Phytosterols from Dunaliella tertiolecta Reduce Cell Proliferation in Sheep Fed Flaxseed during Post Partum

 , ,
, , 
Abstract
:1. Introduction
2. Results
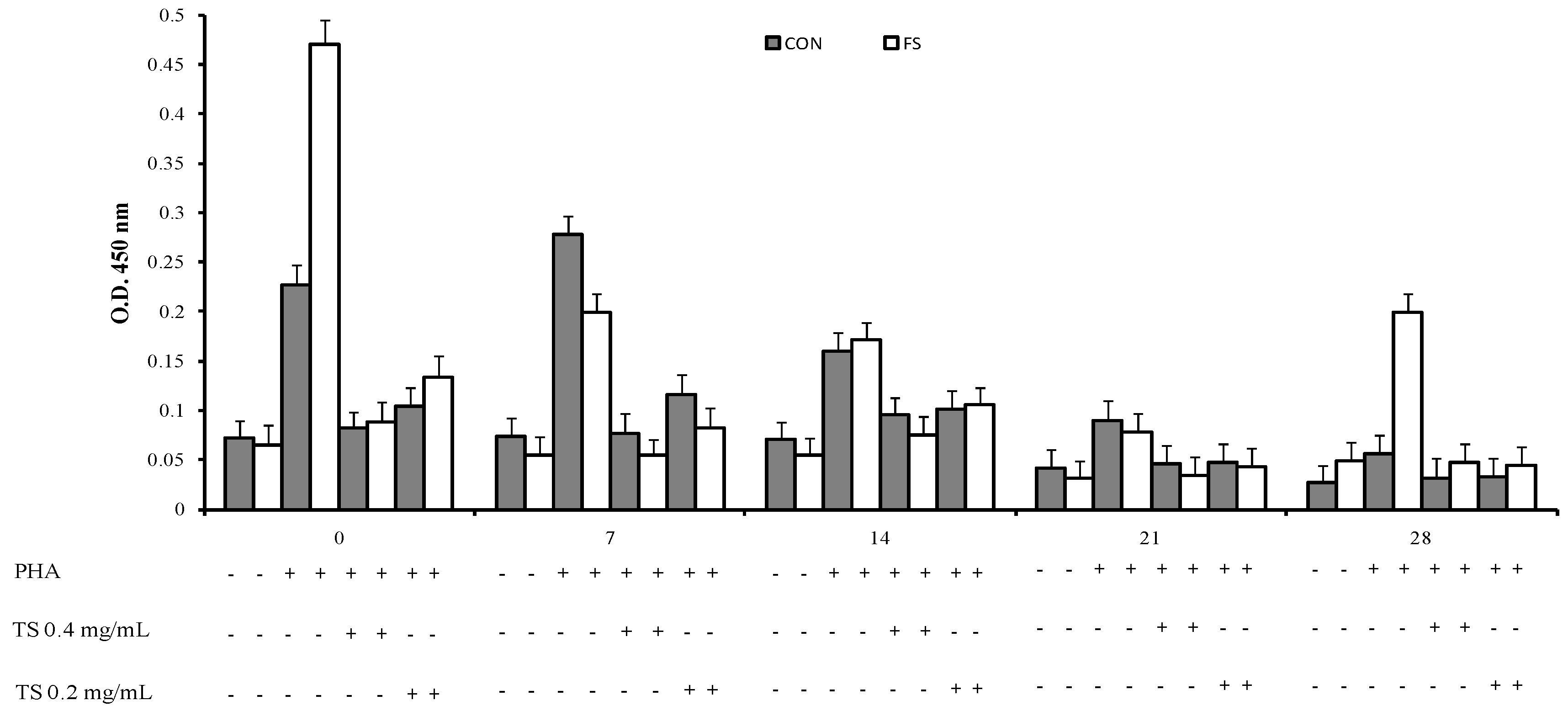
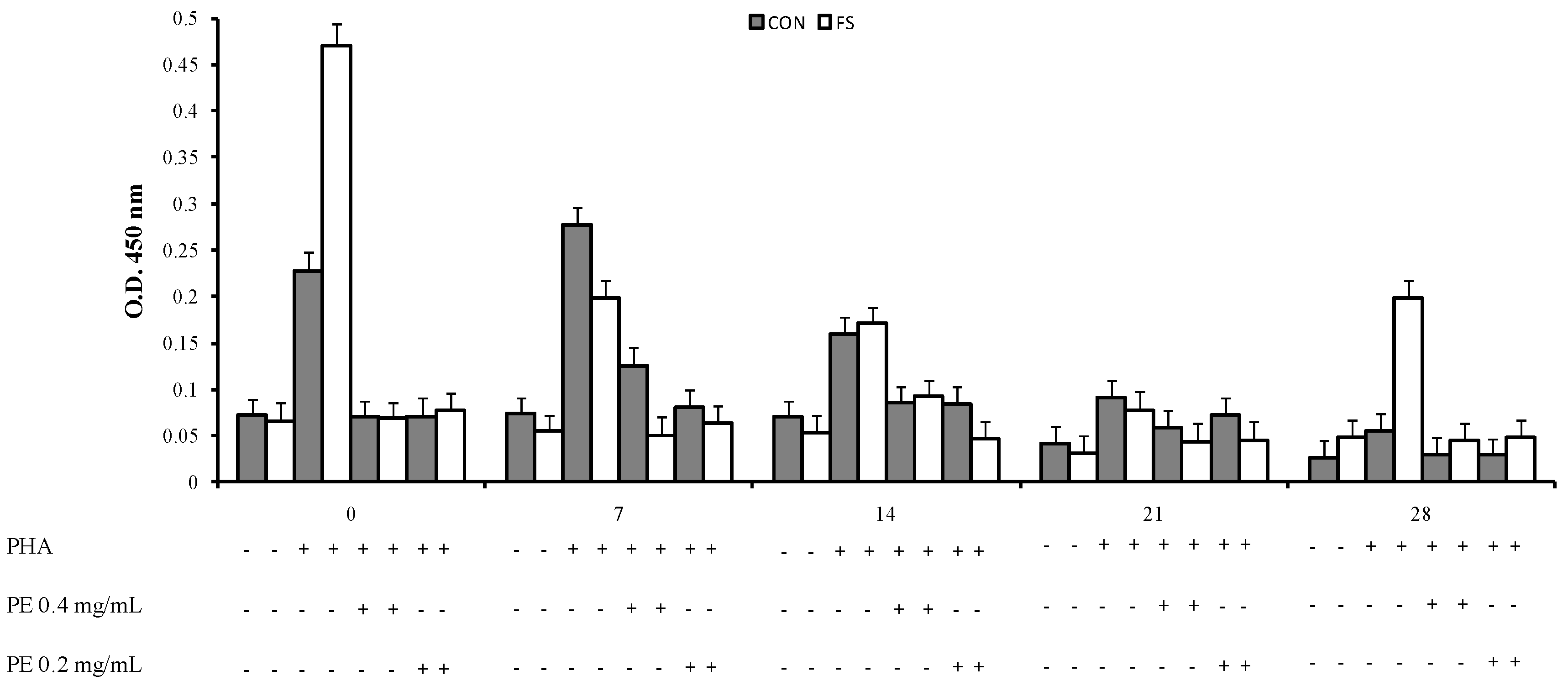
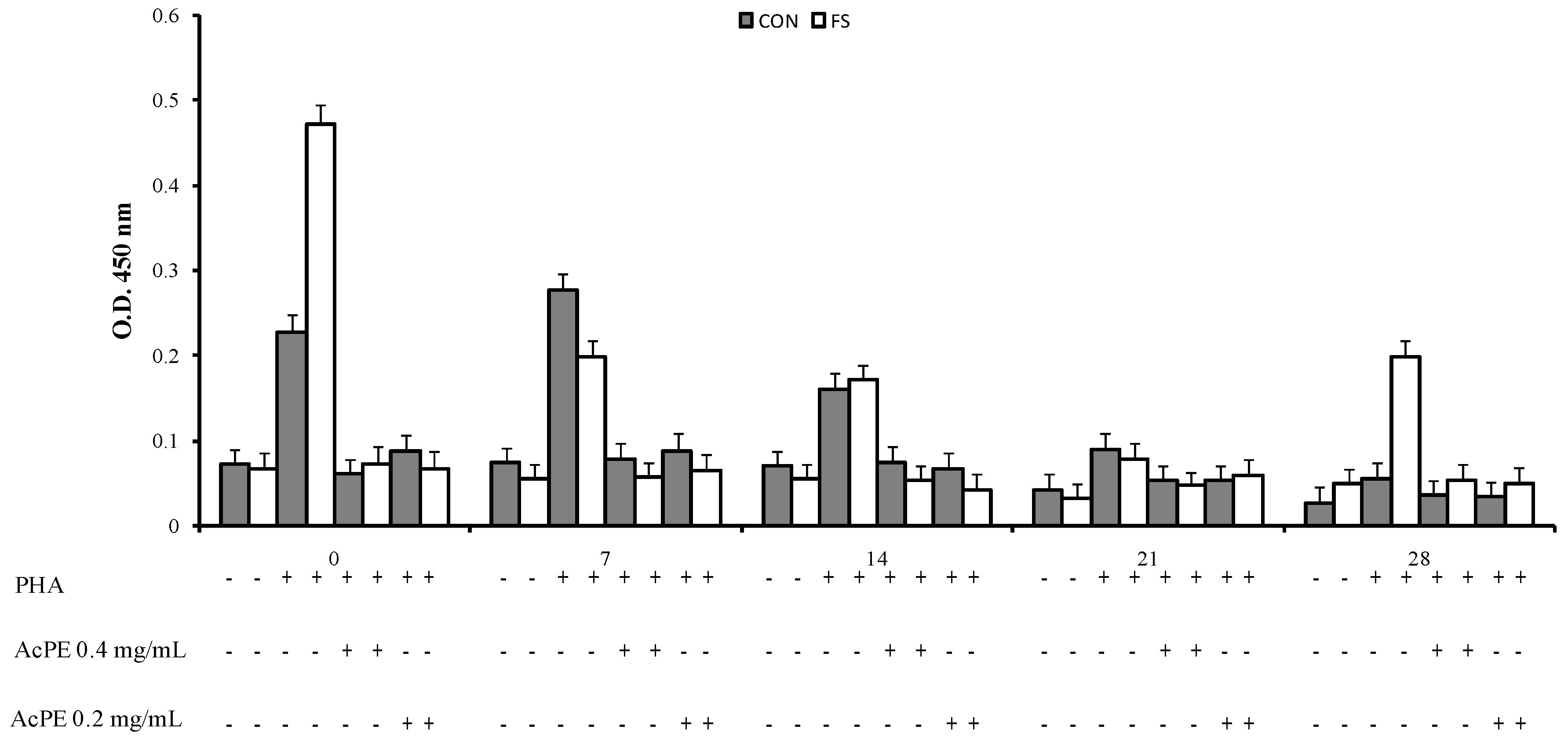
2.1. Proliferative Response to Phytohemoagglutinin (PHA)
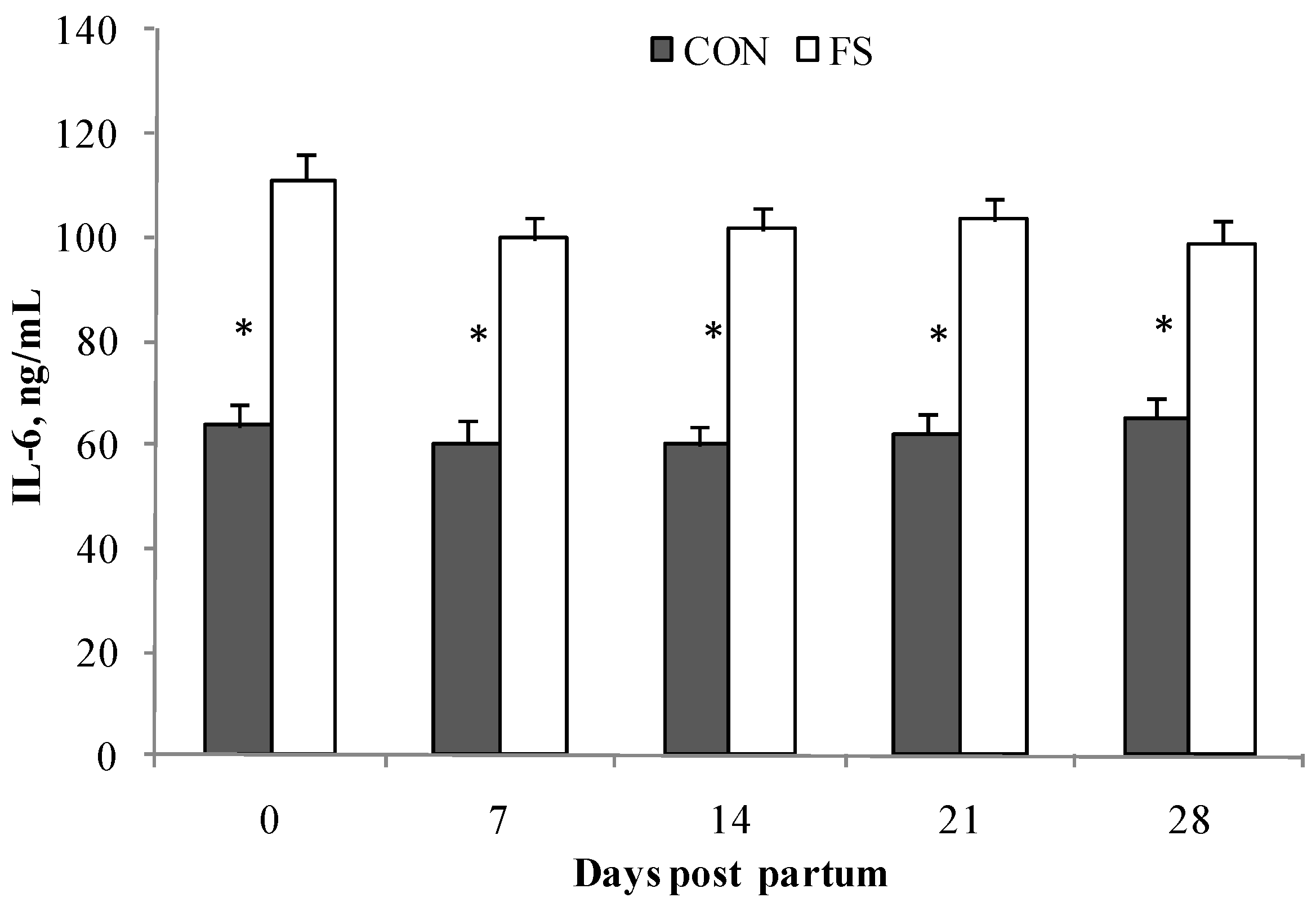
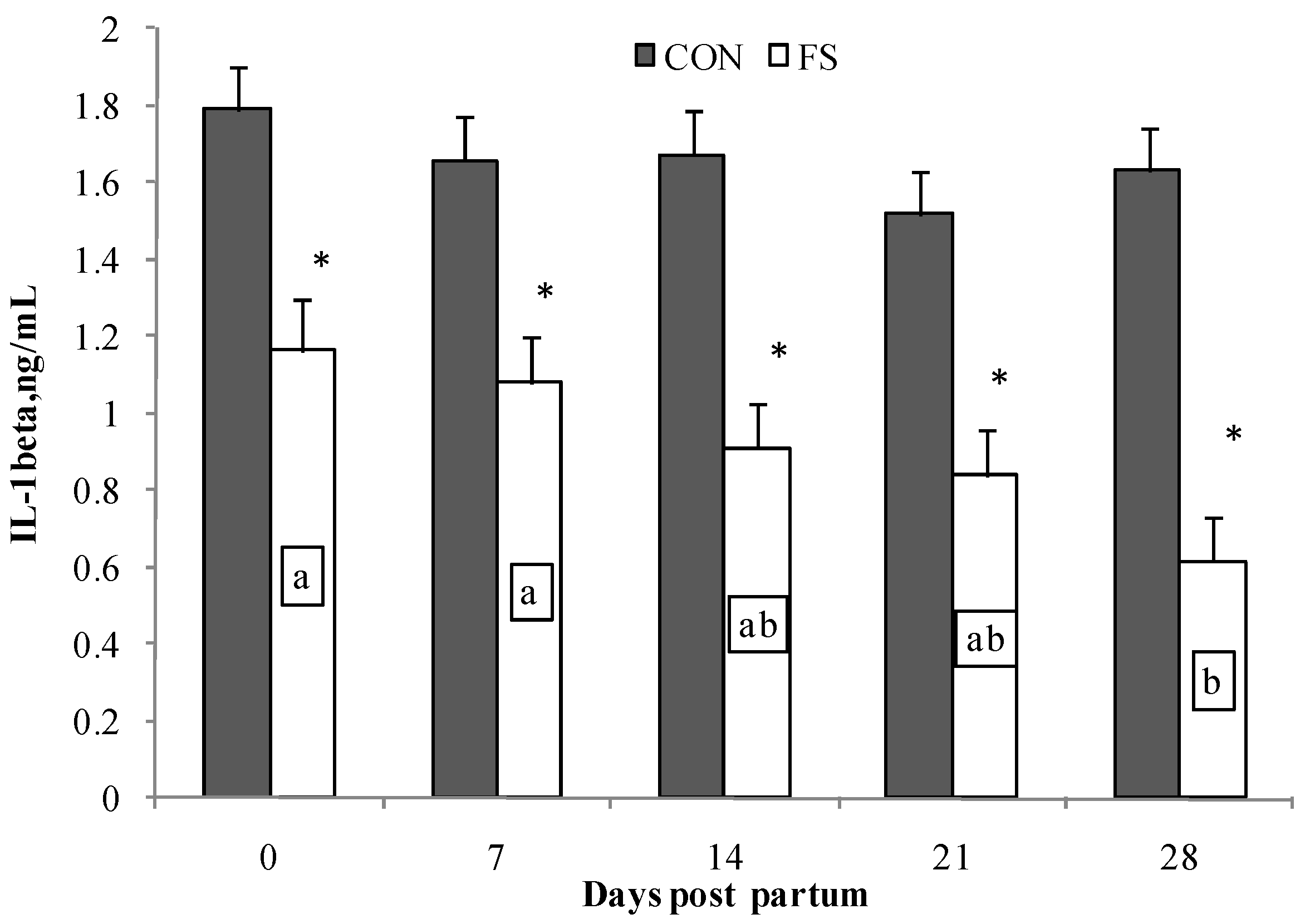
2.2. Cytokine Production by PBMC
3. Discussion
4. Materials and Methods
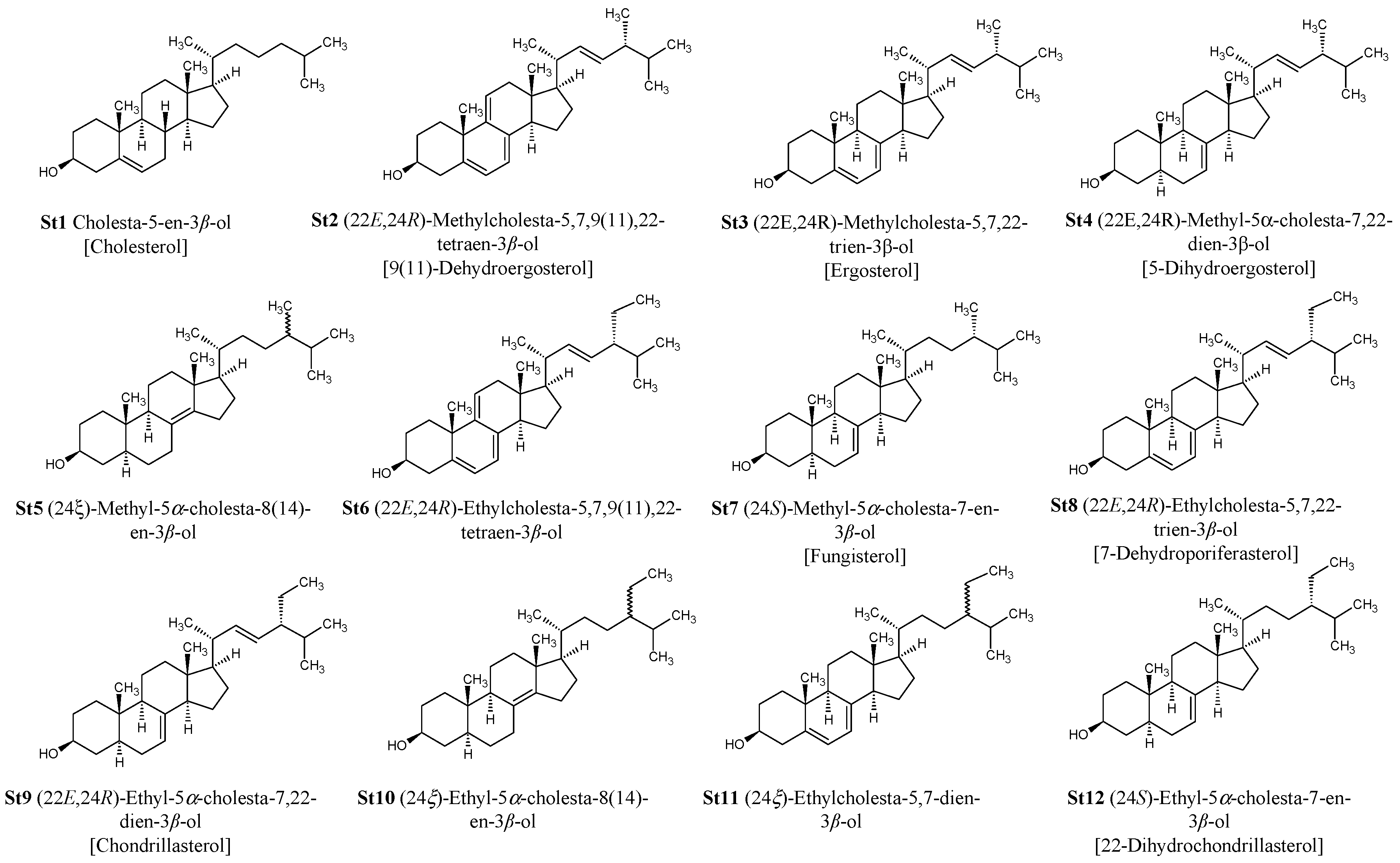
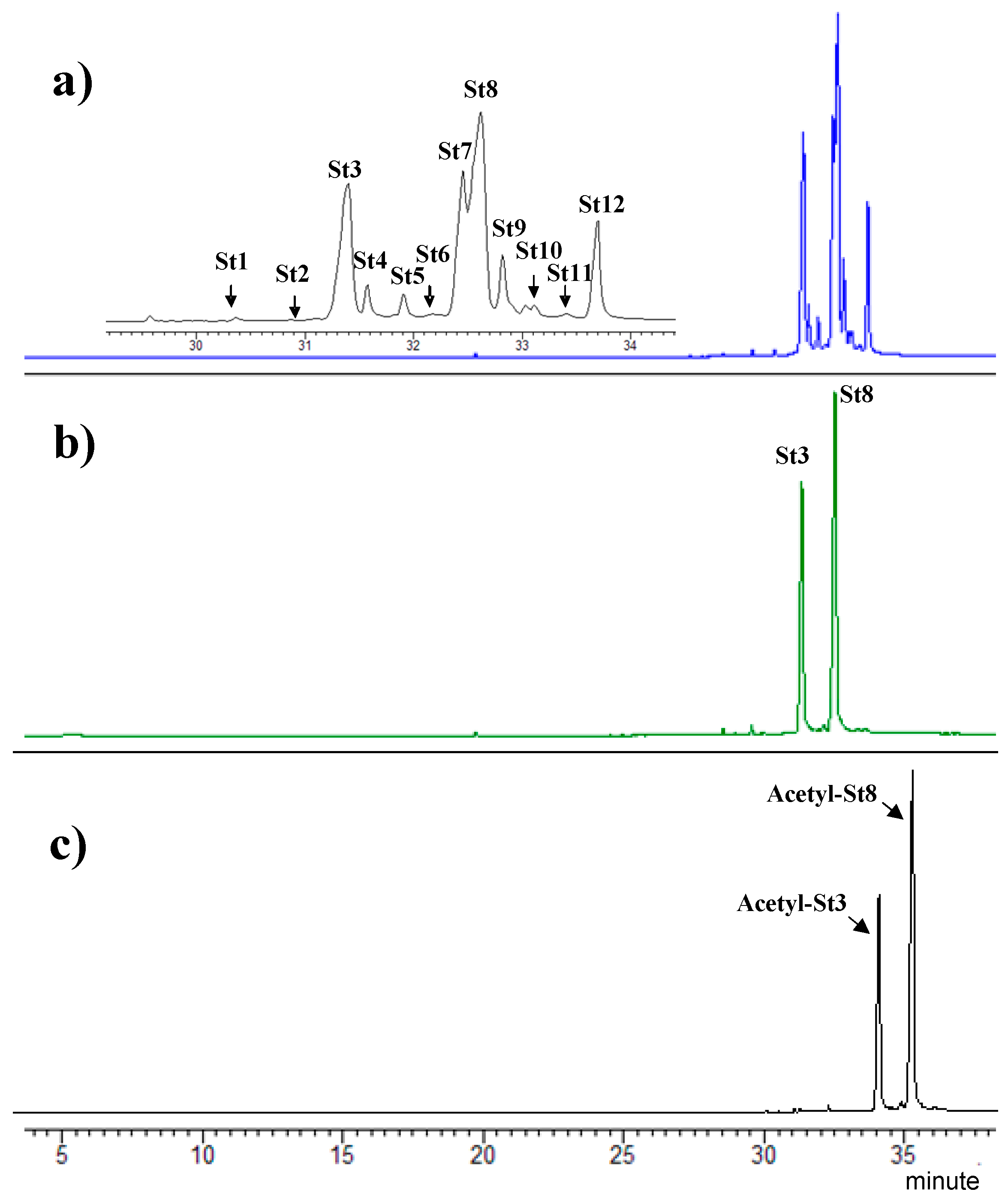
4.1. Extraction and Purification of Total Sterols Fraction, of Ergosterol/7-Dehydroporiferasterol, and of Acetylated Ergosterol/7-Dehydroporiferasterol Mixture from D. tertiolecta
4.2. Analyses by Gas Chromatography-Mass (Tandem) Spectrometry
4.3. Animals and Experimental Treatments
4.4. Isolation of PBMC
4.5. PBMC for Lymphocyte Stimulation Assay and Cytokine Determination
4.6. Determination of Interleukins in Culture Supernatant by ELISA Testing
4.7. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Owiny, J.R.; Gilbert, R.O.; Wahl, C.H.; Nathanielsz, P.W. Leukocytic invasion of the ovine cervix at parturition. J. Soc. Gynecol. Investig. 1995, 2, 593–596. [Google Scholar] [CrossRef]
- Goligthly, E.; Jobbour, H.N.; Normana, J.E. Endocrine immune interactions in human parturition. Mol. Cell Endocrinol. 2011, 335, 52–59. [Google Scholar] [CrossRef] [PubMed]
- Osman, I.; Young, A.; Ledingham, M.A.; Thomson, A.J.; Jordan, F.; Greer, I.A.; Norman, J.E. Leukocyte density and pro-inflammatory cytokine expression in human fetal membranes, decidua, cervix and myometrium before and during labour at term. Mol. Hum. Reprod. 2003, 9, 41–45. [Google Scholar] [CrossRef] [PubMed]
- Lewis, G.S. Uterine health and disorders. J. Dairy Sci. 1997, 80, 984–994. [Google Scholar] [CrossRef]
- Mallard, B.A.; McBride, B.W.; Kehrli, M.E.; Coussens, P.M. Bovine immunophysiology and gentetics: A review of the research and career of Jeanne L. Burton. Vet. Immunol. Immunopathol. 2009, 128, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Islam, R.; Kumar, H.; Nandi, S.; Rai, R.B. Determination of anti-inflammatory cytokine in periparturient cows for prediction of postpartum reproductive diseases. Theriogenology 2013, 79, 974–979. [Google Scholar] [CrossRef] [PubMed]
- Sordillo, L.M.; Raphael, W. Significance of metabolic stress, lipid mobilization, and inflammation on transition cow disorders. Vet. Clin. Food Anim. 2013, 29, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Polynsaturated fatty acids and inflammatory processes: New twist in an old tale. Biochimie 2009, 91, 791–795. [Google Scholar] [CrossRef] [PubMed]
- Bradford, B.; Yuan, K.; Farney, J.K.; Mamedova, L.K.; Carpernter, A.J. Invited review: Inflammation during the transition to lactation: New adventures with an old flame. J. Dairy Sci. 2015, 98, 6631–6650. [Google Scholar] [CrossRef] [PubMed]
- Contreras, G.A.; O’Boyle, N.J.; Herdt, T.H.; Sordillo, L.M. Lipomobilization in periparturient dairy cows influences the composition of plasma nonesterified fatty acids and leukocyte phospholipid fatty acids. J. Dairy Sci. 2010, 93, 2508–2516. [Google Scholar] [CrossRef] [PubMed]
- Lock, A.L.; Preseault, C.L.; Dann, H.M. Exploring the potential for using erythrocyte membranes in the assessment of long-chain polyunsaturated fatty acid status of dairy cows. In Ruminant Physiology: Digestion, Metabolism, and Effect of Nutrition on Reproduction and Welfare; Wageningen Academic Publishers: Wageningen, The Netherlands, 2009; pp. 584–585. [Google Scholar]
- Contreras, G.A.; Mattmiller, S.A.; Raphael, W.; Gandy, J.C.; Sordillo, L.M. Enhanced n-3 phospholipid content reduces inflammatory responses in bovine endothelial cells. J. Dairy Sci. 2012, 95, 7137–7150. [Google Scholar] [CrossRef] [PubMed]
- Wattegedera, S.; Rocchi, M.; Sales, J.; Howard, C.J.; Hope, J.C.; Entrican, G. Antigen-specific peripheral immune responses are unaltered during normal pregnancy in sheep. Am. J. Reprod. 2008, 77, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Caroprese, M.; Ciliberti, M.G.; Albenzio, M.; Annicchiarico, G.; Sevi, A. Dietary polyunsaturated fatty acids from flaxseed affect immune responses of dairy sheep around parturition. Vet. Immunol. Immunopathol. 2015, 168, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Francavilla, M.; Trotta, P.; Luque, R. Phytosterols from Dunaliella tertiolecta and Dunaliella salina: A potentially novel industrial application. Biores. Technol. 2010, 101, 4144–4150. [Google Scholar] [CrossRef] [PubMed]
- Caroprese, M.; Albenzio, M.; Ciliberti, M.G.; Francavilla, M.; Sevi, A. A mixture of phytosterols from Dunaliella tertiolecta affects proliferation of peripheral blood mononuclear cells and cytokine production in sheep. Vet. Immunol. Immunopathol. 2012, 150, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Vane, J.R.; Botting, R.M. New insights into the mode of action of anti-inflammatory drugs. Inflamm. Res. 1995, 44, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sordillo, L.M.; Contreras, G.A.; Aitken, S.L. Metabolic factors affecting the inflammatory response of periparturient dairy cows. Anim. Health Res. Rev. 2009, 10, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Foldi, J.; Kulcsar, M.; Pecsi, A.; Huyghe, B.; de Sa, C.; Lohuis, J.A.C.M.; Cox, P.; Huszenicza, G. Bacterial complications of postpartum uterine involution in cattle. Anim. Reprod. Sci. 2006, 96, 265–281. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, I.M.; Lewis, G.S.; LeBlanc, S.J.; Gilbert, R.O. Defining postpartum uterine disease in cattle. Theriogenology 2006, 65, 1516–1530. [Google Scholar] [CrossRef] [PubMed]
- Allen, M.S.; Bradford, B.J.; Harvatine, K.J. The cow as a model to study food intake regulation. Annu. Rev. Nutr. 2005, 25, 523–547. [Google Scholar] [CrossRef] [PubMed]
- Grummer, R.R.; Mashek, D.G.; Hayirli, A. Dry matter intake and energy balance in the transition period. Vet. Clin. N. Am. Food Anim. Pract. 2004, 20, 447–470. [Google Scholar] [CrossRef] [PubMed]
- Contreras, G.A.; Sordillo, L.M. Lipid mobilization and inflammatory responses during the transition period of dairy cows. Comp. Immunol. Microbiol. Infect. Dis. 2011, 34, 281–289. [Google Scholar] [CrossRef] [PubMed]
- Spite, M.; Claria, J.; Serhan, C.N. Resolvins, specialized proresolving lipid mediators, and their potential roles in metabolic diseases. Cell Metab. 2014, 19, 21–36. [Google Scholar] [CrossRef] [PubMed]
- Hunter, C.A.; Jones, S.A. IL-6 a keystone cytokine in health and disease. Nat. Immunol. 2015, 16, 448–457. [Google Scholar] [CrossRef] [PubMed]
- Stumhofer, J.S.; Silver, J.S.; Laurence, A.; Porrett, P.M.; Harris, T.H.; Turka, L.A.; Ernst, M.; Saris, C.J.; O’Shea, J.J.; Hunter, C.A. Interleukin 27 and 6 induce STAT3-mediated T cell production of interleukin 10. Nat. Immunol. 2007, 8, 1363–1371. [Google Scholar] [CrossRef] [PubMed]
- McGeachy, M.J.; Bak-Jensen, K.S.; Chen, Y.; Cua, D.J. TGF-β and IL-6 drive the production of IL-17 and IL-10 by T cells and restrain Th-17 cell-mediated pathology. Nat. Immunol. 2007, 8, 1390–1397. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.R.; Urakaze, M.; Huang, R.; Taki, H.; Sugiyama, E.; Knoell, C.T.; Xu, L.; Yeh, E.T.; Auron, P.E. Dietary marine lipids suppress continuous expression of interleukin-1 beta gene transcription. Lipids 1996, 31, S23–S31. [Google Scholar] [CrossRef] [PubMed]
- Bannerman, D.D. Pathogen-dependent induction of cytokines and other soluble inflammatory mediators during intramammary infection of dairy cows. J. Anim. Sci. 2009, 87, 10–25. [Google Scholar] [CrossRef] [PubMed]
- Hansen, P.J. Regulation of uterine immune function by progesterone-lessons from the sheep. J. Reprod. Immunol. 1998, 40, 63–79. [Google Scholar] [CrossRef]
- Shanura Fernando, I.P.; Nah, J.W.; Jeon, Y.J. Potential anti-inflammatory natural products from marine algae. Environ. Toxicol. Pharmacol. 2016, 48, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Suarez, Y.; Fernandez, C.; Ledo, B.; Martin, M.; Gomez-Corodado, D.; Lasuncion, M.A. Sterol stringency of proliferation and cell cycle progression in human cells. Biochim. Biophys. Acta 2005, 1734, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhang, C.Y.; Xu, D.M.; Huang, G.Q.; Xu, Y.L.; Wang, Z.Y.; Fang, S.D.; Chen, Y.; Gu, Y.L. The antiatherogenic effects of components isolated from pollen Typhae. Thromb. Res. 1990, 57, 957–966. [Google Scholar] [CrossRef]
- Yasukawa, K.; Akihisa, T.; Kanno, H.; Kaminaga, T.; Izumida, M.; Sakoh, T.; Tamura, T.; Takido, M. Inhibitory effects of sterols isolated from Chlorella vulgaris on 12-Otetradecanoylphorbol-13-acetate-induced inflammation and tumor promotion in mouse skin. Biol. Pharm. Bull. 1996, 19, 573–576. [Google Scholar] [CrossRef] [PubMed]
- Kobori, M.; Yoshida, M.; Ohnishi-Kameyama, M.; Shinmoto, H. Ergosterol peroxide from edible mushroom suppresses inflammatory responses in RAW 264.7 macrophages and growth of HT29 colon adenocarcinoma cells. Br. J. Pharmacol. 2007, 150, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Wilkins, P.; Hu, W.; Murthy, K.S.; Chen, J.; Lee, Z.; Oyesanya, R.; Wu, J.; Barbour, S.E.; Fang, X. Inhibition of calcium-independent phospholipase A2 suppresses proliferation and tumorigenicity of ovarian carcinoma cells. Biochem. J. 2007, 406, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Francavilla, M.; Colaianna, M.; Zotti, M.; Morgese, M.G.; Trotta, P.; Tucci, P.; Schiavone, S.; Trabace, L. Extraction, Characterization and In Vivo Neuromodulatory Activity of Phytosterols from Microalga Dunaliella tertiolecta. Curr. Med. Chem. 2012, 19, 3058–3067. [Google Scholar] [CrossRef] [PubMed]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Phys. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Meshram, G.A.; Patil, V.D. Simple and Efficient Method for Acetylation of Alcohols, Phenols, Amines, and Thiols Using Anhydrous NiCl2 under Solvent-Free Conditions. Synth. Commun. 2009, 39, 2516–2528. [Google Scholar] [CrossRef]
- Gealt, M.A.; Adler, J.H.; Nes, W.R. The sterols and fatty acids from purified Xagella of Chlamydomonas reinhardi. Lipids 1981, 16, 133–136. [Google Scholar] [CrossRef]
- Patterson, G.W.; Gladu, P.K.; Wikfors, G.H.; Lusby, W.R. Unusual tetraen sterols in some phytoplankton. Lipids 1992, 27, 154–156. [Google Scholar] [CrossRef]
- Chu, F.L.E.; Lund, E.D.; Littreal, P.R.; Ruck, K.E.; Harvey, E.; Le Coz, J.R.; Marty, Y.; Moal, J.; Soudant, P. Sterol production and phytosterols bioconversion in two species of heterotrophic protists, Oxyrrhis marina and Gyrodinium dominans. Mar. Biol. 2008, 156, 155–169. [Google Scholar] [CrossRef]
- AOAC 2000 Official Methods of Analysis, 17th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 2000.
- O’Fallon, J.V.; Busboom, J.R.; Nelson, M.L.; Gaskins, C.T. A direct method for fatty acid methyl ester synthesis: Application to wet meat tissues, oils, and feedstuffs. J. Anim. Sci. 2007, 85, 1511–1521. [Google Scholar] [CrossRef] [PubMed]
- Wattegedera, S.; Sills, K.; Howard, C.J.; Hope, J.C.; McInnes, C.J.; Entrican, G. Variability in cytokine production and cell proliferation by mitogen-activate ovine peripheral blood mononuclear cells: Modulation by interleukin (IL)-10 and IL-12. Vet. Immunol. Immunopathol. 2004, 102, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Caroprese, M.; Albenzio, M.; Annicchiarico, G.; Sevi, A. Changes occurring in immune responsiveness of single- and twin bearing Comisana ewes during the transition period. J. Dairy Sci. 2006, 89, 562–568. [Google Scholar] [CrossRef]
- Kwong, L.S.; Hope, J.C.; Thom, M.L.; Sopp, P.; Duggan, S.; Bembridge, G.P.; Howard, C.J. Development of an ELISA for bovine IL-10. Vet. Immunol. Immunopathol. 2002, 85, 213–223. [Google Scholar] [CrossRef]
- Hope, J.C.; Whelan, A.O.; Hewinson, R.G.; Vordermeier, M.; Howard, C.J. Maturation of bovine dentritic cells by lipopeptides. Vet. Immunol. Immunopathol. 2003, 95, 21–31. [Google Scholar] [CrossRef]
- EU Directive 2010/63/EU of 22 September 2010 on the protection of animals used for scientific purposes, 2010. Official Journal of the European Union. Available online: http://eur-lex.europa.eu/LexUriServ/LexUriServ.do?uri=OJ:L:2010:276:0033:0079:en:PDF (accessed on 3 July 2017).
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality. Biometrika 1965, 52, 591–601. [Google Scholar] [CrossRef]
- SAS Institute. SAS Enterprise Guide: Statistics, version 6.1; SAS Institute Inc.: Cary, NC, USA, 2013. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diet | Days Post Partum | ||||
|---|---|---|---|---|---|
| 0 | 7 | 14 | 21 | 28 | |
| CON | 0.097 ± 0.006 Ab | 0.115 ± 0.007 Aa | 0.092 ± 0.006 A | 0.058 ± 0.007 B | 0.034 ± 0.007 Cb |
| FS | 0.130 ± 0.008 Aa | 0.078 ± 0.007 Bb | 0.080 ± 0.006 B | 0.048 ± 0.007 C | 0.067 ± 0.007 ABa |
| Cytokines | Diet | PBMC Treatment | SEM | Effect, p | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| NC | SC | TS | TS | PE | PE | AcPE | AcPE | ||||||
| 0.2 mg/mL | 0.4 mg/mL | 0.2 mg/mL | 0.4 mg/mL | 0.2 mg/mL | 0.4 mg/mL | DT | TM | PS | |||||
| IL-10 | CON | 0.347 | 0.322 | 0.277 | 0.314 | 0.238 | 0.233 | 0.256 | 0.266 | ||||
| FS | 0.439 | 0.473 | 0.468 | 0.568 | 0.492 | 0.401 | 0.348 | 0.353 | 0.06 | *** | NS | NS | |
| IL-6 | CON | 64.498 | 58.967 | 59.832 | 63.179 | 62.576 | 60.681 | 62.050 | 66.274 | ||||
| FS | 99.183 | 101.890 | 102.170 | 101.170 | 105.610 | 103.940 | 105.120 | 104.900 | 5.23 | *** | NS | NS | |
| IL-1β | CON | 1.462 | 1.443 | 1.995 | 1.498 | 2.034 | 1.449 | 1.747 | 1.614 | ||||
| FS | 0.761 | 0.873 | 1.059 | 0.945 | 0.927 | 0.891 | 1.008 | 0.920 | 0.15 | *** | * | * | |
| Fatty Acids, g/100 g of Total Fatty Acids | Concentrate 1 | Oat Hay | Flaxseed 2 |
|---|---|---|---|
| C14:0 | 16.09 | 5.62 | 29.04 |
| C16:0 | 2.64 | 3.67 | 5.41 |
| C18:0 | 19.05 | 19.50 | 20.85 |
| C18:1cis-9 | 53.73 | 16.15 | 20.84 |
| C18:2cis-9cis-12 | 6.87 | 54.03 | 11.79 |
| C18:3n3 | 1.77 | 1.18 | 12.07 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ciliberti, M.G.; Francavilla, M.; Intini, S.; Albenzio, M.; Marino, R.; Santillo, A.; Caroprese, M. Phytosterols from Dunaliella tertiolecta Reduce Cell Proliferation in Sheep Fed Flaxseed during Post Partum. Mar. Drugs 2017, 15, 216. https://doi.org/10.3390/md15070216
Ciliberti MG, Francavilla M, Intini S, Albenzio M, Marino R, Santillo A, Caroprese M. Phytosterols from Dunaliella tertiolecta Reduce Cell Proliferation in Sheep Fed Flaxseed during Post Partum. Marine Drugs. 2017; 15(7):216. https://doi.org/10.3390/md15070216
Chicago/Turabian StyleCiliberti, Maria Giovanna, Matteo Francavilla, Simona Intini, Marzia Albenzio, Rosaria Marino, Antonella Santillo, and Mariangela Caroprese. 2017. "Phytosterols from Dunaliella tertiolecta Reduce Cell Proliferation in Sheep Fed Flaxseed during Post Partum" Marine Drugs 15, no. 7: 216. https://doi.org/10.3390/md15070216
APA StyleCiliberti, M. G., Francavilla, M., Intini, S., Albenzio, M., Marino, R., Santillo, A., & Caroprese, M. (2017). Phytosterols from Dunaliella tertiolecta Reduce Cell Proliferation in Sheep Fed Flaxseed during Post Partum. Marine Drugs, 15(7), 216. https://doi.org/10.3390/md15070216