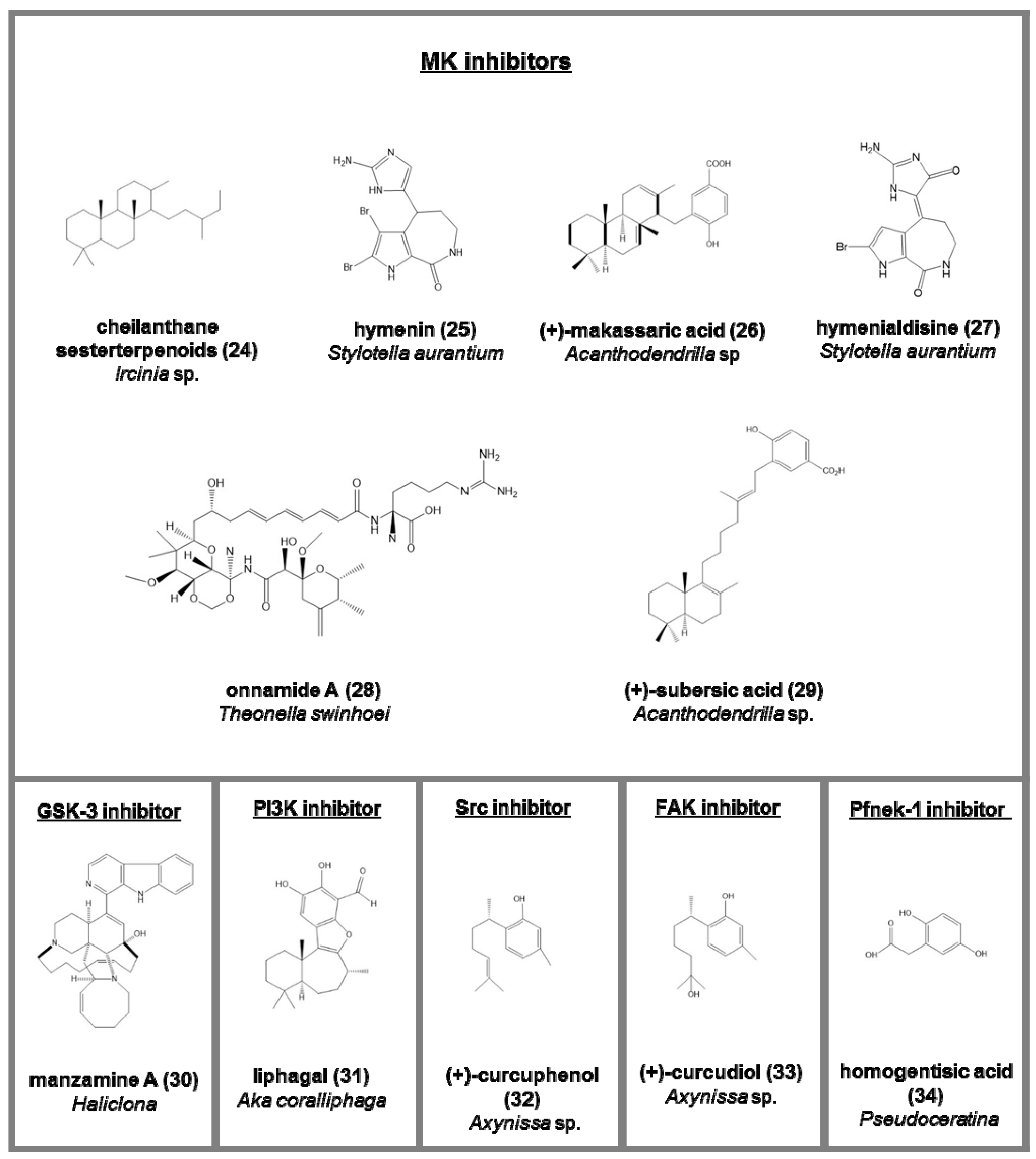
- hymenin (25) and hymenialdisine (27), from Stylotella aurantium [75];
- -
- a methanol fraction from Batzella sp. [76];
- -
- onnamide A (28), heterocyclic compounds belonging to pederin family from Theonella swinhoei. Its activity is through the stimulation of plasminogen activator inhibitor-1 (PAI-1) considered a drug target against metastasis in human cancer cells [77];
- -
- (+)-makassaric acid (26) and (+)-subersic acid (29), meroterpenoids from Acanthodendrilla sp., acting inflammatory responses and cellular stress processes [78].
- -
- -
- liphagal (31), a meroterpenoid from Aka coralliphaga, with inhibitory activity against PI3K and cytotoxic against human colon and human breast cancer [81];
- -
- (+)-curcuphenol (32) (Src protein kinase inhibitor) and (+)-curcudiol (33) (focal adhesion kinase, FAK, inhibitor), two bisabolenes type sesquiterpenoids Axynissa sp. [82];
- -
- -
- hymenialdisine, debromohymenialdisine, and four novel dihydrohymenialdisine derivatives from Cymbastela cantharella, able to inhibit the Polo-Like kinase-1 [84].
3. Marine Bacteria
- -
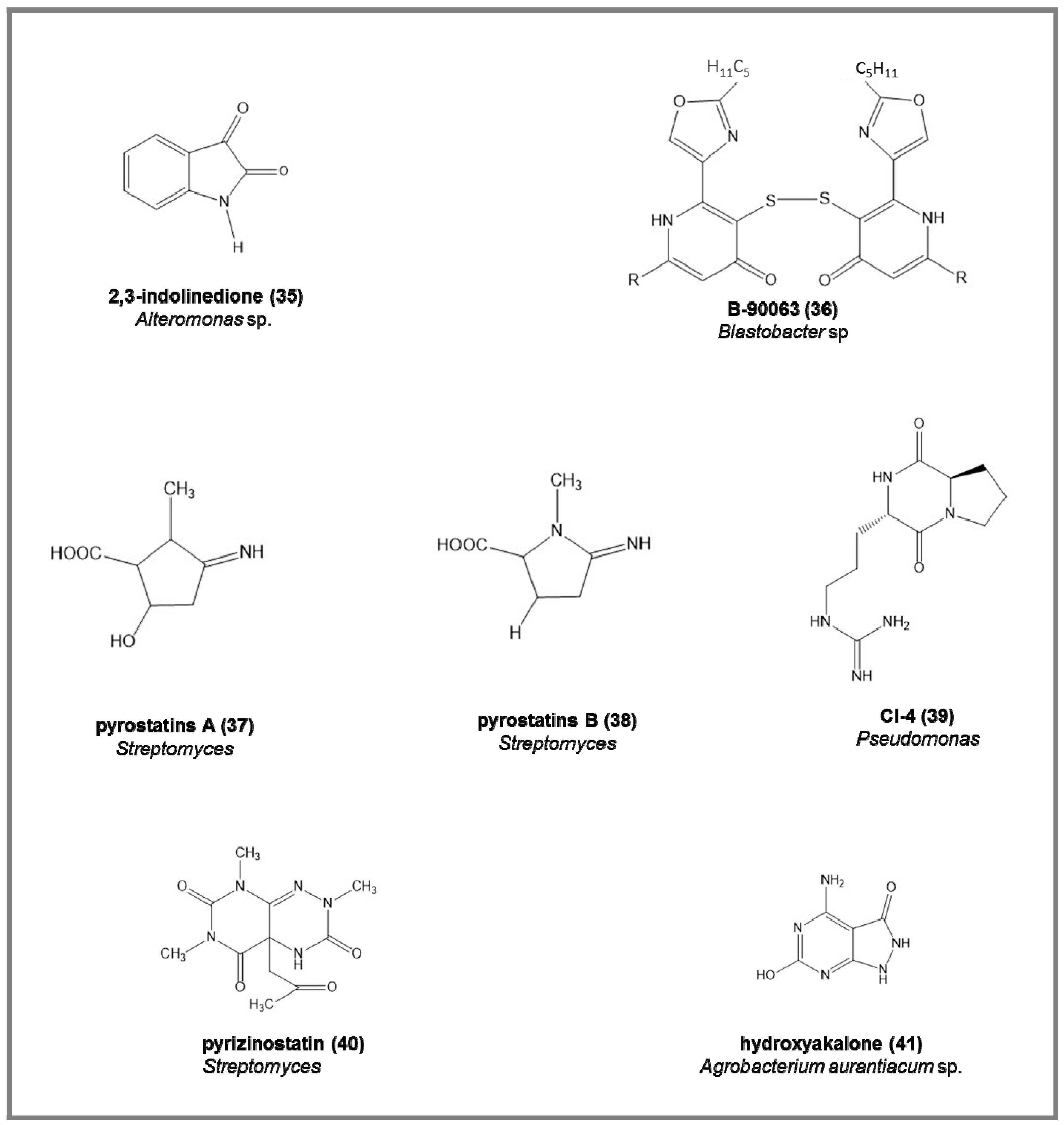
- endogenous monoamine oxidase (MAO) inhibitors, the 2,3-indolinedione (35), from marine Alteromonas sp., increasing the acetylcholine and dopamine in neurotransmission processes (IC50 = 9.2 µM) [88].
- -
- -
- -
- -
- -
- -
- angiotensin-converting enzyme (ACE) inhibitors and adenosine deaminase inhibitors (ADA) [27], from different Streptomyces strains. ACE inhibitors were able to reduce hypertension either by the suppression of angiotensin II biosynthesis or by the stimulation of bradykinin breakdown; ADA inhibitors were responsible for the alteration in adenosine and deoxyadenosine levels and in lymphocytic growth and functions, and enhance the effects of chemotherapeutic effects of adenosine analogs. Among 94 Streptomyces strains screened, 8 and 4 strains were positive for ACE and ADE inhibitors, respectively [27].
- -
4. General Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs over the last 25 years. J. Nat. Prod. 2007, 70, 461–477. [Google Scholar] [CrossRef] [PubMed]
- Skropeta, D.; Pastro, N.; Zivanovic, A. Kinase inhibitors from marine sponges. Mar. Drugs 2011, 9, 2131–2154. [Google Scholar] [CrossRef] [PubMed]
- Bode, W.; Huber, R. Natural protein proteinase inhibitors and their interaction with proteinases. Eur. J. Biochem. 1992, 204, 433–451. [Google Scholar] [CrossRef] [PubMed]
- Terashita, T.; Kono, M.; Murao, S. Promoting effect of S-PI on fruiting of Lentinus edodes. Trans. Mycol. Soc. Jpn. 1980, 21, 137–140. [Google Scholar]
- Demuth, H.U. Recent developments in inhibiting cysteine and serine proteases. J. Enzym. Inhib. 1990, 3, 249–278. [Google Scholar] [CrossRef] [PubMed]
- Delston, R.B.; Kothary, M.H.; Shangraw, K.A.; Tall, B.D. Isolation and characterization of a zinc-containing metalloprotease expressed by Vibrio tubiashii. Can. J. Microbiol. 2003, 49, 525. [Google Scholar] [CrossRef] [PubMed]
- Bo-Linn, G.W.; Santa Ana, C.A.; Morawski, S.G.; Fordtran, J.S. Starch blockers-their effect on calorie absorption from a high-starch meal. N. Engl. J. Med. 1982, 307, 1413–1416. [Google Scholar] [CrossRef] [PubMed]
- Puls, W.; Keup, U. Influence of an a-amylase inhibitor (BAY d 7791) on blood glucose, serum insulin and NEFA in starch loading tests in rats, dogs and man. Diabetologia 1973, 9, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Umezawa, H. Enzyme Inhibitor of Microbial Origin; University of Tokyo Press: Tokyo, Japan, 1972. [Google Scholar]
- Hamato, N.; Takano, R.; Kamei-Hayashi, T.; Imada, C.; Hara, S. Leupeptins produced by the marine Alteromonas sp. B-10-31. Biosci. Biotechnol. Biochem. 1992, 56, 1316–1318. [Google Scholar] [CrossRef]
- Imada, C. Enzyme inhibitors of marine microbial origin with pharmaceutical importance. Mar. Biotechnol. 2004, 6, 193–198. [Google Scholar] [CrossRef] [PubMed]
- Aneiros, A.; Garateix, A. Bioactive peptides from marine sources: Pharmacological properties and isolation procedures. J. Chromatogr. B. Anal. Technol. Biomed. Life Sci. 2004, 15, 41–53. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2012, 29, 144–222. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2014, 31, 160–258. [Google Scholar] [CrossRef] [PubMed]
- Mehbub, M.F.; Lei, J.; Franco, C.; Zhang, W. Marine sponge derived natural products between 2001 and 2010: Trends and opportunities for discovery of bioactives. Mar. Drugs 2014, 12, 4539–4577. [Google Scholar] [CrossRef] [PubMed]
- Reen, F.; Gutierrez-Barranquero, J.; Dobson, A.; Adams, C.; O’Gara, F. Emerging concepts promising new horizons for marine biodiscovery and synthetic biology. Mar. Drugs 2015, 13, 2924–2954. [Google Scholar] [CrossRef] [PubMed]
- Amsler, C.D.; Iken, K.B.; McClintock, J.B.; Baker, B.J. Secondary metabolites from Antarctic marine organisms and their ecological implications. In Marine ChemicalEcology; McClintock, J.B., Baker, J.B., Eds.; CRC Press: Boca Raton, FL, USA, 2001; pp. 267–300. [Google Scholar]
- Lindquist, N. Chemical defense of early stages of benthic marine invertebrates. J. Chem. Ecol. 2002, 28, 1987–2000. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Hu, W.P.; Munro, M.H.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2007, 24, 31–86. [Google Scholar] [CrossRef] [PubMed]
- Faulkner, D.J. Marine pharmacology. Antonie Leeuwenhoek 2000, 77, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2013, 30, 237–323. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2015, 32, 116–211. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2016, 33, 382–431. [Google Scholar] [CrossRef] [PubMed]
- MacLeod, R.A. On the role of inorganic ions in the physiology of marine bacteria, VI. In Advances in Microbiology of the Sea; Droop, M.R., Wood, E.J.F., Eds.; Academic Press: New York, NY, USA, 1968; Volume 1, pp. 95–126. [Google Scholar]
- Maeda, M.; Taga, N. Extracellular nuclease produced by a marine bacterium, II: Purification and properties of extracellular nuclease from a marine Vibrio sp. Can. J. Microbiol. 1976, 22, 1443–1452. [Google Scholar] [CrossRef]
- Kobori, H.; Taga, N. Extracellular alkaline phosphatase from marine bacteria: Purification and properties of extracellular phosphatase from a marine Pseudomonas sp. Can. J. Microbiol. 1980, 26, 833–838. [Google Scholar] [CrossRef]
- Chellaram, C.; Prem Anand, T.; Felicia Shanthini, C.; Chandrika, M.; Gladis, C. Screening for enzyme inhibitors in marine bacteria. Int. J. PharmTech Res. 2014, 6, 351–355. [Google Scholar]
- Agarwal, R.P. Inhibitors of adenosine deaminase. Pharmacol. Ther. 1982, 17, 399–429. [Google Scholar] [CrossRef]
- Sepčić, K.; Marcel, V.; Klaebe, A.; Turk, T.; Šuput, D.; Fournier, D. Inhibition of acetylcholinesterase by an alkylpyridinium polymer from the marine sponge, Reniera sarai. Biochim. Biophys. Acta 1998, 1387, 217–225. [Google Scholar] [CrossRef]
- Lee, Y.K.; Lee, J.-H.; Lee, H.K. Microbial symbiosis in marine sponges. J. Microbiol. 2001, 39, 254–264. [Google Scholar]
- Fusetani, N.; Asai, N.; Matsunaga, S.; Honda, K.; Yasumuro, K. Cyclostellettamines A–F, pyridine alkaloids which inhibit binding of methyl quinuclidinyl benzilate (QNB) to muscarinic acetylcholine receptors, from the marine sponge, Stelletta maxima. Tetrahedron Lett. 1994, 35, 3967–3970. [Google Scholar] [CrossRef]
- Davies-Coleman, M.T.; Faulkner, J.; Dubowchik, G.; Roth, G.P.; Polson, C.; Fairchild, C. A new EGF-active polymeric pyridinium alkaloid from the sponge Callyspongia fibrosa. J. Org. Chem. 1993, 58, 5925–5930. [Google Scholar] [CrossRef]
- Abramson, S.N.; Radic, Z.; Manker, D.; Faulkner, J.; Taylor, P. Onchidal: A naturally occurring irreversible inhibitor of acetylcholinesterase with a novel mechanism of action. Mol. Pharmacol. 1989, 36, 349–354. [Google Scholar] [PubMed]
- Turk, T.; Macek, P.; Sŭput, D. Inhibition of acetylcholinesterase by a pseudozoanthoxanthin-like compound isolated from the zoanthid Parazoanthus axinellae (O. Schmidt). Toxicon 1995, 33, 133–142. [Google Scholar] [CrossRef]
- Seleghim, M.H.R.; Lira, S.P.; Kossuga, M.H.; Batista, T.; Berlinck, R.G.S.; Hajdu, E.; Muricy, G.; Rocha, R.M.; Nascimento, G.F.; Silva, M.; et al. Antibiotic, cytotoxic and enzyme inhibitory activity of crude extracts from Brazilian marine invertebrates. Rev. Bras. Farmacogn. 2007, 17, 287–318. [Google Scholar] [CrossRef]
- Gray, C.A.; de Lira, S.P.; Silva, M.; Pimenta, E.F.; Thiemann, O.H.; Oliva, G.; Hajdu, E.; Andersen, R.J.; Berlinck, R.G.S. Sulfated meroterpenoids from the Brazilian sponge Callyspongia sp. are inhibitors of the antileishmaniasis target adenosine phosphoribosyl transferase. J. Org. Chem. 2006, 71, 8685–8690. [Google Scholar] [CrossRef] [PubMed]
- Duong-Ly, K.C.; Peterson, J.R. The human kinome and kinase inhibition. Curr. Protoc. Pharmacol. 2013. [Google Scholar] [CrossRef]
- Novak, K. Conference report-protein kinase inhibitors in cancer treatment: Mixing and matching? In Proceedings of the Keystone Symposium on Protein Kinases and Cancer, Medscape General Medicine, Lake Tahoe, CA, USA, 24–29 February 2004. [Google Scholar]
- Mariaule, G.; Belmont, P. Cyclin-dependent kinase inhibitors as marketed drugs: Where are we now? A short survey. Molecules 2014, 19, 14366–14382. [Google Scholar] [CrossRef] [PubMed]
- Norman, P. Overview: Kinase Therapeutics Pipelines: An Assessment of Targets and Agents in Development; Cambridge Healthtech Institute: Needham, MA, USA, 2007. [Google Scholar]
- Goldstein, D.; Gray, N.; Zarrinkar, P. High-throughput kinase profiling as a platform for drug discovery. Nat. Rev. Drug Discov. 2008, 7, 391–397. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.; Peters, B.M.; Kurz, L.; Schatzman, R.C.; McCarley, D.; Lou, L.; Crews, P. An alkaloid protein kinase C inhibitor, xestocyclamine A, from the marine sponge Xestospongia sp. J. Am. Chem. Soc. 1993, 115, 10436–10437. [Google Scholar] [CrossRef]
- Rodriguez, J.; Crews, P. Revised structure of xestocyclaminee A and description of new analogue. Tetrahedon Lett. 1994, 35, 4719–4722. [Google Scholar] [CrossRef]
- Yun, H.; Gagnon, A.; Danishefsky, S.J. Toward the synthesis of xestocyclamine A: Investigation of double Michael reaction and direct aza Diels-Alder reaction. Tetrahedron Lett. 2006, 47, 5311–5315. [Google Scholar] [CrossRef]
- Patil, A.D.; Freyer, A.J.; Killmer, L.; Hofmann, G.; Johnson, R.K. Z-Axinohydantoin and debromo-Z-axinohydantoin from the sponge Stylotella aurantium: Inhibitors of protein kinase C. Nat. Prod. Res. 1997, 9, 201–207. [Google Scholar]
- Freyer, A.J.; Patil, A.D.; Killmer, L.; Offen, P.; Carte, B.; Jurewicz, A.J.; Johnson, R.K. Frondosins, five new sesquiterpene hydroquinone derivatives with novel skeletons from the sponge Dysidea frondosa: Inhibitors of interleukin-8 receptors. Tetrahedron 1997, 53, 5047–5060. [Google Scholar]
- Willis, R.H.; de Vries, D.J. BRS1, a C30 bis-amino, bis-hydroxy polyunsaturated lipid from an Australian calcareous sponge that inhibits protein kinase C. Toxicon 1997, 35, 1125–1129. [Google Scholar] [CrossRef]
- Shigemori, H.; Madono, T.; Sasaki, T.; Mikami, Y.; Kobayashi, J. Nakijiquinones A and B, new antifungal sesquiterpenoid quinones with an amino acid residue from an Okinawan marine sponge. Tetrahedron 1994, 50, 8347–8354. [Google Scholar] [CrossRef]
- Kobayashi, J.; Madono, T.; Shigemori, H. Nakijiquinones C and D, new sesquiterpenoid quinones with a hydroxy amino acid residue from a marine sponge inhibiting c-erbB-2 kinase. Tetrahedron 1995, 51, 10867–10874. [Google Scholar] [CrossRef]
- Kissau, L.; Stahl, P.; Mazitschek, R.; Giannis, A.; Waldmann, H. Development of natural product-derived receptor tyrosine kinase inhibitors based on conservation of protein domain fold. J. Med. Chem. 2003, 46, 2917–2931. [Google Scholar] [CrossRef] [PubMed]
- Longley, R.E.; Harmody, D. A rapid colorimetric microassay to detect agonist/antagonists of protein-kinase-C based on adherence of EL-4.IL-2 cells. J. Antibiot. 1991, 44, 93–102. [Google Scholar] [CrossRef] [PubMed]
- Horton, P.A.; Koehn, F.E.; Longley, R.E.; McConnell, O.J. Lasonolide A, a new cytotoxic macrolide from the marine sponge Forcepia sp. J. Am. Chem. Soc. 1994, 116, 6015–6016. [Google Scholar] [CrossRef]
- Isbrucker, R.A.; Guzman, E.A.; Pitts, T.P.; Wright, A.E. Early effects of lasonolide A on pancreatic cancer cells. J. Pharmacol. Exp. Ther. 2009, 331, 733–739. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Kulanthaivel, P.; Baker, B.J. New cytotoxic sesterterpenes from the marine sponge Spongia sp. Tetrahedron Lett. 1994, 35, 7189–7192. [Google Scholar] [CrossRef]
- Alvi, K.A.; Jaspars, M.; Crews, P.; Strulovici, B.; Oto, E. Penazetidine-A, an alkaloid inhibitor of protein kinase C. Bioorg. Med. Chem. Lett. 1994, 4, 2447–2450. [Google Scholar] [CrossRef]
- Chan, J.A.; Freyer, A.J.; Carte, B.K.; Hemling, M.E.; Hofmann, G.A.; Mattern, M.R.; Mentzer, M.A.; Westley, J.W. Protein kinase C inhibitors: Novel spirosesquiterpene aldehydes from a marine sponge Aka (=Siphonodictyon) coralliphagum. J. Nat. Prod. 1994, 57, 1543–1548. [Google Scholar] [CrossRef] [PubMed]
- Meijer, L.; Thunnissen, A.M.; White, A.W.; Garnier, M.; Nikolic, M.; Tsai, L.H.; Walter, J.; Cleverley, K.E.; Salinas, P.C.; Wu, Y.Z.; et al. Inhibition of cyclin-dependent kinases, GSK-3β and CK1 by hymenialdisine, a marine sponge constituent. Chem. Biol. 2000, 7, 51–63. [Google Scholar] [CrossRef]
- Killday, B.; Yarwood, D.; Sills, M.; Murphy, P.; Hooper, J.; Wright, A. Microxine, a new cdc2 kinase inhibitor from the Australian marine sponge Microxima species. J. Nat. Prod. 2001, 64, 525–526. [Google Scholar] [CrossRef] [PubMed]
- Trimurtulu, G.; Faulkner, D.J.; Perry, N.B.; Ettouati, L.; Litaudon, M.; Blunt, J.W.; Munro, M.H.G.; Jameson, G.B. Alkaloids from the antarctic sponge Kirkpatrickia varialosa. Part 2: Variolin A and N(3′)-methyl tetrahydrovariolin B. Tetrahedron 1994, 50, 3993–4000. [Google Scholar] [CrossRef]
- Walker, S.R.; Carter, E.J.; Huff, B.C.; Morris, J.C. Variolins and related alkaloids. Chem. Rev. 2009, 109, 3080–3098. [Google Scholar] [CrossRef] [PubMed]
- Soni, R.; Muller, L.; Furet, P.; Schoepfer, J.; Stephan, C.; Zumstein-Mecker, S.; Fretz, H.; Chaudhuri, B. Inhibition of cyclin-dependent kinase 4 (Cdk4) by fascaplysin, a marine natural product. Biochem. Biophys. Res. Commun. 2000, 275, 877–884. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, J.; Suzuki, M.; Tsuda, M. Konbu’acidin A, a new bromopyrrole alkaloid with cdk4 inhibitory activity from hymeniacidon sponge. Tetrahedron 1997, 53, 15681–15684. [Google Scholar] [CrossRef]
- Mukku, V.; Edrada, R.A.; Schmitz, F.J.; Shanks, M.K.; Chaudhuri, B.; Fabbro, D. New sesquiterpene quinols from a micronesian sponge, Aka sp. J. Nat. Prod. 2003, 66, 686–689. [Google Scholar] [CrossRef] [PubMed]
- Bifulco, G.; Bruno, I.; Minale, L.; Riccio, R.; Debitus, C.; Bourdy, G.; Vassas, A.; Lavayre, J. Bioactive prenylhydroquinone sulfates and a novel C31 furanoterpene alcohol sulfate from the marine sponge, Ircinia sp. J. Nat. Prod. 1995, 58, 1444–1449. [Google Scholar] [CrossRef]
- Alvi, K.A.; Diaz, M.C.; Crews, P.; Slate, D.L.; Lee, R.H.; Moretti, R. Evaluation of new sesquiterpene quinones from two Dysidea sponge species as inhibitors of protein tyrosine kinase. J. Org. Chem. 1992, 57, 6604–6607. [Google Scholar] [CrossRef]
- Lee, R.H.; Slate, D.L.; Moretti, R.; Alvi, K.A.; Crews, P. Marine sponge polyketide inhibitors of protein tyrosine kinase. Biochem. Biophys. Res. Commun. 1992, 184, 765–772. [Google Scholar] [CrossRef]
- Kobayashi, J.; Inaba, K.; Tsuda, M. Tauroacidins A and B, new bromopyrrole alkaloids possessing a taurine residue from hymeniacidon sponge. Tetrahedron 1997, 53, 16679–16682. [Google Scholar] [CrossRef]
- Kobayashi, J.; Hirano, K.; Kubota, T.; Tsuda, M.; Watanabe, K.; Fromont, J. Ma’edamines A and B, cytotoxic bromotyrosine alkaloids with a unique 2(1H)pyrazinone ring from sponge Suberea sp. Tetrahedron 2000, 56, 8107–8110. [Google Scholar]
- Inaba, K.; Sato, H.; Tsuda, M.; Kobayashi, J. Spongiacidins A–D, new bromopyrrole alkaloids from hymeniacidon sponge. J. Nat. Prod. 1998, 61, 693–695. [Google Scholar] [CrossRef] [PubMed]
- Kreuter, M.-H.; Leake, R.E.; Rinaldi, F.; Müller-Klieser, W.; Maidhof, A.; Müller, W.E.G.; Schröder, H.C. Inhibition of intrinsic protein tyrosine kinase activity of EGF-receptor kinase complex from human breast cancer cells by the marine sponge metabolite (+)-aeroplysinin-1. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1990, 97, 151–158. [Google Scholar] [CrossRef]
- Rateb, M.E.; Houssen, W.E.; Legrave, N.M.; Clements, C.; Jaspars, M.; Ebel, R. Dibenzofurans from the marine sponge-derived ascomycete Super1F1-09. Bot. Mar. 2010, 53, 499–506. [Google Scholar] [CrossRef]
- Rateb, M.E.; Houssen, W.E.; Schumacher, M.; Harrison, W.T.A.; Diederich, M.; Ebel, R.; Jaspars, M. Bioactive diterpene derivatives from the marine sponge Spongionella sp. J. Nat. Prod. 2009, 72, 1471–1476. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, M.S.; Edser, A.; King, G.; Whitmore, J.; Quinn, R.J. Cheilanthane sesterterpenes, protein kinase inhibitors, from a marine sponge of the genus Ircinia. J. Nat. Prod. 2001, 64, 300–303. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.; Kesler, C.; Neary, J.; Fishman, L. Effects of marine sponge extracts on mitogen-activated protein kinase (MAPK/ERK1,2) activity in SW-13 human adrenal carcinoma cells. Toxicon 2001, 39, 1835–1839. [Google Scholar] [CrossRef]
- Tasdemir, D.; Mallon, R.; Greenstein, M.; Feldberg, L.; Kim, S.; Collins, K.; Wojciechowicz, D.; Mangalindan, G.; Concepcion, G.; Harper, M.K.; et al. Aldisine alkaloids from the Philippine sponge Stylissa massa are potent Inhibitors of mitogen-activated protein kinase kinase-1 (MEK-1). J. Med. Chem. 2002, 45, 529–532. [Google Scholar] [CrossRef] [PubMed]
- Freitas, J.; Malpezzi, E.; Costa, L.; Berlinck, R.; Almeida, A.; Ogawa, C.; Sanchez, M.; Hajdu, E. Cytotoxic and neurotoxic effects induced by halitoxin isolated from Amphimedon viridis (Porifera). Toxicon 1996, 34, 335. [Google Scholar] [CrossRef]
- Lee, K.H.; Nishimura, S.; Matsunaga, S.; Fusetani, N.; Horinouchi, S.; Yoshida, M. Inhibition of protein synthesis and activation of stress-activated protein kinases by onnamide A and theopederin B, antitumor marine natural products. Cancer Sci. 2005, 96, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Williams, D.E.; Telliez, J.B.; Liu, J.; Tahir, A.; van Soest, R.; Andersen, R.J. Meroterpenoid MAPKAP (MK2) inhibitors isolated from the Indonesian marine sponge Acanthodendrilla sp. J. Nat. Prod. 2004, 67, 2127–2129. [Google Scholar] [CrossRef] [PubMed]
- Hamann, M.; Alonso, D.; Martin-Aparicio, E.; Fuertes, A.; Perez-Puerto, M.; Castro, A.; Morales, S.; Navarro, M.; Monte-Millan, M.; Medina, M.; et al. Glycogen synthase kinase-3 (GSK-3) inhibitory activity and structure activity relationship (SAR). Studies of the manzamine alkaloids. Potential for Alzheimer’s disease. J. Nat. Prod. 2007, 70, 1397–1405. [Google Scholar] [CrossRef] [PubMed]
- Ang, K.; Holmes, M.; Higa, T.; Hamann, M.; Kara, U. In vivo antimalarial activity of the beta-carboline alkaloid manzamine A. Antimicrob. Agents Chemother. 2000, 44, 1645–1649. [Google Scholar] [CrossRef] [PubMed]
- Marion, F.; Williams, D.E.; Patrick, B.O.; Hollander, I.; Mallon, R.; Kim, S.C.; Roll, D.M.; Feldberg, L.; Van Soest, R.; Andersen, R.J. Liphagal, a selective inhibitor of PI3 kinase alpha isolated from the sponge Aka coralliphaga: Structure elucidation and biomimetic synthesis. Org. Lett. 2006, 8, 321–324. [Google Scholar] [CrossRef] [PubMed]
- Hertiani, T.; Edrada-Ebel, R.A.; Kubbutat, M.H.G.; van Soest, R.W.M.; Proksch, P. Protein kinase inhibitors from Indonesian sponge Axynissa sp. Maj. Farm. Indones. 2008, 19, 78–85. [Google Scholar]
- Lebouvier, N.; Jullian, V.; Desvignes, I.; Maurel, S.; Parenty, A.; Dorin-Semblat, D.; Doerig, C.; Sauvain, M.; Laurent, D. Antiplasmodial activities of homogentisic acid derivative protein kinase inhibitors isolated from a vanuatu marine sponge Pseudoceratina sp. Mar. Drugs 2009, 7, 640–653. [Google Scholar] [CrossRef] [PubMed]
- Sauleau, P.; Retailleau, P.; Nogues, S.; Carletti, I.; Marcourt, L.; Raux, R.; Al Mourabit, A.; Debitus, C. Dihydrohymenialdisines, new pyrrole-2-aminoimidazole alkaloids from the marine sponge Cymbastela cantharella. Tetrahedron Lett. 2011, 52, 2676–2678. [Google Scholar] [CrossRef]
- Imada, C.; Taga, N.; Maeda, M. Cultivation conditions for subtilisin inhibitor-producing bacterium and general properties of the inhibitor “marinostatin”. Bull. Jpn. Soc. Sci. Fish 1985, 51, 805–810. [Google Scholar] [CrossRef]
- Imada, C.; Hara, S.; Maeda, M.; Simidu, U. Aminoacid sequences of marinostatins C-1 and C-2 from marine Alteromonas sp. Bull. Jpn. Soc. Sci. Fish 1986, 52, 1455–1459. [Google Scholar] [CrossRef]
- Imada, C.; Maeda, M.; Hara, S.; Taga, N.; Simidu, U. Purification and characterization of subtilisin inhibitors ‘‘marinostatin’’ produced by marine Alteromonas sp. J. Appl. Bacteriol. 1986, 60, 469–476. [Google Scholar] [CrossRef]
- Gil-Turnes, M.S.; Hay, M.E.; Fenical, W. Symbiotic marine bacteria defend crustacean embryos from a pathogenic fungus. Science 1989, 240, 116–118. [Google Scholar] [CrossRef]
- Imada, C.; Maeda, M.; Taga, N. Purification and characterization of the protease inhibitor “monastatin” from a marine Alteromonas sp. with reference to inhibition of the protease produced by a bacterium pathogenic to fish. Can. J. Microbiol. 1985, 31, 1089–1094. [Google Scholar] [CrossRef]
- Imada, C.; Nishimoto, S.; Hara, S. The effect of addition of protease inhibitor from marine bacterium on the strength of gel formation of sardine meat gel (Kamaboko). Nippon Suisan Gakkaishi 2001, 67, 85–89. (In Japanese) [Google Scholar] [CrossRef]
- Takaishi, S.; Tuchiya, N.; Sato, A.; Negishi, T.; Takamatsu, Y.; Matsushita, Y.; Watanabe, T.; Iijima, Y.; Haruyama, H.; Kinoshita, T.; et al. B-90063, a novel endothelin converting enzyme inhibitor isolated from a new marine bacterium, Blastobacter sp. SANK 71894. J. Antibiot. 1998, 51, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, K.; Hasegawa, H.; Umekawa, K.; Ueki, Y.; Ohashi, N.; Kanaoka, M. Design, synthesis and biological activity of novel non-peptidyl endothelin converting enzyme inhibitors, 1-phenyl-tetrazole-formazan analogues. Bioorg. Med. Chem. Lett. 2002, 12, 1275–1278. [Google Scholar] [CrossRef]
- Aoyama, T.; Kojima, F.; Imada, C.; Muraoka, Y.; Naganawa, H.; Okami, Y.; Takeuchi, T.; Aoyagi, T. Pyrostatins A and B, new inhibitors of N-acetyl-b-glucosaminidase, produced by Streptomyces sp. SA-3501. J. Enzym. Inhib. 1995, 8, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Alhadeff, J.A.; Holzinger, R.T. Characterization of bhexosaminidase from liver and sera of diabetic patients and control. Biochem. Med. 1983, 27, 214–225. [Google Scholar] [CrossRef]
- Drexler, H.G.; Gaedicke, G. Isoenzyme studies in human leukemia, III: B-hexosaminidase (E.C. 3.3.1.30). Leuk. Res. 1983, 7, 611–619. [Google Scholar] [CrossRef]
- Pluncinsky, M.C.; Prorok, J.J.; Alhadeff, J.A. Variant serum beta-hexosaminidase as a biochemical marker of malignancy. Cancer 1986, 58, 1484–1487. [Google Scholar] [CrossRef]
- Aoyagi, T.; Hatsu, M.; Imada, C.; Naganawa, H.; Okami, Y.; Takeuchi, T. Pyrizinostatin: A new inhibitor of pyroglutamyl peptidase. J. Antibiot. 1992, 45, 1795–1796. [Google Scholar] [CrossRef] [PubMed]
- Prasad, C.A.; Peterkofsky, A. Determination of pyroglutamylpeptidase and amidase activities toward thyrotropinreleasing hormone in hamster hypothalamus extracts. J. Biol. Chem. 1976, 251, 3229–3234. [Google Scholar] [PubMed]
- Izumida, H.; Miki, W.; Sano, H.; Endo, M. Agar plate method, a new assay for chitinase inhibitors using a chitindegrading bacterium. J. Mar. Biotechnol. 1995, 2, 163–166. [Google Scholar]
- Izumida, H.; Imamura, N.; Sano, H. A novel chitinase inhibitor from a marine bacterium, Pseudomonas sp. J. Antibiot. 1996, 49, 76–80. [Google Scholar] [CrossRef] [PubMed]
- Van Aalten, D.M.F.; Komander, D.; Systad, B.; Gaseidnes, S.; Peter, M.G.; Eijsink, V.G.H. Structural insights into the catalytic mechanism of a family 18 exo-chitinase. Proc. Natl. Acad. Sci. USA 2001, 98, 8979–8984. [Google Scholar] [CrossRef] [PubMed]
- Izumida, H.; Adachi, K.; Nishijima, M.; Endo, M.; Miki, W. Akalone: A novel xanthine oxidase inhibitor produced by the marine bacterium, Agrobacterium aurantiacum sp. nov. J. Mar. Biotechnol. 1985, 2, 115–118. [Google Scholar]
- Borges, F.; Fernandes, E.; Roleira, F. Progress towards the discovery of xanthine oxidase inhibitors. Curr. Med. Chem. 2002, 9, 195–217. [Google Scholar] [CrossRef] [PubMed]
- Ganesan, S.; Raja, S.; Sampathkumar, P.; Sivakumar, K.; Thangaradjou, T. Isolation and screening of α-glucosidase enzyme inhibitor producing marine actinobacteria. Afr. J. Microbiol. Res. 2011, 5, 3437–3445. [Google Scholar] [CrossRef]
- Wahyudi, A.T.; Mubarik, N.R. Screening and characterization of protease inhibitors from marine bacteria associated with sponge Jaspis sp. HAYATI J. Biosci. 2010, 4, 173–178. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Kinase Enzyme | Compound | Reference | Pharmacological Application | IC50 (μM) |
|---|---|---|---|---|
| PKC | ||||
| xestocyclamine A (1) | [42,43,44] | anticancer | 10 | |
| (Z)-Axinohydantoin (2) | [45] | 9 | ||
| frondosin A (3) | [46] | HIV virus | 1.8 | |
| frondosin B | [46] | HIV virus | 4.8 | |
| frondosin C | [46] | HIV virus | 20.9 | |
| frondosin D | [46] | HIV virus | 26 | |
| frondosin E | [46] | HIV virus | 30.6 | |
| nakijiquinones A | [48,49] | anticancer | 270 * | |
| nakijiquinones B | [48,49] | anticancer | 200 * | |
| nakijiquinones C | [48,49] | anticancer | 23 | |
| nakijiquinones D | [48,49,50] | anticancer | 220 * | |
| lasonolide A (4) | [51,52,53] | thymoma cells | 0.03 ** | |
| spongianolides A (5) –E | [54] | 20–30 | ||
| penazetidine A (6) | [55] | anticancer | 1 | |
| corallidictyals A (7) –B | [56] | 28 | ||
| CDK | ||||
| hymenialdisine (8) | [57] | rheumatoid arthritis | 0.02 ** | |
| microxine (9) | [58] | 13 | ||
| variolin B (10) | [59,60] | antiviral, anticancer | 0.03 | |
| fascaplysin (11) | [61] | anticancer, angiogenesis | 0.4 | |
| konbu’acidin A (12) | [62] | anticancer | 20 | |
| halistanol (13) | [63] | anticancer | 0.013 ** | |
| penta-prenylhydroquinone 4-sulfates (14) | [64] | antiviral, anticancer | 8 | |
| hexa-prenylhydroquinone 4-sulfates | [64] | antiviral, anticancer | 4 | |
| hepta-prenylhydroquinone 4-sulfates | [64] | antiviral, anticancer | 8 | |
| melemeleone (15) | [65] | anticancer | 28 | |
| halenaquinone (16) | [66] | anticancer | 1.5 | |
| EGFR | ||||
| tauroacidin A (17) | [67] | anticancer | 0.001 ** | |
| ma’edamine A (18) | [68] | anticancer | 11 | |
| spongiacidin A (19) | [69] | 8.5 | ||
| spongiacidin B | [69] | 6 | ||
| (+)-aeroplysinin-1 (20) | [70] | anticancer | 0.25–0.5 | |
| butyrolactone derivative (21) | [71] | anticancer | 22.9 | |
| 3-norspongiolactone (22) | [72] | anticancer | 0.6–15 | |
| gracilins J–L (23) | [73] | anticancer | 0.6–15 | |
| MK | ||||
| cheilanthane (24) | [74] | anticancer | 4 | |
| hymenin (25) | [75] | 128.8–250 * | ||
| hymenialdisine (27) | [76] | antitumor | 0.003–0.006 ** | |
| onnamide A (28) | [77] | anti-inflammatory | 30 | |
| (+)-makassaric acid (26) | [78] | anti-inflammatory | 20 | |
| (+)-subersic acid (29) | [78] | anti-inflammatory | 9.6 | |
| GSK-3 | ||||
| manzamine A (30) | [79,80] | Alzheimer’s disease | 10.2 | |
| Other kinases inhibitors | ||||
| liphagal (31) | [81] | anticancer | 0.1 | |
| (+)-curcuphenol (32) | [82] | 36 | ||
| homogentisic acid (34) | [83] | antimalarial | 1.8 | |
| Kinase Enzyme | Compound | Reference | Application | IC50 (μM) |
|---|---|---|---|---|
| Serine protease | marinostatins C1–C2 | [86,87] | pancreatitis pathogenesis | 1.0–3.2 |
| Monoamine oxidase | 2,3-indolinedone (35) | [88] | neurodegenrative diseases | 9.2 |
| Protease | monostatin | [89,90] | cooked fish meat gel | |
| ECE | B-90063 (36) | [92] | hypertension, renal disease | 1.0–3.2 |
| GluNAc-ase | pyrostatins A–B (37,38) | [93,94,95,96] | diabetes, leukemia, cancer | 1 |
| PGP | pyrizinostatin (40) | [97,98] | hormone diseases | 21 |
| Chitinase | CI-4 (39) | [99,100,101] | antifungal and insecticidal | |
| Xanhine oxidase | hydroxyakalone (41) | [102,103] | uric acid accumulation | 4.6 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ruocco, N.; Costantini, S.; Palumbo, F.; Costantini, M. Marine Sponges and Bacteria as Challenging Sources of Enzyme Inhibitors for Pharmacological Applications. Mar. Drugs 2017, 15, 173. https://doi.org/10.3390/md15060173
Ruocco N, Costantini S, Palumbo F, Costantini M. Marine Sponges and Bacteria as Challenging Sources of Enzyme Inhibitors for Pharmacological Applications. Marine Drugs. 2017; 15(6):173. https://doi.org/10.3390/md15060173
Chicago/Turabian StyleRuocco, Nadia, Susan Costantini, Flora Palumbo, and Maria Costantini. 2017. "Marine Sponges and Bacteria as Challenging Sources of Enzyme Inhibitors for Pharmacological Applications" Marine Drugs 15, no. 6: 173. https://doi.org/10.3390/md15060173
APA StyleRuocco, N., Costantini, S., Palumbo, F., & Costantini, M. (2017). Marine Sponges and Bacteria as Challenging Sources of Enzyme Inhibitors for Pharmacological Applications. Marine Drugs, 15(6), 173. https://doi.org/10.3390/md15060173