The Biological Activities of Sesterterpenoid-Type Ophiobolins



Abstract
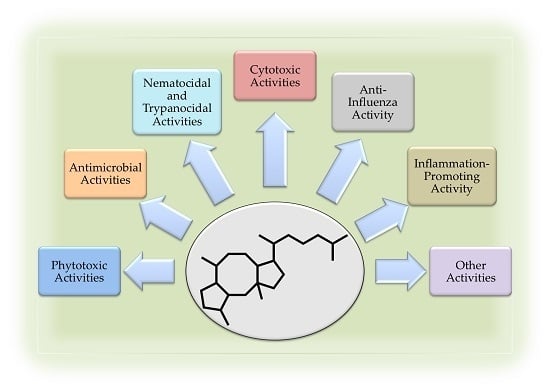
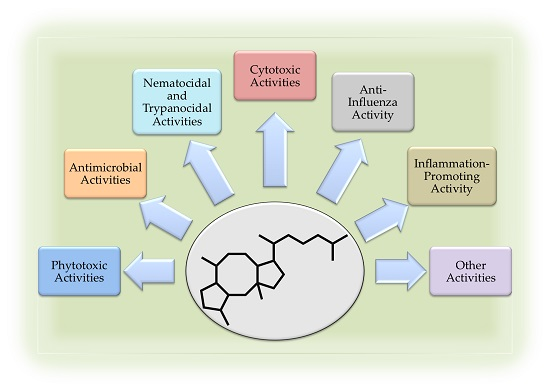
:
1. Introduction
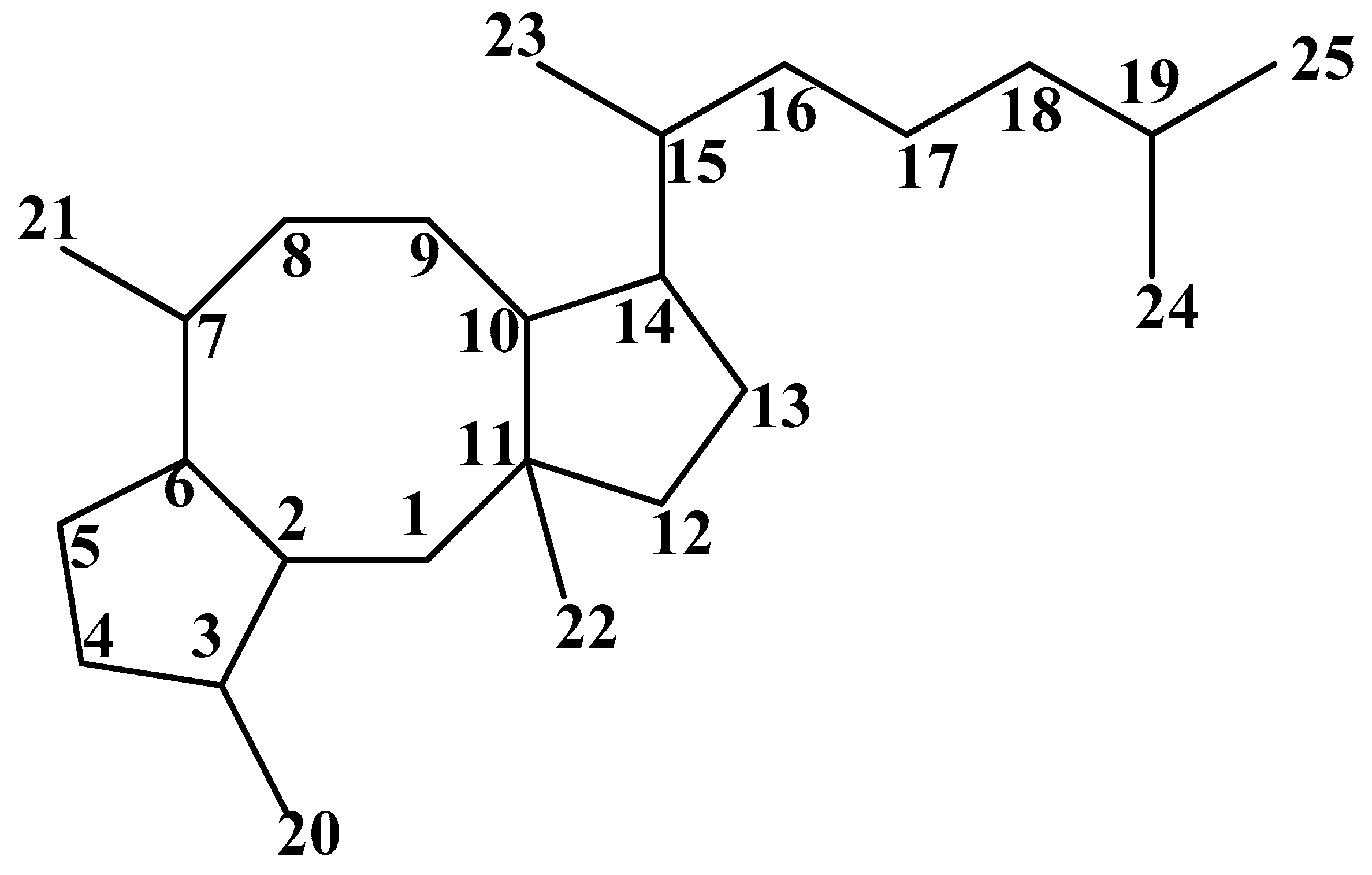
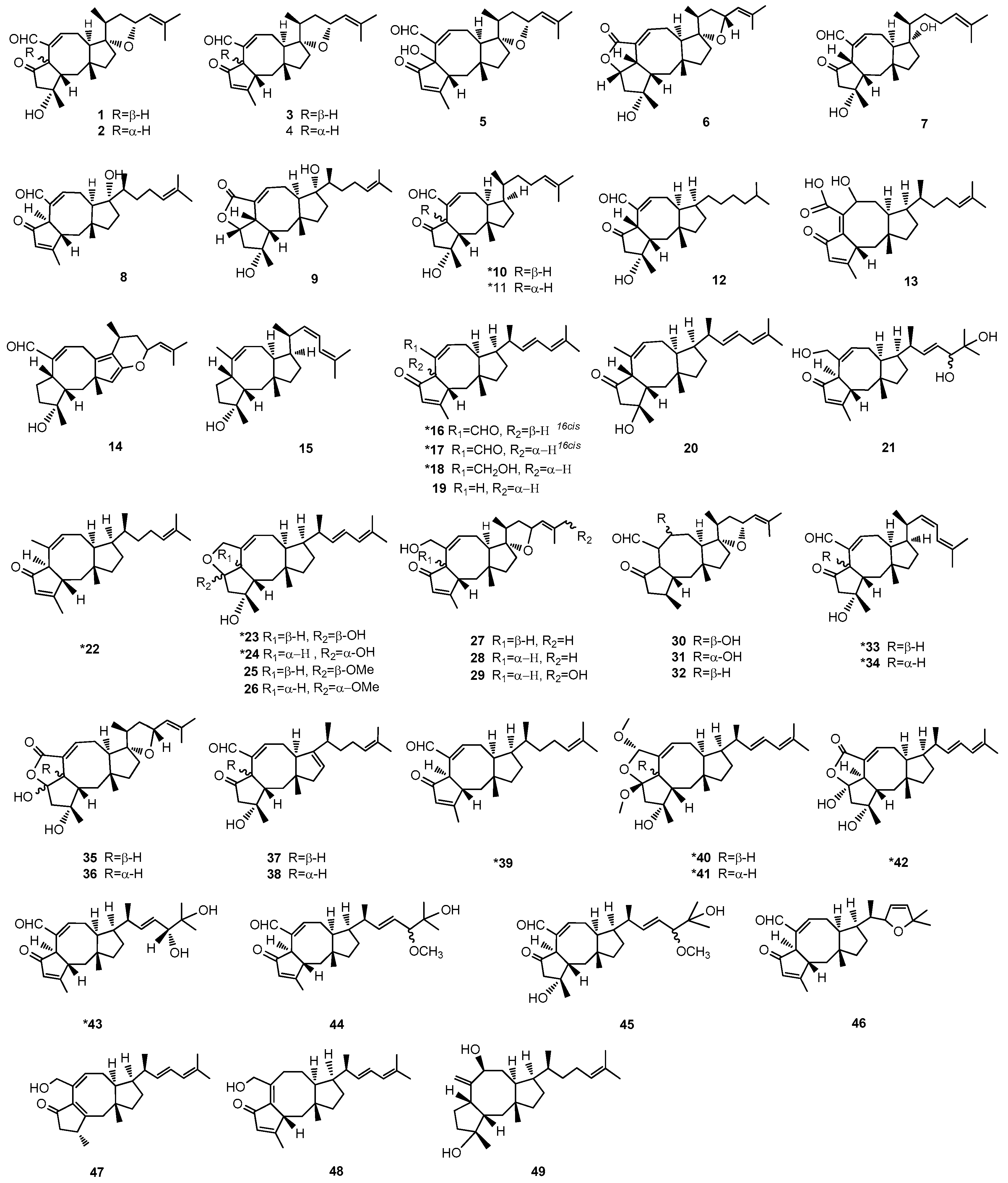
2. Chemical Structure and Source of Ophs
3. Phytotoxic Activities
4. Antimicrobial Activities
4.1. Antibacterial Activity
4.2. Antifungal Activity
5. Nematocidal and Trypanocidal Activities
6. Cytotoxic Activities
6.1. Cytotoxic Activity In Vitro
6.2. Cytotoxic Activity In Vivo
6.3. Structure–Cytotoxic Activity Relationship
6.3.1. Stereochemistry at C6
6.3.2. Hydroxy Group at C3, C6, C14
6.3.3. Tetrahydrofuran Ring
6.3.4. Other Structure–Cytotoxic Activity Relationship
6.4. Mechanism of Cytotoxic Activities on Cancer Cells
6.4.1. Presumed Protein and Non-Protein Targets
6.4.2. Pathways Related to Ophs Cytotoxicity
6.4.3. Cell Death Phenotypes
7. Anti-Influenza Activity
8. Inflammation-Promoting Activity
9. Other Activities
10. Conclusions and Future Perspectives
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Evidente, A.; Kornienko, A.; Cimmino, A.; Andolfi, A.; Lefranc, F.; Mathieu, V.; Kiss, R. Fungal metabolites with anticancer activity. Nat. Prod. Rep. 2014, 31, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Ebada, S.S.; Proksch, P. Marine-derived fungal metabolites. In Springer Handbook of Marine Biotechnology; Kim, S.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 759–788. [Google Scholar]
- Tommonaro, G.; Rosa, S.D.; Carnuccio, R.; Maiuri, M.C.; Stefano, D.D. Marine sponge sesterpenoids as potent apoptosis-inducing factors in human carcinoma cell lines. In Handbook of Anticancer Drugs from Marine Origin; Kim, S.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 439–479. [Google Scholar]
- Evidente, A.; Kornienko, A.; Lefranc, F.; Cimmino, A.; Dasari, R.; Evidente, M.; Mathieu, V.; Kiss, R. Sesterterpenoids with anticancer activity. Curr. Med. Chem. 2015, 22, 3502–3522. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yang, B.; Lin, X.P.; Zhou, X.F.; Liu, Y. Sesterterpenoids. Nat. Prod. Rep. 2013, 30, 455–473. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Dong, M.; Wang, H.; Crews, P. Secondary metabolites from the marine sponge genus Phyllospongia. Mar. Drugs 2017, 15, 12. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Fukuzawa, S.; Satake, M.; Li, X.; Kuranaga, T.; Niitsu, A.; Yoshizawa, K.; Tachibana, K. Ophiobolin O and 6-epi-ophiobolin O, two new cytotoxic sesterterpenes from the marine derived fungus Aspergillus sp. Nat. Prod. Commun. 2012, 7, 1411–1414. [Google Scholar] [PubMed]
- Nakamura, M.; Ishibashi, K. On the new antibiotics “Ophiobolin”, produced by Ophiobolus miyabeanus. J. Agric. Chem. Soc. Jpn. 1958, 32, 739–744. [Google Scholar]
- Au, T.K.; Chick, W.S.; Leung, P.C. The biology of ophiobolins. Life Sci. 2000, 67, 733–742. [Google Scholar] [CrossRef]
- Tipton, C.L.; Paulsen, P.V.; Betts, R.E. Effects of ophiobolin a on ion leakage and hexose uptake by maize roots. Plant Physiol. 1977, 59, 907–910. [Google Scholar] [CrossRef] [PubMed]
- Ohkawa, H.; Tamura, T. Studies on the metabolites of Cochliobolus miyabeanus. Agric. Biol. Chem. 1966, 30, 285–291. [Google Scholar]
- Evidente, A.; Andolfi, A.; Cimmino, A.; Vurro, M.; Fracchiolla, M.; Charudattan, R. Herbicidal potential of ophiobolins produced by Drechslera gigantea. J. Agric. Food Chem. 2006, 54, 1779–1783. [Google Scholar] [CrossRef] [PubMed]
- Sugawara, F.; Strobel, G.; Strange, R.N.; Siedow, J.N.; Van Duyne, G.D.; Clardy, J. Phytotoxins from the pathogenic fungi Drechslera maydis and Drechslera sorghicola. Proc. Natl. Acad. Sci. USA 1987, 84, 3081–3085. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.Z.; Tsuda, M.; Doke, N.; Nishimura, S. Phytotoxins produced by germinating spores of Bipolaris oryzae. Phytopathology 1991, 81, 58–64. [Google Scholar] [CrossRef]
- Kim, H.J.; Kim, J.C.; Kim, B.S.; Kim, H.G.; Cho, K.Y. Antibiotic and phytotoxic activities of Ophiobolins from Helminthosporium species. Plant Pathol. J. 1999, 15, 14–20. [Google Scholar]
- Mandal, S.M.; Roy, A.; Paul, D.; Korpole, S.; Shrivastava, S.L.; Chakraborty, R.; Basak, A. Fungi fights fungi: Tip-off in antifungal chemotherapy. In Recent Trends in Antifungal Agents and Antifungal Therapy; Basak, A., Chakraborty, R., Mandal, S.M., Eds.; Springer: Delhi, India, 2016; pp. 1–27. [Google Scholar]
- Li, G.H.; Zhang, K.Q. Nematode-toxic fungi and their nematicidal metabolites. In Nematode-Trapping Fungi; Zhang, K.Q., Hyde, K.D., Eds.; Fungal Diversity Research Series; Springe: Berlin, Germany, 2014; pp. 313–375. [Google Scholar]
- Tsipouras, A.; Adefarati, A.A.; Tkacz, J.S.; Frazier, E.G.; Rohrer, S.P.; Birzin, E.; Rosegay, A.; Zink, D.L.; Goetz, M.A.; Singh, S.B.; et al. Ophiobolin M and analogues, noncompetitive inhibitors of ivermectin binding with nematocidal activity. Bioorg. Med. Chem. 1996, 4, 531–536. [Google Scholar] [CrossRef]
- De Carvalho, C.R.; Vieira, M.D.L.A.; Cantrell, C.L.; Wedge, D.E.; Alves, T.M.A.; Zani, C.L.; Pimenta, R.S.; Sales Junior, P.A.; Murta, S.M.F.; Romanha, A.J.; et al. Biological activities of ophiobolin K and 6-epi-ophiobolin K produced by the endophytic fungus Aspergillus calidoustus. Nat. Prod. Res. 2016, 30, 478–481. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Lv, C.; Zhu, T.; Yang, X.; Wei, S.; Sun, J.; Hong, K.; Zhu, W.; Huang, C. Ophiobolin-O reverses adriamycin resistance via cell cycle arrest and apoptosis sensitization in adriamycin-resistant human breast carcinoma (MCF-7/ADR) cells. Mar. Drugs 2013, 11, 4570–4584. [Google Scholar] [CrossRef] [PubMed]
- Bury, M.; Novo-Uzal, E.; Andolfi, A.; Cimini, S.; Wauthoz, N.; Heffeter, P.; Lallemand, B.; Avolio, F.; Delporte, C.; Cimmino, A.; et al. Ophiobolin A, a sesterterpenoid fungal phytotoxin, displays higher in vitro growth-inhibitory effects in mammalian than in plant cells and displays in vivo antitumor activity. Int. J. Oncol. 2013, 43, 575–585. [Google Scholar] [PubMed]
- Wang, S.; Luo, X.; Yan, R.; Wang, Q.; Qi, Q.; Chi, X.; Zhang, L.; Yu, Z.; Cai, B.; Chen, J.L.; et al. 3-Anhydro-6-hydroxy-ophiobolin A displays high in vitro and in vivo efficacy against influenza A virus infection. Protein Cell 2016, 7, 839–843. [Google Scholar] [CrossRef] [PubMed]
- Pósa, A.; Szabó, R.; Szalai, Z.; Kupai, K.; Deim, Z.; Murlasits, Z.; Bencsik, O.; Szekeres, A.; Vágvölgyi, C.; Balogh, L.; et al. The effect of acute ophiobolin A treatment on HO-mediated inflammatory processes. Hum. Exp. Toxicol. 2016, 36, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Bladt, T.T.; Dürr, C.; Knudsen, P.B.; Kildgaard, S.; Frisvad, J.C.; Gotfredsen, C.H.; Seiffert, M.; Larsen, T.O. Bio-activity and dereplication-based discovery of ophiobolins and other fungal secondary metabolites targeting leukemia cells. Molecules 2013, 18, 14629–14650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kildgaard, S.; Mansson, M.; Dosen, I.; Klitgaard, A.; Frisvad, J.C.; Larsen, T.O.; Nielsen, K.F. Accurate dereplication of bioactive secondary metabolites from marine-derived fungi by UHPLC-DAD-QTOFMS and a MS/HRMS library. Mar. Drugs 2014, 12, 3681–3705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, H.; Itoh, T.; Kinoshita, M.; Nakai, Y.; Kurotaki, M.; Kobayashi, M. Cytotoxic sesterterpenes, 6-epi-ophiobolin G and 6-epi-ophiobolin N, from marine derived fungus Emericella variecolor GF10. Tetrahedron 2004, 60, 6015–6019. [Google Scholar] [CrossRef]
- Arai, M.; Niikawa, H.; Kobayashi, M. Marine-derived fungal sesterterpenes, ophiobolins, inhibit biofilm formation of Mycobacterium species. J. Nat. Med. 2013, 67, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Evidente, A.; Andolfi, A.; Cimmino, A.; Vurro, M.; Fracchiolla, M.; Charudattan, R.; Motta, A. Ophiobolin E and 8-epi-ophiobolin J produced by Drechslera gigantea, a potential mycoherbicide of weedy grasses. Phytochemistry 2006, 67, 2281–2287. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.X.; Bao, L.; Yang, X.L.; Liu, D.L.; Guo, H.; Dai, H.Q.; Song, F.H.; Zhang, L.X.; Guo, L.D.; Li, S.J.; et al. Ophiobolins P–T, five new cytotoxic and antibacterial sesterterpenes from the endolichenic fungus Ulocladium sp. Fitoterapia 2013, 90, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Li, E.; Clark, A.M.; Rotella, D.P.; Hufford, C.D. Microbial metabolites of ophiobolin A and antimicrobial evaluation of ophiobolins. J. Nat. Prod. 1995, 58, 74–81. [Google Scholar] [CrossRef] [PubMed]
- Jayasuriya, H.; Herath, K.B.; Ondeyka, J.G.; Polishook, J.D.; Bills, G.F.; Dombrowski, A.W.; Springer, M.S.; Siciliano, S.; Malkowitz, L.; Sanchez, M.; et al. Isolation and structure of antagonists of chemokine receptor (CCR5). J. Nat. Prod. 2004, 67, 1036–1038. [Google Scholar] [CrossRef] [PubMed]
- Itai, A.; Nozoe, S. Structure and chemistry of some ophiobolin D derivatives. Chem. Pharm. Bull. 1980, 28, 1043–1050. [Google Scholar] [CrossRef]
- Phuwapraisirisan, P.; Sawang, K.; Siripong, P.; Tip-pyang, S. Anhydrocochlioquinone A, a new antitumor compound from Bipolaris oryzae. Tetrahedron Lett. 2007, 38, 5193–5195. [Google Scholar] [CrossRef]
- Sugawara, F.; Takahashi, N.; Strobel, G.; Yun, C.H.; George, G.; Fu, Y.; Clardy, J. Some new phytotoxic ophiobolins produced by Drechslera oryzae. J. Org. Chem. 1988, 53, 2170–2172. [Google Scholar] [CrossRef]
- Shen, X.; Krasnoff, S.B.; Lu, S.W.; Dunbar, C.D.; O’Neal, J.; Turgeon, B.G.; Yoder, O.C.; Gibson, D.M.; Hamann, M.T. Characterization of 6-epi-3-anhydroophiobolin B from Cochliobolus heterostrophus. J. Nat. Prod. 1999, 62, 895–897. [Google Scholar] [CrossRef] [PubMed]
- Ahn, J.W.; Lee, M.K.; Choi, S.U.; Lee, C.O.; Kim, B.S. Cytotoxic ophiobolins produced by Bipolaris sp. J. Microbiol. Biotechnol. 1998, 8, 406–408. [Google Scholar]
- Zhao, X.L.; Niu, Y.C.; Deng, H.; Luo, D.Q. Secondary metabolites from Bipolaris setariae NY1 strain. Mycosystema 2013, 32, 286–291. [Google Scholar]
- Wang, Q.X.; Yang, J.L.; Qi, Q.Y.; Bao, L.; Yang, X.L.; Liu, M.M.; Huang, P.; Zhang, L.X.; Chen, J.L.; Cai, L.; et al. 3-Anhydro-6-hydroxy-ophiobolin A, a new sesterterpene inhibiting the growth of methicillin-resistant Staphylococcus aureus and inducing the cell death by apoptosis on K562, from the phytopathogenic fungus Bipolaris oryzae. Bioorg. Med. Chem. Lett. 2013, 23, 3547–3550. [Google Scholar] [CrossRef] [PubMed]
- Gianani, L.; Cocucci, S.; Pardi, D.; Randazzo, G. Effects of ophiobolin B on cell enlargement and H+/K+ exchange in maize coleoptile tissues. Planta 1979, 146, 271–274. [Google Scholar] [CrossRef] [PubMed]
- Bladt, T.T.; Frisvad, J.C.; Knudsen, P.B.; Larsen, T.O. Anticancer and antifungal compounds from Aspergillus, Penicillium and other filamentous fungi. Mol. Basel Switz. 2013, 18, 11338–11376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akihiko, K.; Shigeo, N.; Shigenobu, O. Subcellular distribution of sesterterpene- and sterol-biosynthesizing activities in Cochliobolus heterostrophus. Biochim. Biophys. Acta BBA-Lipids Lipid Metab. 1973, 296, 615–623. [Google Scholar] [CrossRef]
- Nozoe, S.; Morisaki, M. Enzymic formation of a tricyclic sesterterpene alcohol from mevalonic acid and all-trans-geranylfarnesyl pyrophosphate. Chem. Commun. 1969, 22, 1319–1320. [Google Scholar] [CrossRef]
- Liu, X.H.; Miao, F.P.; Qiao, M.F.; Cichewicz, R.H.; Ji, N.Y. Terretonin, ophiobolin, and drimane terpenes with absolute configurations from an algicolous Aspergillus ustus. RSC Adv. 2013, 3, 588–595. [Google Scholar] [CrossRef]
- Cutler, H.G.; Crumley, F.G.; Cox, R.H.; Springer, J.P.; Arrendale, R.F.; Cole, R.J.; Cole, P.D. Ophiobolins G and H: New fungal metabolites from a novel source, Aspergillus ustus. J. Agric. Food Chem. 1984, 32, 778–782. [Google Scholar] [CrossRef]
- Liu, H.B.; Edrada-Ebel, R.; Ebel, R.; Wang, Y.; Schulz, B.; Draeger, S.; Müller, W.E.G.; Wray, V.; Lin, W.H.; Proksch, P. Ophiobolin sesterterpenoids and pyrrolidine alkaloids from the sponge-derived fungus Aspergillus ustus. Helv. Chim. Acta 2011, 94, 623–631. [Google Scholar] [CrossRef]
- Zhu, W.M.; Lu, Z.Y.; Hong, K.; Miao, C. The Preparation and Application of Sesterterpenoids Compounds Ophiobolins. CN Patent 101591314, 2 December 2009. [Google Scholar]
- Proksch, P.; Ebel, R.; Edrada, R.; Riebe, F.; Liu, H.; Diesel, A.; Bayer, M.; Li, X.; Lin, W.H.; Grebenyuk, V.; et al. Sponge-associated fungi and their bioactive compounds: The suberites case. Bot. Mar. 2008, 51, 209–218. [Google Scholar] [CrossRef]
- Singh, S.B.; Smith, J.L.; Sabnis, G.S.; Dombrowski, A.W.; Schaeffer, J.M.; Goetz, M.A.; Bills, G.F. Structure and conformation of ophiobolin K and 6-epi-ophiobolin K from Aspergillus ustus as a nematocidal agent. Tetrahedron 1991, 47, 6931–6938. [Google Scholar] [CrossRef]
- Grunewald, S.; Kriegel, C.; Baumann, T.; Glander, H.J.; Paasch, U. Interactions between apoptotic signal transduction and capacitation in human spermatozoa. Hum. Reprod. 2009, 24, 2071–2078. [Google Scholar] [CrossRef] [PubMed]
- Locato, V.; Uzal, E.N.; Cimini, S.; Zonno, M.C.; Evidente, A.; Micera, A.; Foyer, C.H.; Gara, L.D. Low concentrations of the toxin ophiobolin A lead to an arrest of the cell cycle and alter the intracellular partitioning of glutathione between the nuclei and cytoplasm. J. Exp. Bot. 2015, 66, 2991–3000. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.V.K.; Reddy, M.S.; Kloepper, J.W.; Lawrence, K.S.; Zhou, X.G.; Groth, D.E.; Zhang, S.; Rao, R.S.; Wang, Q.; Raju, M.R.B.; et al. Commercial potential of microbial inoculants for sheath blight management and yield enhancement of rice. In Bacteria in Agrobiology: Crop Ecosystems; Maheshwari, D.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 237–264. [Google Scholar]
- Dallagnol, L.J.; Rodrigues, F.A.; Martins, S.C.V.; Cavatte, P.C.; DaMatta, F.M. Alterations on rice leaf physiology during infection by Bipolaris oryzae. Australas. Plant Pathol. 2011, 40, 360–365. [Google Scholar] [CrossRef]
- Chattopadhyay, A.K.; Samaddar, K.R. Effects of Helminthosporium oryzae infection and ophiobolin on the cell membranes of host tissues. Physiol. Plant Pathol. 1976, 8, 131–139. [Google Scholar] [CrossRef]
- Dallagnol, L.J.; Martins, S.C.V.; DaMatta, F.M.; Rodrigues, F.Á. Brown spot negatively affects gas exchange and chlorophyll a fluorescence in rice leaves. Trop. Plant Pathol. 2015, 40, 275–278. [Google Scholar] [CrossRef]
- Leung, P.C.; Taylor, W.A.; Wang, J.H.; Tipton, C.L. Ophiobolin A, A natural product inhibitor of calmodulin. J. Biol. Chem. 1984, 259, 2742–2747. [Google Scholar] [PubMed]
- Leung, P.C.; Graves, L.M.; Tipton, C.L. Characterization of the interaction of ophiobolin A and calmodulin. Int. J. Biochem. 1988, 20, 1351–1359. [Google Scholar] [CrossRef]
- Krizsán, K.; Bencsik, O.; Nyilasi, I.; Galgóczy, L.; Vágvölgyi, C.; Papp, T. Effect of the sesterterpene-type metabolites, ophiobolins A and B, on zygomycetes fungi. FEMS Microbiol. Lett. 2010, 313, 135–140. [Google Scholar] [CrossRef] [PubMed]
- De Vries-van Leeuwen, I.J.; Kortekaas-Thijssen, C.; Nzigou Mandouckou, J.A.; Kas, S.; Evidente, A.; de Boer, A.H. Fusicoccin-A selectively induces apoptosis in tumor cells after interferon-α priming. Cancer Lett. 2010, 293, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Bencsik, O.; Papp, T.; Berta, M.; Zana, A.; Forgó, P.; Dombi, G.; Andersson, M.A.; Salkinoja-Salonen, M.; Vágvölgyi, C.; Szekeres, A. Ophiobolin A from Bipolaris oryzae perturbs motility and membrane integrities of porcine sperm and induces cell death on mammalian somatic cell lines. Toxins 2014, 6, 2857–2871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fujiwara, H.; Matsunaga, K.; Kumagai, H.; Ishizuka, M.; Ohizumi, Y. Ophiobolin A, a novel apoptosis-inducing agent from fungus strain f-7438. Pharm. Pharmacol. Commun. 2000, 6, 427–431. [Google Scholar] [CrossRef]
- Bhatia, D.R.; Dhar, P.; Mutalik, V.; Deshmukh, S.K.; Verekar, S.A.; Desai, D.C.; Kshirsagar, R.; Thiagarajan, P.; Agarwal, V. Anticancer activity of Ophiobolin A, isolated from the endophytic fungus Bipolaris setariae. Nat. Prod. Res. 2016, 30, 1455–1458. [Google Scholar] [CrossRef] [PubMed]
- Bury, M.; Girault, A.; Mégalizzi, V.; Spiegl-Kreinecker, S.; Mathieu, V.; Berger, W.; Evidente, A.; Kornienko, A.; Gailly, P.; Vandier, C.; et al. Ophiobolin A induces paraptosis-like cell death in human glioblastoma cells by decreasing BKCa channel activity. Cell Death Dis. 2013, 4, e561. [Google Scholar] [CrossRef] [PubMed]
- Morrison, R.; Gardiner, C.; Evidente, A.; Kiss, R.; Townley, H. Incorporation of ophiobolin A into novel chemoembolization particles for cancer cell treatment. Pharm. Res. 2014, 31, 2904–2917. [Google Scholar] [CrossRef] [PubMed]
- Morrison, R.; Lodge, T.; Evidente, A.; Kiss, R.; Townley, H. Ophiobolin A, a sesterpenoid fungal phytotoxin, displays different mechanisms of cell death in mammalian cells depending upon the cancer cell origin. Int. J. Oncol. 2017, 50, 773–786. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, N.; Koizumi, M.; Adachi, I.; Kawakami, J. Inhibition of P-glycoprotein-mediated transport by terpenoids contained in herbal medicines and natural products. Food Chem. Toxicol. 2006, 44, 2033–2039. [Google Scholar] [CrossRef] [PubMed]
- Lv, C.; Qin, W.; Zhu, T.; Wei, S.; Hong, K.; Zhu, W.; Chen, R.; Huang, C. Ophiobolin O isolated from Aspergillus ustus induces G1 arrest of MCF-7 cells through interaction with AKT/GSK3β/Cyclin D1 signaling. Mar. Drugs 2015, 13, 431–443. [Google Scholar] [CrossRef] [PubMed]
- Dasari, R.; Masi, M.; Lisy, R.; Ferdérin, M.; English, L.R.; Cimmino, A.; Mathieu, V.; Brenner, A.J.; Kuhn, J.G.; Whitten, S.T.; et al. Fungal metabolite ophiobolin A as a promising anti-glioma agent: In vivo evaluation, structure-activity relationship and unique pyrrolylation of primary amines. Bioorg. Med. Chem. Lett. 2015, 25, 4544–4548. [Google Scholar] [CrossRef] [PubMed]
- Xue, D.; Wang, Q.; Chen, Z.; Cai, L.; Bao, L.; Qi, Q.; Liu, L.; Wang, X.; Jin, H.; Wang, J.; et al. 3-Anhydro-6-hydroxy-ophiobolin A, a fungal sesterterpene from Bipolaris oryzae induced autophagy and promoted the degradation of α-synuclein in PC12 cells. Bioorg. Med. Chem. Lett. 2015, 25, 1464–1470. [Google Scholar] [CrossRef] [PubMed]
- Cabuk, D.; Yetimoglu, E.; Simsek, T.; Gacar, G.; Subasi, C.; Canturk, Z.; Ercin, C.; Karaoz, E.; Uygun, K. The distribution of CD44+/CD24− cancer stem cells in breast cancer and its relationship with prognostic factors. JBUON 2016, 21, 1121–1128. [Google Scholar] [PubMed]
- Camerlingo, R.; Ferraro, G.A.; De Francesco, F.; Romano, M.; Nicoletti, G.; Di Bonito, M.; Rinaldo, M.; D’Andrea, F.; Pirozzi, G. The role of CD44+/CD24−/low biomarker for screening, diagnosis and monitoring of breast cancer. Oncol. Rep. 2014, 31, 1127–1132. [Google Scholar] [PubMed]
- Najumudeen, A.K.; Jaiswal, A.; Lectez, B.; Oetken-Lindholm, C.; Guzmán, C.; Siljamäki, E.; Posada, I.M.D.; Lacey, E.; Aittokallio, T.; Abankwa, D. Cancer stem cell drugs target K-ras signaling in a stemness context. Oncogene 2016, 35, 5248–5262. [Google Scholar] [CrossRef] [PubMed]
- Kong, A.T.; Chow, L.P. Identification of the binding and inhibition sites in the calmodulin molecule for ophiobolin A by site-directed mutagenesis. Plant Physiol. 1998, 118, 965–973. [Google Scholar] [CrossRef]
- Linxweiler, M.; Schorr, S.; Schäuble, N.; Jung, M.; Linxweiler, J.; Langer, F.; Schäfers, H.J.; Cavalié, A.; Zimmermann, R.; Greiner, M. Targeting cell migration and the endoplasmic reticulum stress response with calmodulin antagonists: A clinically tested small molecule phenocopy of SEC62 gene silencing in human tumor cells. BMC Cancer 2013, 5, 574. [Google Scholar] [CrossRef] [PubMed]
- Chidley, C.; Trauger, S.A.; Birsoy, K.; O’Shea, E.K. The anticancer natural product ophiobolin A induces cytotoxicity by covalent modification of phosphatidylethanolamine. Elife 2016, 5, e14601. [Google Scholar] [CrossRef] [PubMed]
- Rodolfo, C.; Rocco, M.; Cattaneo, L.; Tartaglia, M.; Sassi, M.; Aducci, P.; Scaloni, A.; Camoni, L.; Marra, M. Ophiobolin A induces autophagy and activates the mitochondrial pathway of apoptosis in human melanoma cells. PLoS ONE 2016, 11, e0167672. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Lu, Z.; Meng, L.; Wei, S.; Hong, K.; Zhu, W.; Huang, C. The novel agent ophiobolin O induces apoptosis and cell cycle arrest of MCF-7 cells through activation of MAPK signaling pathways. Bioorg. Med. Chem. Lett. 2012, 22, 579–585. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.H.; Zheng, Z.H.; Ma, Y.; Shi, Y.; Dong, Y.S.; Ren, X.; Mu, D.; Zhang, H.; He, J.G. Factor Xa inhibitors from metabolites of microorganisms. Chin. J. Antibiot. 2007, 32, 277–279. [Google Scholar]
- Van Nuffel, A.M.; Sukhatme, V.; Pantziarka, P.; Meheus, L.; Sukhatme, V.P.; Bouche, G. Repurposing drugs in oncology (ReDO)—Clarithromycin as an anti-cancer agent. E Cancer Med. Sci. 2015, 9, 513. [Google Scholar] [CrossRef] [PubMed]
- Rowley, M.; Tsukamoto, M.; Kishi, Y. Total synthesis of (+)-ophiobolin C. J. Am. Chem. Soc. 1989, 111, 2735–2737. [Google Scholar] [CrossRef]
- Ruprah, P.K.; Cros, J.P.; Pease, J.E.; Whittingham, W.G.; Williams, J.M.J. Studies towards the total synthesis of cycloaraneosene and ophiobolin M: A general strategy for the construction of the 5–8 bicyclic ring system. Eur. J. Org. Chem. 2002, 2002, 3145–3152. [Google Scholar] [CrossRef]
- Tsuna, K.; Noguchi, N.; Nakada, M. Convergent total synthesis of (+)-ophiobolin A. Angew. Chem. Int. Ed. 2011, 50, 9452–9455. [Google Scholar] [CrossRef] [PubMed]
- Tsuna, K.; Noguchi, N.; Nakada, M. Enantioselective total synthesis of (+)-ophiobolin A. Chem. Eur. J. 2013, 19, 5476–5486. [Google Scholar] [CrossRef] [PubMed]
- Brill, Z.G.; Grover, H.K.; Maimone, T.J. Enantioselective synthesis of an ophiobolin sesterterpene via a programmed radical cascade. Science 2016, 352, 1078–1082. [Google Scholar] [CrossRef] [PubMed]
- Chiba, R.; Minami, A.; Gomi, K.; Oikawa, H. Identification of ophiobolin F synthase by a genome mining approach: A sesterterpene synthase from aspergillus clavatus. Org. Lett. 2013, 15, 594–597. [Google Scholar] [CrossRef] [PubMed]
- Narita, K.; Chiba, R.; Minami, A.; Kodama, M.; Fujii, I.; Gomi, K.; Oikawa, H. Multiple oxidative modifications in the ophiobolin biosynthesis: P450 oxidations found in genome mining. Org. Lett. 2016, 18, 1980–1983. [Google Scholar] [CrossRef] [PubMed]
- Chai, H.Z.; Yin, R.; Liu, Y.F.; Meng, H.Y.; Zhou, X.Q.; Zhou, G.L.; Bi, X.P.; Yang, X.; Zhu, T.H.; Zhu, W.M.; et al. Sesterterpene ophiobolin biosynthesis involving multiple gene clusters in Aspergillus ustus. Sci. Rep. 2016, 6, 27181. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Number | Compound | Source | Reference | Source | Reference |
|---|---|---|---|---|---|
| 1 | Ophiobolin A | Bipolaris oryzae | [13,14,33] | Drechslera oryzae 1 | [34] |
| Cochliobolus heterostrophus 2 | [30,35] | Drechslera gigantea | [12,28] | ||
| 2 | 6-epi-ophiobolin A | Bipolaris oryzae | [13,14,33] | Drechslera oryzae 1 | [34] |
| Penicillium patulum | [30] | Bipolaris sp. | [36] | ||
| Cochliobolus heterostrophus 2 | [35] | Drechslera gigantea | [12,28] | ||
| 3 | 3-anhydroophiobolin A | Cochliobolus heterostrophus 2 | [30,35] | Bipolaris oryzae | [33] |
| Bipolaris setariae NY1 | [37] | - | - | ||
| 4 | 3-anhydro-6-epi-ophiobolin A | Bipolaris oryzae | [13,14] | Drechslera oryzae 1 | [34] |
| Bipolaris sp. | [36] | Cochliobolus heterostrophus 2 | [35] | ||
| Drechslera gigantea | [12,28] | - | - | ||
| 5 | 3-anhydro-6-hydroxy-Ophiobolin A | Bipolaris oryzae | [38] | - | - |
| 6 | Ophiobolin A lactone | Polyangium cellulosum | [30] | - | - |
| 7 | Ophiobolin B | Helminthosporium oryzae 3 | [39] | Drechslera oryzae 1 | [34] |
| Bipolaris oryzae | [14] | Cochliobolus heterostrophus 2 | [30] | ||
| Drechslera gigantea | [28] | - | - | ||
| 8 | 3-anhydro-6-epi-ophiobolin B | Cochliobolus heterostrophus 2 | [35] | - | - |
| 9 | Ophiobolin B lactone | Pseudomonas aeruginosa | [30] | - | - |
| 10 | Ophiobolin C | Bipolaris oryzae | [13] | Cochliobolus heterostrophus 2 | [18] |
| Mollisia sp. | [31] | Emericella variecolor 4 GF10 | [26] | ||
| Aspergillus insuetus | [24,40] | Aspergillus calidoustus | [24,40] | ||
| 11 | 6-epi-ophiobolin C | Cochliobolus heterostrophus 2 | [18] | Emericella variecolor 4 GF10 | [26] |
| 12 | 18-dihydroophiobolin C | Cochliobolus heterostrophus 2 | [18] | - | - |
| 13 | Ophiobolin D | Cephalosporium caerulens | [32] | - | - |
| 14 | Ophiobolin E | Drechslera gigantea | [28] | - | - |
| 15 | Ophiobolin F | Cochliobolus heterostrophus 2 | [41,42] | Aspergillus ustus cf-42 | [43] |
| 16 | Ophiobolin G | Aspergillus ustus | [44] | Emericella variecolor 4 GF10 | [26] |
| Aspergillus sp. | [7] | - | - | ||
| 17 | 6-epi-ophiobolin G | Emericella variecolor 4 GF10 | [26] | Ulocladium sp. | [29] |
| Aspergillus calidoustus | [24,40] | Emericella variecolor 4 | [27] | ||
| Aspergillus ustus cf-42 | [43] | - | - | ||
| 18 | (6a)-21,21-O-dihydroophiobolin G | Aspergillus ustus | [23,45] | Ulocladium sp. | [29] |
| 19 | (6a)-21-deoxyophiobolin G | Aspergillus ustus cf-42 | [43] | - | - |
| 20 | 3-anhydro-21-deoxyophiobolin G | Aspergillus ustus | [46] | - | - |
| 21 | (6a)-18,19,21,21-Otetrahydro-18,19-dihydroxy-ophiobolin G | Aspergillus ustus | [45] | - | - |
| 22 | (6a)-16,17-dihydro-21-deoxy-ophiobolin G | Aspergillus sp. | [7] | Aspergillus ustus cf-42 | [42] |
| 23 | Ophiobolin H | Aspergillus ustus | [43,44,47] | Cochliobolus heterostrophus 2 | [30] |
| Emericella variecolor4 GF10 | [26] | Aspergillus insuetus | [24,40] | ||
| 24 | (5a,6a)-ophiobolin H | Aspergillus ustus | [43,45] | - | - |
| 25 | 5-O-methylophiobolin H | Aspergillus ustus | [45] | - | - |
| 26 | (5a,6a)-5-O-methylophiobolin H | Aspergillus ustus | [45] | - | - |
| 27 | Ophiobolin I | Bipolaris oryzae | [13,14,33] | Drechslera oryzae 1 | [34] |
| Polyangium cellulosum 5 | [30] | Bipolaris sp. | [36] | ||
| Cochliobolus heterostrophus 2 | [35] | Drechslera gigantea | [12,28] | ||
| Bipolaris setariae NY1 | [37] | - | - | ||
| 28 | 6-epi-ophiobolin I | Drechslera oryzae 1 | [34] | - | - |
| 29 | 25-Hydroxyophiobolin I | Bipolaris oryzae | [13] | Drechslera oryzae 1 | [34] |
| 30 | Ophiobolin J | Drechslera oryzae 1 | [34] | Drechslera gigantea | [28] |
| 31 | 8-epi-ophiobolin J | Drechslera gigantea | [28] | - | - |
| 32 | 8-deoxyophiobolin J | Drechslera oryzae 1 | [34] | - | - |
| 33 | Ophiobolin K | Aspergillus ustus | [48] | Cochliobolus heterostrophus 2 | [18] |
| Emericella variecolor 4 | [26,27] | Aspergillus sp. | [7,24,40] | ||
| Aspergillus insuetus | [24,40] | Aspergillus calidoustus | [19,24,40] | ||
| 34 | 6-epi-Ophiobolin K | Aspergillus ustus | [48] | Cochliobolus heterostrophus 2 | [18] |
| Emericella variecolor 4 | [26,27] | Aspergillus sp. | [7,24,40] | ||
| Aspergillus insuetus | [24,40] | Ulocladium sp. | [29] | ||
| Aspergillus calidoustus | [19,24,40] | - | - | ||
| 35 | Ophiobolin L | Cochliobolus heterostrophus 2 | [30] | - | - |
| 36 | 6-epi-ophiobolin L | Penicillium patulum 6 | [30] | - | - |
| 37 | Ophiobolin M | Cochliobolus heterostrophus 2 | [49] | - | - |
| 38 | 6-epi-ophiobolin M | Cochliobolus heterostrophus 2 | [18] | - | - |
| 39 | 6-epi-ophiobolin N | Emericella variecolor 4 GF10 | [26] | Aspergillus insuetus | [24,40] |
| 40 | Ophiobolin O | Aspergillus sp. | [7] | - | - |
| 41 | 6-epi-ophiobolin O | Aspergillus sp. | [7] | - | - |
| 42–46 | Ophiobolin P-T | Ulocladium sp. | [29] | - | - |
| 47–49 | Ophiobolin U-W | Aspergillus insuetus | [24,40] | Aspergillus ustus | [43] |
| Plant | 1 | 2 | 3 | 4 | 7 | 10 | 27 | 29 |
|---|---|---|---|---|---|---|---|---|
| Barley | >4.5 a | 3.0–4.5 a | 3.0–4.5 a | <0.5 a | 0.5–1.5 a | NA | 0.5–1.5 a | NA |
| Cabbage | 0–2 *, b | >2 *, b | >0 *, b | >0 *, b | >0 *, b | NA | >0 *, b | NA |
| Cocklebur | 0.5–1.5 a | 1.5–3.0 a | 1.5–3.0 a | <0.5 a | 0.5–1.5 a | NA | 0.5–1.5 a | NA |
| Corn | 3.0–4.5 a | 1.5–3.0 a | 3.0–4.5 a | 0.5–1.5 a | 0.5–1.5 a | NA | 0.5–1.5 a | NA |
| >2 *, b | 0–2 *, b | >2 *, b | >0 *, b | >0 *, b | NA | N *, b | NA | |
| 2–3 c | >3 c | NA | NA | 0.5–2.0 c | 0.5–2.0 c | 0.5–2.0 c | 0.5–2.0 c | |
| Goose grass | 1.5–3.0 a | 3.0–4.5 a | 1.5–3.0 a | 1.5–3.0 a | 1.5–3.0 a | NA | 1.5–3.0 a | NA |
| Grain sorghum | 3.0–4.5 a | 1.5–3.0 a | 3.0–4.5 a | 1.5–3.0 a | 3.0–4.5 a | NA | 0.5–1.5 a | NA |
| Johnson grass | 2–3 c | >3 c | NA | NA | 0.5–2.0 c | 0.5–2.0 c | 0.5–2.0 c | 0.5–2.0 c |
| Rice | 3.0–4.5 a | 1.5–3.0 a | 3.0–4.5 a | 0.5–1.5 a | 0.5–1.5 a | NA | 0.5–1.5 a | 3.0–4.5 a |
| >0 *, b | 0–2 *, b | 0–2 *, b | >0 *, b | 0–2 *, b | NA | >0 *, b | NA | |
| Sorghum | 2–3 c | 2–3 c | NA | NA | 0.5–2.0 c | 0.5–2.0 c | 0.5–2.0 c | 0.5–2.0 c |
| Soybean | 1.5–3.0 a | 1.5–3.0 a | 1.5–3.0 a | <0.5 a | 1.5–3.0 a | NA | 0.5–1.5 a | NA |
| Tobacco | >0 *, b | >0 *, b | >0 *, b | N b | >0 *, b | NA | >0 *, b | NA |
| Velvet-leaf | 0.5–1.5 a | <0.5 a | 1.5–3.0 a | <0.5 a | <0.5 a | NA | 0.5–1.5 a | NA |
| Number | Antibacterial Activity | ||||||
|---|---|---|---|---|---|---|---|
| E. coli * | S. aureus * | MRSA | M. intracellulare | B. subtilis | BCG | M. smegmatis | |
| 1 | >62 a | 62/7–12 a | NA | 62/3–6 a | NA | NA | NA |
| 3,4 | NA | >100 b | >100 b | NA | >100 b | >100 b | NA |
| 5 | NA | 31 b | 31 b | NA | 31 b | 31 b | NA |
| 7 | >62 a | 62/3–6 a | NA | >62 a | NA | NA | NA |
| 16 | >2.7 × 103 c | NA | NA | NA | 136/10 c | NA | NA |
| 17 | NA | NA | >250d | NA | >250 d | 125/NR d | 68/NR e |
| 18 | NA | NA | >250 d | NA | >250 d | 125/NR d | NA |
| 23 | >2.6 × 103 c | >1.6 × 103 f | NA | NA | 682/16 c | NA | NA |
| 24 | 1.6 × 103/10 f | >1.6 × 103 f | NA | NA | NA | NA | NA |
| 27 | >63 a | >63 a | NA | 63/3–6 a | NA | NA | NA |
| 33 | NA | NA | NA | NA | NA | 64/NR e | 33/NR e |
| 34 | NA | NA | 125/NR d | NA | 125/NR d | >250 d | 260/NR e |
| 35 | >60.a | >60 a | NA | 60/3–6 a | NA | NA | NA |
| 36 | >60 a | 60/>12 a | NA | >60 a | NA | NA | NA |
| 42 | NA | NA | 31/NR d | NA | 62/NR d | >250 d | NA |
| 43–45 | NA | NA | >250 d | NA | >250 d | >250 d | NA |
| 46 | NA | NA | 16/NR d | NA | 31/NR d | 31/NR d | NA |
| 47 | 1.6 × 103/15 f | 1.6 × 103/10 f | NA | NA | NA | NA | NA |
| Number | Species | Concentration (LD50, μM) | Reference |
|---|---|---|---|
| 10 | C. elegans | 5 | [18] |
| 11 | C. elegans | 130 | [18] |
| 12 | C. elegans | 25 | [18] |
| 33 | C. elegans | 26 | [18] |
| T. cruzi | 13 | [19] | |
| 34 | C. elegans | >260 | [18] |
| T. cruzi | 9.6 | [19] | |
| 37 | C. elegans | 13 | [18] |
| 38 | C. elegans | 130 | [18] |
| Number | Cell Line | IC50 (μM) | Reference | Cell Line | IC50 (μM) | Reference |
|---|---|---|---|---|---|---|
| 1 | CLL | 1 × 10−3 * | [24] | P388 | 0.06 | [35] |
| HUVECs | 0.08 | [58] | MNA | <0.12 | [59] | |
| HT-29 | 0.12 | [35] | A549 | 0.12, 0.42 ± 0.01 | [35,58] | |
| FFL | 0.12 | [59] | PK-15 | 0.12–0.24 | [59] | |
| OVCAR3 | 0.28 | [58] | B16F10 | 0.29 ± 0.05 | [21] | |
| L1210 | 0.3 | [60] | SKMEL28 | 0.37 ± 0.03 | [21] | |
| RPMI8226 | 0.4 | [61] | Hs683 | 0.62 ± 0.04 | [21] | |
| MM1R | 0.7 | [61] | U266B1 | 0.7 | [61] | |
| U373-MG | 0.87 ± 0.01 | [62] | Jurkat | 1.0 | [61] | |
| A2780 | 1.2 | [61] | RD | 1–2, 1.3 | [63,64] | |
| KB 3-1 | 1.4 | [64] | RH30 | 1.5 | [64] | |
| MDAMB-231 | 1.9, 0.7 | [61,64] | T98G | 1.9 ± 0.2 | [62] | |
| PC3 | 2.5 | [61] | U2OS | 2.8 | [64] | |
| GL19 | 3.7 ± 1.4 | [62] | U-87MG | 3.8 | [64] | |
| MCF-7 | 4.3; 4.0 | [61,64] | LLC-GA5-COL150 | 7.9 ± 0.40 | [65] | |
| hPBMC | 21 | [61] | HeLa | 62; 4.5 | [33,64] | |
| KB | 78 | [33] | - | - | - | |
| 2 | HeLa | 14 | [33] | KB | 22 | [33] |
| 3 | HeLa | 10 | [33] | KB | 12 | [33] |
| K562 | 40 ± 5.5 | [38] | HepG 2 | 56 ± 3.8 | [38] | |
| CLL | Inactive * | [24] | - | - | - | |
| 4 | B16F10 | 22 ± 3 | [21] | SKMEL28 | 27 ± 0.4 | [21] |
| A549 | 30 ± 1 | [21] | Hs683 | 30 ± 3 | [21] | |
| K562 | 36 ± 4.4 | [38] | HepG 2 | 47 ± 4.8 | [38] | |
| CLL | Inactive * | [24] | - | - | - | |
| 5 | K562 | 4.1 ± 0.50 | [38] | HepG2 | 6.5 ± 0.34 | [38] |
| 7 | CLL | 2 × 10−3 * | [24] | - | - | - |
| 10 | CLL | 8 × 10−3 * | [24] | - | - | - |
| 16 | P388 | 25 | [7] | - | - | - |
| 17 | HepG2 | 0.37 ± 0.03 | [29] | KB | 1.4 ± 0.06 | [29] |
| CLL | Inactive * | [24] | - | - | ||
| 18 | HepG2 | 2.2 ± 0.18 | [29] | KB | 4.0 ± 0.22 | [29] |
| L5178Y | >27 | [45] | - | - | - | |
| 21 | L5178Y | >25 | [45] | - | - | - |
| 23 | P388 | 106 | [7] | CLL | Inactive * | [24] |
| 24–26 | L5178Y | >25 | [45] | - | - | - |
| 7 | HeLa | 0.25 | [33] | HUVECs | >1 | [58] |
| KB | 2.3 | [33] | OVCAR3 | 71 | [58] | |
| 33 | CLL | 4 × 10−3 * | [24] | ACHN | 0.27 | [26] |
| HCT116 | 0.33 | [26] | T-47D | 0.35 | [26] | |
| P388/ADR | 0.36 | [26] | TK-10 | 0.51 | [19] | |
| MCF-7 | 0.51 | [19] | P388 | 0.51; 13 | [7,26] | |
| MDA-MB-231 | 0.57 | [26] | NCI-H460 | 0.57 | [26] | |
| HOP18 | 0.65 | [26] | - | - | - | |
| 34 | HepG2 | 1.9 ± 0.11 | [29] | TK-10 | 3 | [19] |
| MCF-7 | 3 | [19] | KB | 4.7 ± 0.72 | [29] | |
| P388 | 25 | [7] | CLL | Inactive * | [24] | |
| 39 | CLL | Inactive * | [24] | - | - | - |
| 40 | P388 | 4.7 | [7] | MCF-7 | 13 ± 1.3 | [66] |
| HUVEC | 85 ± 12 | [66] | DU145 | 16 ± 2.7 | [66] | |
| NCI-H460 | 14 ± 1.0 | [66] | - | - | - | |
| 41 | P388 | 9.3 | [7] | - | - | - |
| 42 | HepG2 | 1.5 ± 0.10 | [29] | KB | 6.2 ± 0.25 | [29] |
| 43 | HepG2 | 1.3 ± 0.12 | [29] | KB | 2.4 ± 0.21 | [29] |
| 44 | HepG2 | 1.2 ± 0.08 | [29] | KB | 2.9 ± 0.28 | [29] |
| 45 | HepG2 | 1.4 ± 0.08 | [29] | KB | 4.9 ± 0.32 | [29] |
| 46 | HepG2 | 0.24 ± 0.02 | [29] | KB | 3.0 ± 0.24 | [29] |
| Cell Death | Observations | Ophs | Cell Lines | Reference |
|---|---|---|---|---|
| Apoptosis | Cell soma shrinkage, chromatin condensation, typical apotptotic DNA ladder | 1 | L1210 | [60] |
| Changes of caspase 3/7 | 1 | MDA-MB-231 | [61] | |
| * AV+/PI−, AV+/PI+ and PARP increase | 1 | RD, RH30 | [64] | |
| AV+/PI+ increase | 1 | MDA-MB-231, MCF-7 HeLa | [64] | |
| AV+, Caspase-3 activation | 1, 7, 10, 33 | CLL | [24] | |
| Bcl-2 decrease | 5 | K562 | [38] | |
| Bcl-2 decrease | 40 | MCF-7 | [77] | |
| Necrosis | AV−/PI+ increase | 1 | KB 3-1, U2OS, U-87 MG | [64] |
| Paraptosis | BKCa decreasing, cytoplasmic vacuolization and mitochondrial swelling | 1 | U373-MG, GL19 | [62] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, W.; Deng, Z.; Hong, K. The Biological Activities of Sesterterpenoid-Type Ophiobolins. Mar. Drugs 2017, 15, 229. https://doi.org/10.3390/md15070229
Tian W, Deng Z, Hong K. The Biological Activities of Sesterterpenoid-Type Ophiobolins. Marine Drugs. 2017; 15(7):229. https://doi.org/10.3390/md15070229
Chicago/Turabian StyleTian, Wei, Zixin Deng, and Kui Hong. 2017. "The Biological Activities of Sesterterpenoid-Type Ophiobolins" Marine Drugs 15, no. 7: 229. https://doi.org/10.3390/md15070229
APA StyleTian, W., Deng, Z., & Hong, K. (2017). The Biological Activities of Sesterterpenoid-Type Ophiobolins. Marine Drugs, 15(7), 229. https://doi.org/10.3390/md15070229