The Marine Natural Product Pseudopterosin Blocks Cytokine Release of Triple-Negative Breast Cancer and Monocytic Leukemia Cells by Inhibiting NF-κB Signaling




Abstract
:1. Introduction
2. Results
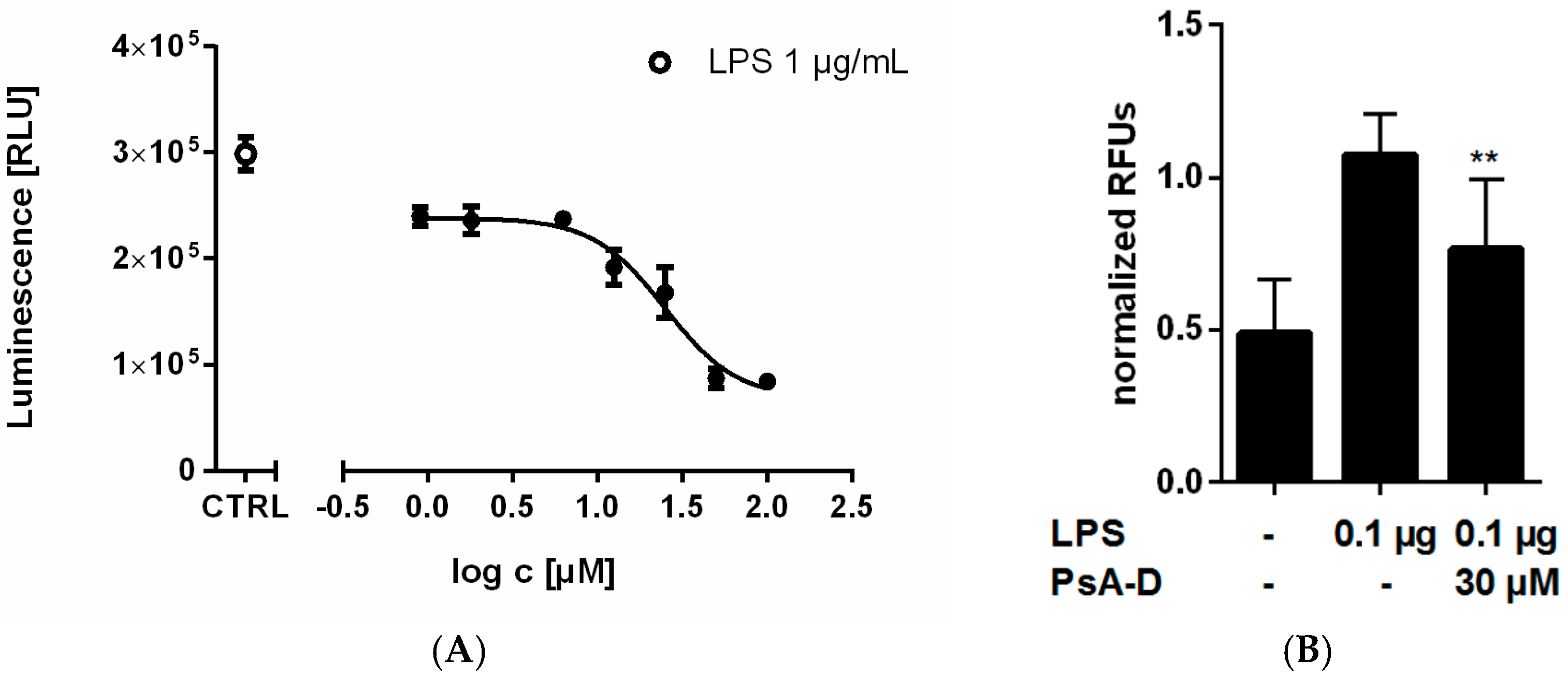
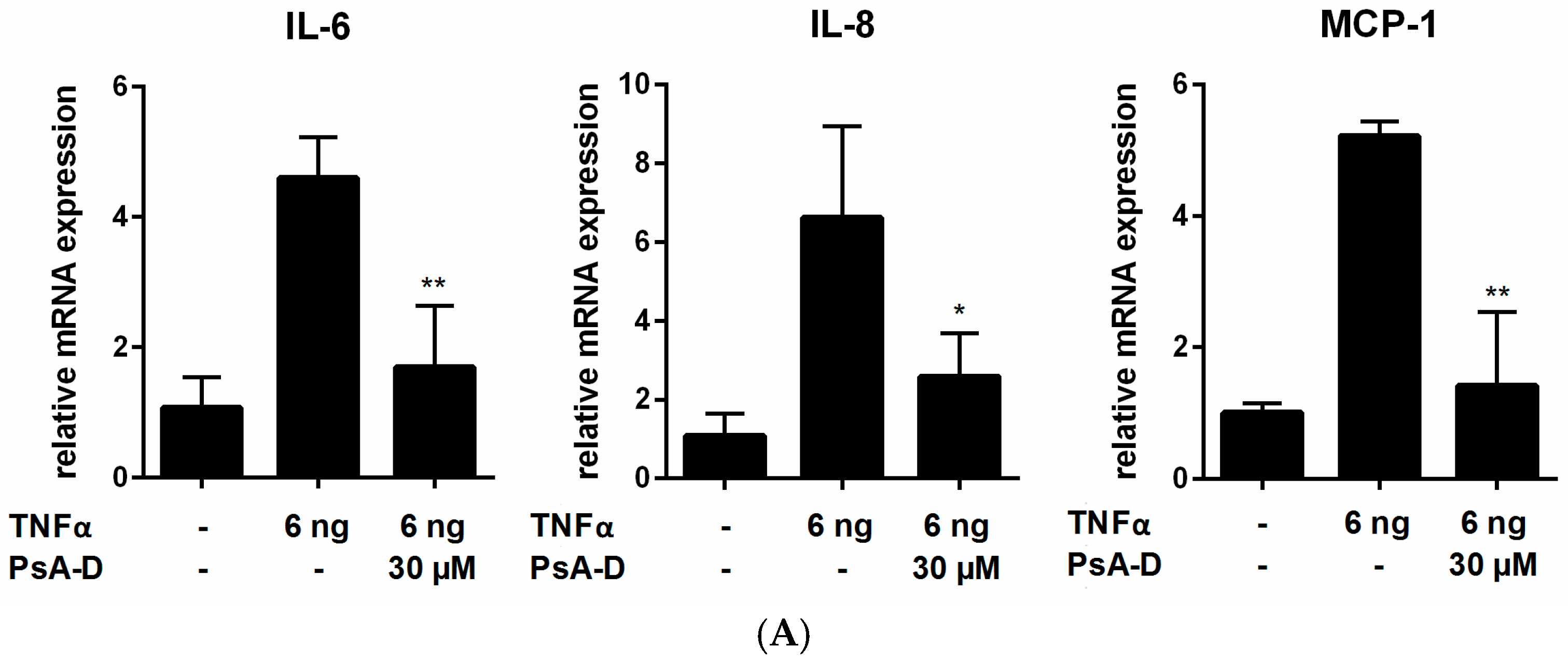
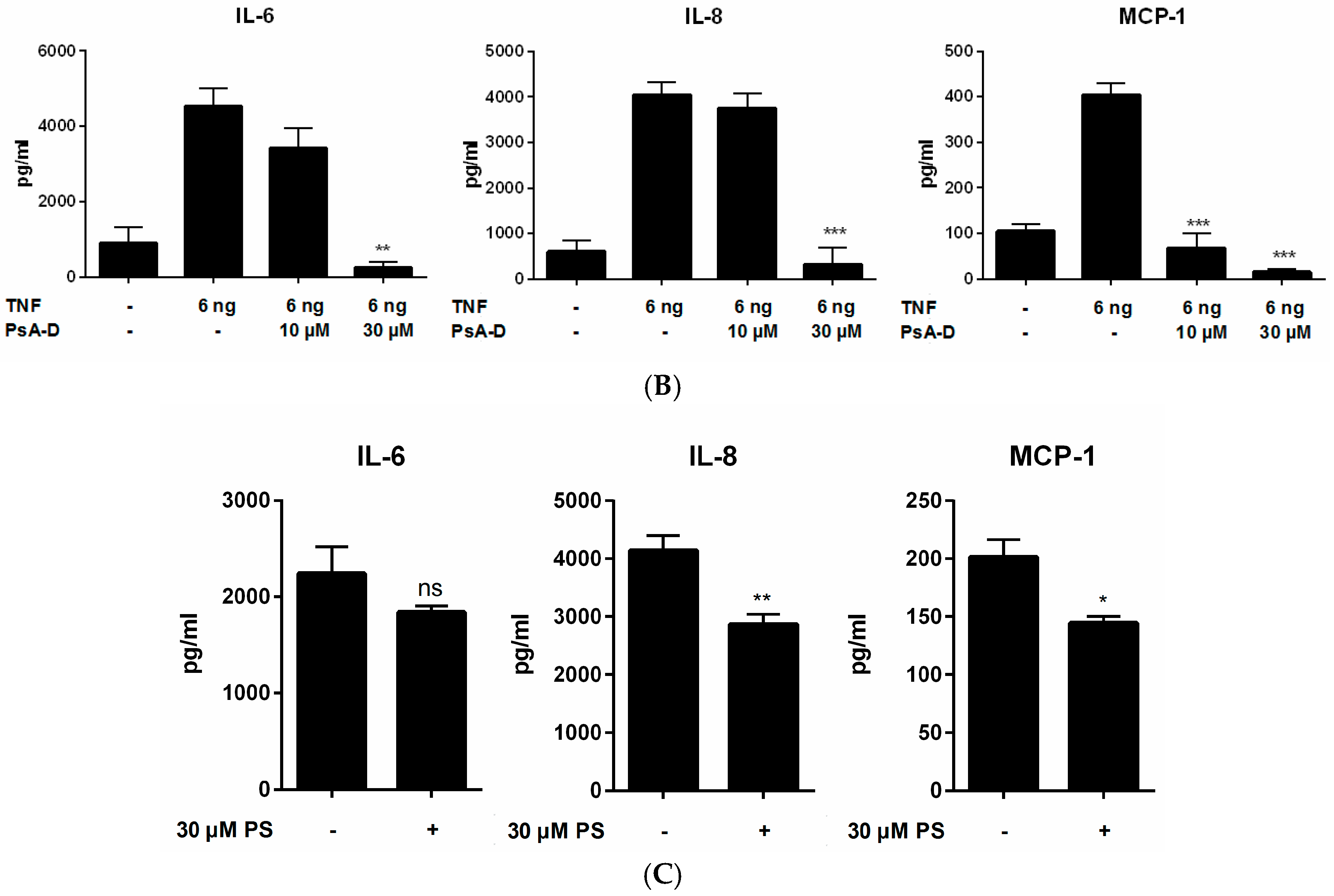
2.1. Pseudopterosin Reduces Cytokine Release by Inhibition of NF-κB Signaling
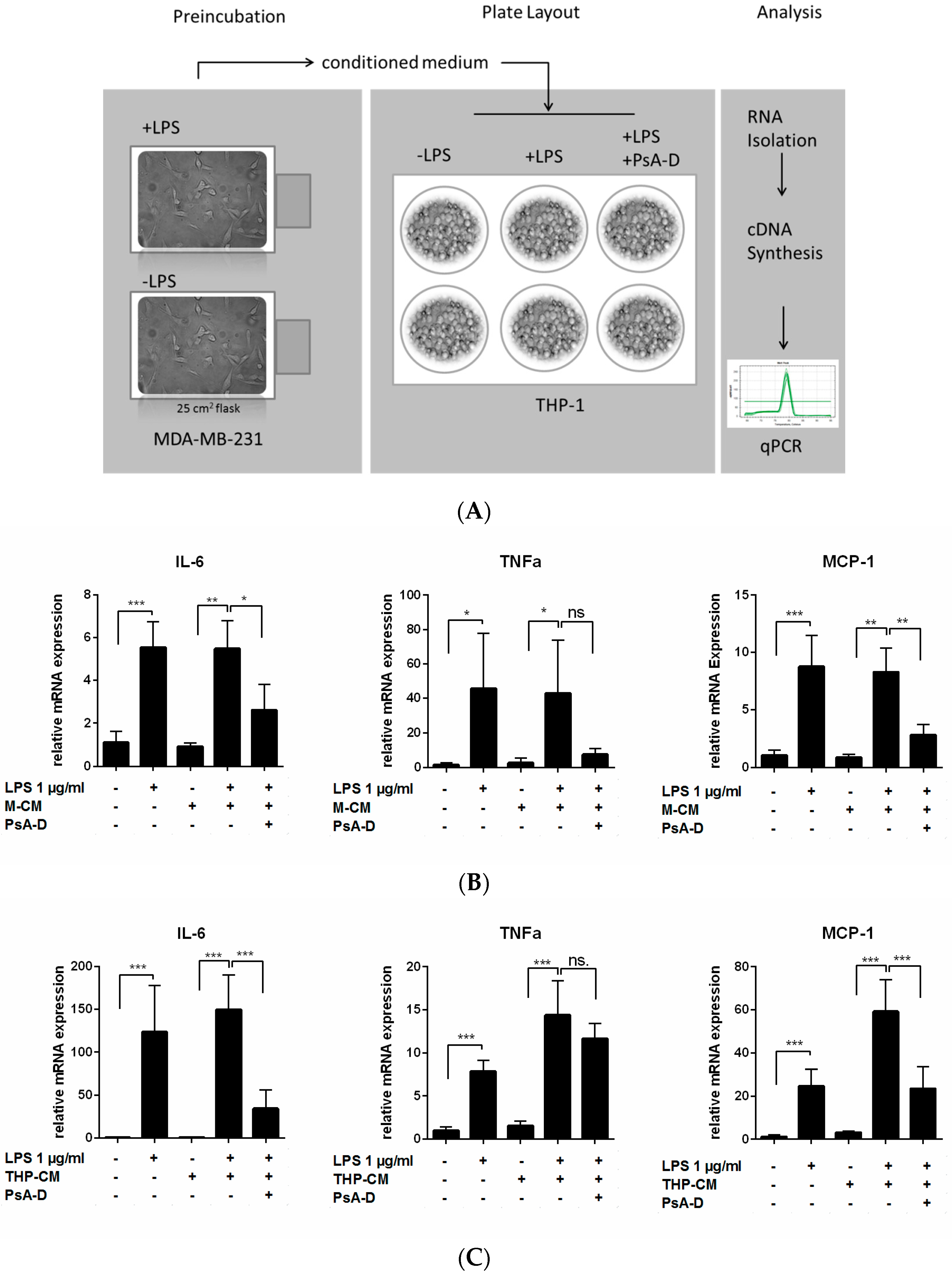
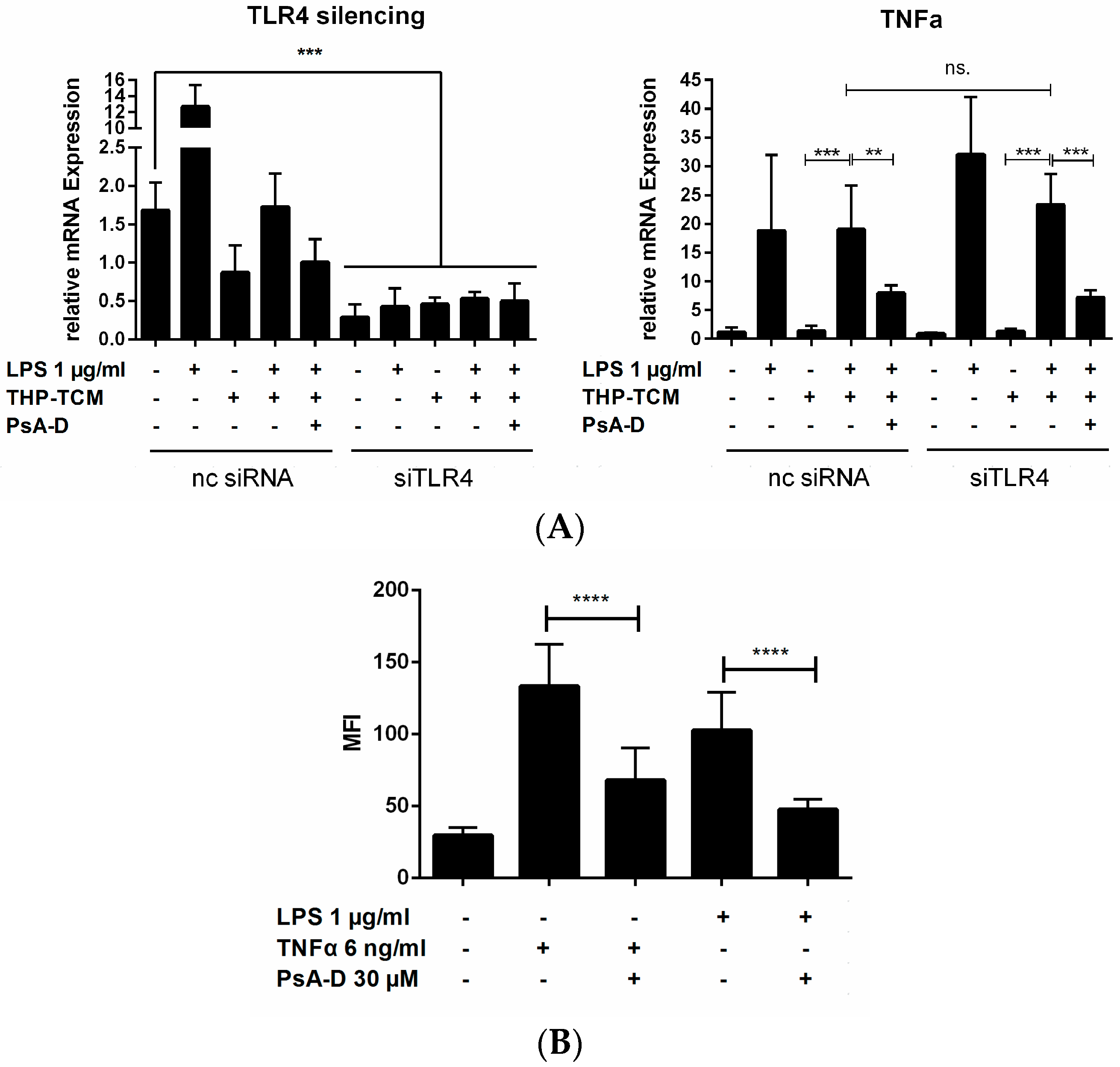
2.2. Pseudopterosin Blocks Bidirectional Communication
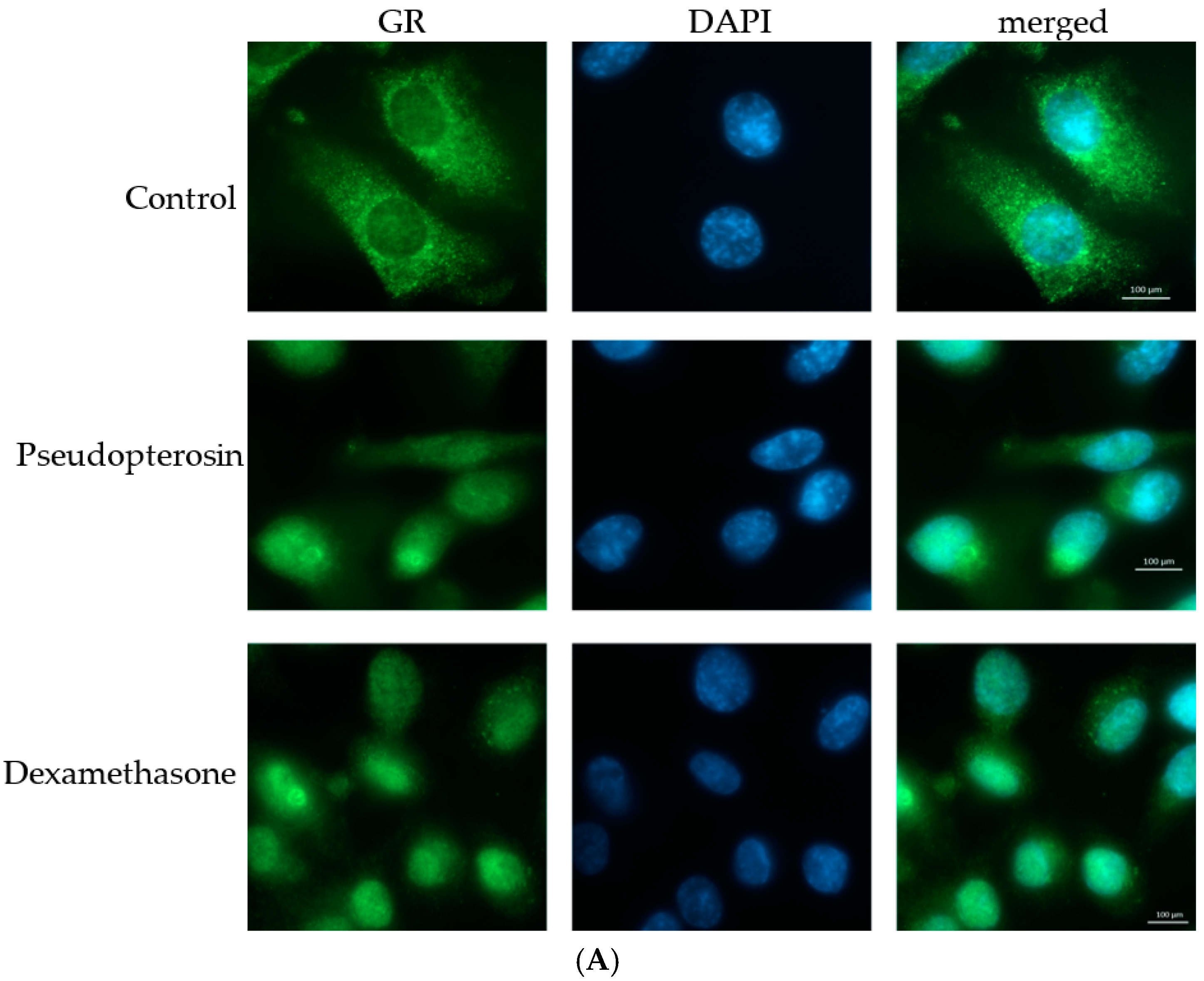
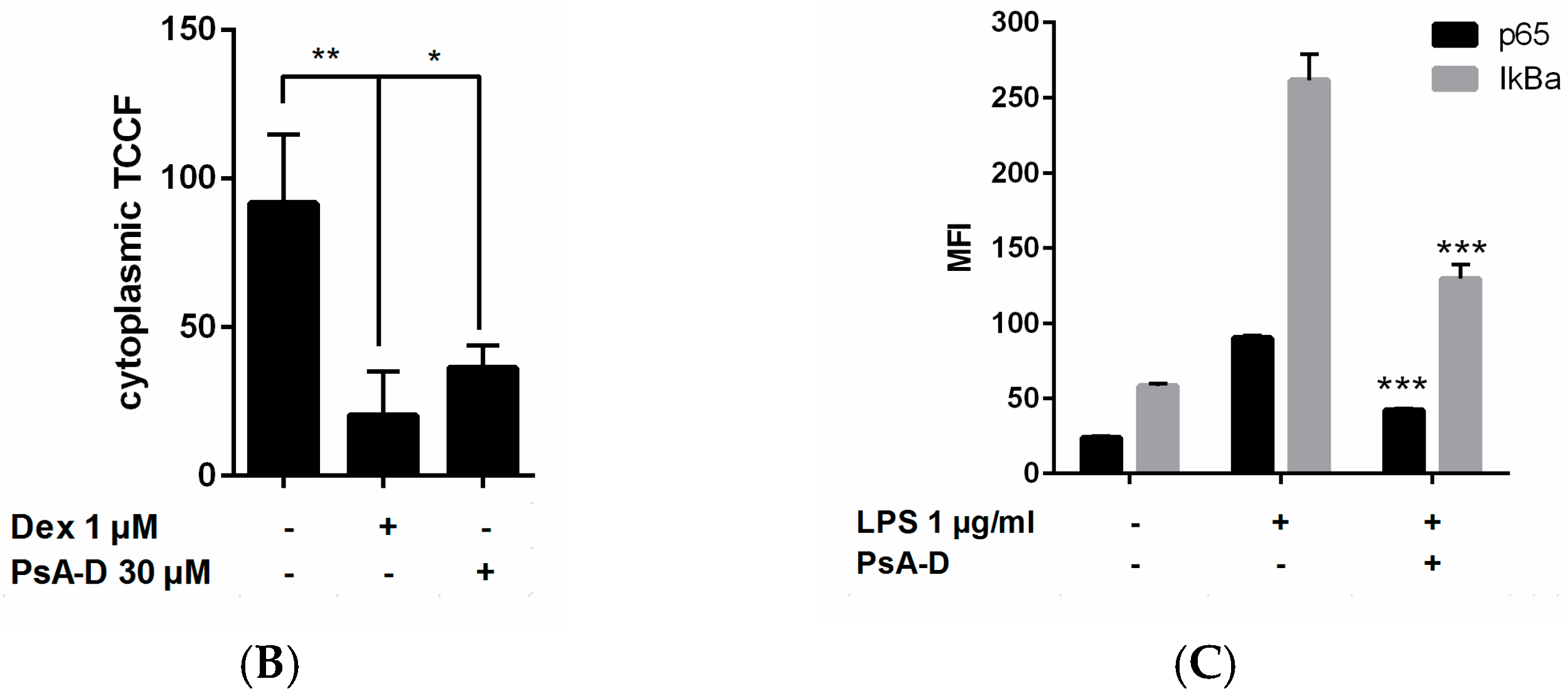
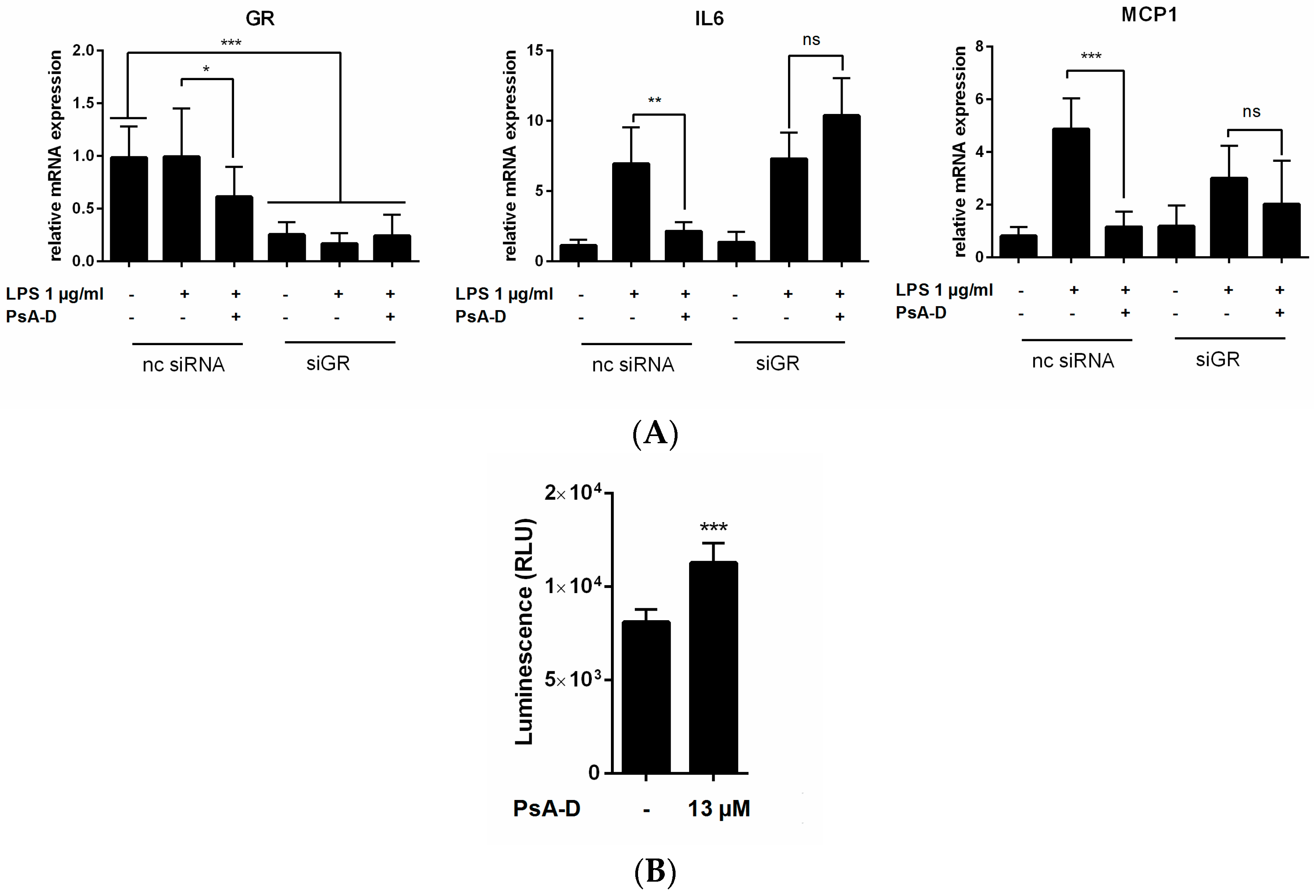
2.3. Pseudopterosin Inhibits NF-κB through Activation of the Glucocorticoid Receptor
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Commercially Available Reagents
4.2. Stable Cell Line Generation
4.3. NF-κB Reportergene Assay
4.4. NF-κB and Human Cytokine Magnetic Bead Kit
4.5. Quantitative Real-Time PCR
4.6. Immunofluorecent Staining
4.7. Conditioned Medium (CM) from Tumor Cells
4.8. Knock-Down Studies
4.9. GR Reportergene Assay
4.10. Preparation of PsA-D Mixture
4.11. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Moutsatsou, P.; Papavassiliou, A.G. The glucocorticoid receptor signalling in breast cancer. J. Cell. Mol. Med. 2008, 12, 145–163. [Google Scholar] [CrossRef] [PubMed]
- Belova, L.; Delgado, B.; Kocherginsky, M.; Melhem, A.; Olopade, O.; Conzen, S. Glucocorticoid receptor expression in breast cancer associates with older patient age. Breast Cancer Res. Treat. 2015, 116, 441–447. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.K.; Childs, B.H.; Pegram, M. Triple negative breast cancer: Unmet medical needs. Breast Cancer Res. Treat. 2011, 125, 627–636. [Google Scholar] [CrossRef] [PubMed]
- Abad, M.J.; Bermejo, P. Bioactive natural products from marine sources. Stud. Nat. Prod. Chem. 2001, 25, 683–755. [Google Scholar]
- Berrué, F.; McCulloch, M.W.B.; Kerr, R.G. Marine diterpene glycosides. Bioorg. Med. Chem. 2011, 19, 6702–6719. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M.S.; Jacobson, P.B.; Fenical, W.; Jacobs, R.S.; Glaser, K.B. Pharmacological characterization of the pseudopterosins: Novel anti-inflammatory natural products isolated from the Caribbean soft coral, Pseudopterogorgia elisabethae. Life Sci. 1998, 62, 401–407. [Google Scholar] [CrossRef]
- Correa, H.; Valenzuela, A.L.; Ospina, L.F.; Duque, C. Anti-inflammatory effects of the gorgonian Pseudopterogorgia elisabethae collected at the Islands of Providencia and San Andrés (SW Caribbean). J. Inflamm. Lond. 2009, 6. [Google Scholar] [CrossRef] [PubMed]
- Ata, A.; Kerr, R.G.; Moya, C.E.; Jacobs, R.S. Identification of anti-inflammatory diterpenes from the marine gorgonian Pseudopterogorgia elisabethae. Tetrahedron 2003, 59, 4215–4222. [Google Scholar] [CrossRef]
- Look, S.A.; Fenical, W.; Matsumoto, G.K.; Clardy, J. The pseudopterosins: A new class of antiinflammatory and analgesic diterpene pentosides from the marine sea whip Pseudopterogorgia elisabethae (Octocorallia). J. Org. Chem. 1986, 51, 5140–5145. [Google Scholar] [CrossRef]
- Look, S.A.; Fenical, W.; Jacobs, R.S.; Clardy, J. The pseudopterosins: Anti-inflammatory and analgesic natural products from the sea whip Pseudopterogorgia elisabethae. Proc. Natl. Acad. Sci. USA 1986, 83, 6238–6240. [Google Scholar] [CrossRef] [PubMed]
- Caplan, S.L.; Zheng, B.; Dawson-Scully, K.; White, C.A.; West, L.M. Pseudopterosin A: Protection of synaptic function and potential as a neuromodulatory agent. Mar. Drugs 2016, 14, 55. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Marine natural products and related compounds in clinical and advanced preclinical trials. J. Nat. Prod. 2004, 67, 1216–1238. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M.S.; Glaser, K.B.; Cuevas, C.; Jacobs, R.S.; Kem, W.; Little, R.D.; Mcintosh, J.M.; Newman, D.J.; Potts, B.C.; Shuster, D.E. The odyssey of marine pharmaceuticals: A current pipeline perspective. Trends Pharmacol. Sci. 2010, 31, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M.S.; Rodriguez, A.D.; Berlinck, R.G.S.; Hamann, M.T. Marine pharmacology in 2003–4: Marine compounds with anthelminthic, antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiplatelet, antiprotozoal, antituberculosis, and antiviral activities; affecting the cardiovascular, immune. Comp. Biochem. Physiol. C Toxicol. Pharmakol. 2008, 145, 553–581. [Google Scholar] [CrossRef] [PubMed]
- Moya, C.E.; Jacobs, R.S. Pseudopterosin A inhibits phagocytosis and alters intracellular calcium turnover in a pertussis toxin sensitive site in Tetrahymena thermophila. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2006, 143, 436–443. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, I.I.; Shi, Y.P.; García, O.J.; Rodríguez, A.D.; Mayer, A.M.S.; Sánchez, J.A.; Ortega-Barria, E.; González, J. New pseudopterosin and seco-pseudopterosin diterpene glycosides from two Colombian isolates of Pseudopterogorgia elisabethae and their diverse biological activities. J. Nat. Prod. 2004, 67, 1672–1680. [Google Scholar] [CrossRef] [PubMed]
- Badr, C.; Niers, J.M.; Tjon-Kon-Fat, L.-A.; Noske, D.P.; Wurdinger, T.; Tannous, B. Real-time monitoring of NF-kappaB activity in cultured cells and in animal models. Mol. Imaging 2009, 8, 278–290. [Google Scholar] [PubMed]
- Kawai, T.; Akira, S. The role of pattern-recognition receptors in innate immunity: Update on Toll-like receptors. Nat. Immunol. 2010, 11, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Ramage, L. Expression of Pro-Inflammatory Proteins in the Lung Epithelial Cell Line A549, in Response to Cytokine, and Environmental Particle Exposure. Ph.D. Thesis, Edinburgh Napier University, Edinburgh, UK, 2003. [Google Scholar]
- Blank, V.; Kourilsky, P.; Israel, A.; Publishers, E.S. NF-kB and related proteins: Rel/dorsal homologies meet ankyrin-like repeats. Trends Biochem. Sci. 1992, 17, 135–140. [Google Scholar] [CrossRef]
- Baeuerle, P.A.; Baltimore, D. Activation of DNA-binding activity in an apparently cytoplasmic precursor of the NF-kappa B transcription factor. Cell 1988, 53, 211–217. [Google Scholar] [CrossRef]
- Balkwill, F. TNF-alpha in promotion and progression of cancer. Cancer Metastasis Rev. 2006, 25, 409–416. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Blackwell, K.; Altaeva, A.; Shi, Z.; Habelhah, H. TRAF2 phosphorylation promotes NF-κB-dependent gene expression and inhibits oxidative stress-induced cell death. Mol. Biol. Cell 2010, 22, 128–140. [Google Scholar] [CrossRef] [PubMed]
- Perkins, N.D. Integrating cell-signalling pathways with NF-kappaB and IKK function. Nat. Rev. Mol. Cell Biol. 2007, 8, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.; Yao, Z. Activation of NF-κB in human breast cancer and its role in cell proliferation and progression. Chin. J. Clin. Oncol. 2006, 3, 5–10. [Google Scholar] [CrossRef]
- Zhao, X.; Sun, X.; Gao, F.; Luo, J.; Sun, Z. Effects of ulinastatin and docataxel on breast tumor growth and expression of IL-6, IL-8, and TNF-α. J. Exp. Clin. Cancer Res. 2011, 30, 22. [Google Scholar] [CrossRef] [PubMed]
- Park, M.; Hong, J. Roles of NF-κB in cancer and inflammatory diseases and their therapeutic approaches. Cells 2016, 5, 15. [Google Scholar] [CrossRef] [PubMed]
- Shostak, K.; Chariot, A. NF-κB, stem cells and breast cancer: The links get stronger. Breast Cancer Res. 2011, 13, 214. [Google Scholar] [CrossRef] [PubMed]
- Becker-Weimann, S.; Xiong, G.; Furuta, S.; Han, J.; Kuhn, I.; Akavia, U.-D.; Pe’er, D.; Bissell, M.J.; Xu, R. NFkB disrupts tissue polarity in 3D by preventing integration of microenvironmental signals. Oncotarget 2013, 4, 2010–2020. [Google Scholar] [CrossRef] [PubMed]
- Bissell, M.J.; Radisky, D. Putting tumors in context. Nat. Rev. Cancer 2001, 1, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Mestdagt, M.; Polette, M.; Buttice, G.; Noël, A.; Ueda, A.; Foidart, J. Transactivation of MCP-1/CCL2 by beta-catenin/TCF-4 in human breast cancer cells. Int. J. Cancer 2006, 118, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Keith, B.D. Systematic review of the clinical effect of glucocorticoids on nonhematologic malignancy. BMC Cancer 2008, 8, 84. [Google Scholar] [CrossRef] [PubMed]
- Skor, M.; Wonder, E.; Kocherginsky, M.; Goyal, A.; Hall, B.; Cai, Y.; Conzen, S. Glucocorticoid receptor antagonsims as a novel therapy for triple-negative breast cancer. Clin. Cancer Res. 2012, 100, 130–134. [Google Scholar]
- Mitre-Aguilar, I.B.; Cabrera-Quintero, A.J.; Zentella-Dehesa, A. Genomic and non-genomic effects of glucocorticoids: Implications for breast cancer. Int. J. Clin. Exp. Pathol. 2015, 8, 1–10. [Google Scholar] [PubMed]
- McKay, L.I.; Cidlowski, J.A. Molecular control of immune/inflammatory responses: Interactions between nuclear factor-κB and steroid receptor-signaling pathways. Endocr. Rev. 1999, 20, 435–459. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, S.; Rainer, J.; Ploner, C.; Presul, E.; RimL, S.; Kofler, R. Glucocorticoid-induced apoptosis and glucocorticoid resistance: Molecular mechanisms and clinical relevance. Cell Death Differ. 2004, 11, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.T.; Wang, L.H. New dimension of glucocorticoids in cancer treatment. Steroids 2016, 111, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Buxant, F.; Kindt, N.; Laurent, G.; Noel, J.; Saussez, S. Antiproliferative effect of dexamethasone in the MCF-7 breast cancer cell line. Mol. Med. Rep. 2015, 4051–4054. [Google Scholar] [CrossRef] [PubMed]
- Mehmeti, M.; Allaoui, R.; Bergenfelz, C.; Saal, L.H.; Ethier, S.P.; Johansson, M.E.; Jirström, K.; Leandersson, K. Expression of functional toll like receptor 4 in estrogen receptor/progesterone receptor-negative breast cancer. Breast Cancer Res. 2015, 17, 130. [Google Scholar] [CrossRef] [PubMed]
- Akira, S.; Uematsu, S.; Takeuchi, O. pathogen recognition and innate immunity. Cell 2006, 3, 783–801. [Google Scholar] [CrossRef] [PubMed]
- Tak, P.P.; Firestein, G.S. NF-κB: A key role in inflammatory diseases. J. Clin. Investig. 2001, 107, 7–11. [Google Scholar] [CrossRef] [PubMed]
- Pahl, H.L. Activators and target genes of Rel/NF-κB transcription factors. Oncogene 1999, 18, 6853–6866. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Takada, Y.; Boriek, A.; Aggarwal, B. Nuclear factor-κB: Its role in health and disease. J. Mol. Med. 2004, 82, 434–448. [Google Scholar] [CrossRef] [PubMed]
- Schütze, S.; Potthoff, K.; Machleidt, T. TNF activates NF-KB by phosphatidylcholine-specific C-induced “Acidic” sphingomyelin breakdown. Cell 1992, 71, 765–776. [Google Scholar] [CrossRef]
- Miyake, K. Innate recognition of lipopolysaccharide by Toll-like receptor 4-MD-2. Trends Microbiol. 2004, 12, 186–192. [Google Scholar] [CrossRef] [PubMed]
- Wajant, H.; Scheurich, P. TNFR1-induced activation of the classical NF-KB pathway. FEBS J. 2011, 278, 862–876. [Google Scholar] [CrossRef] [PubMed]
- Correa, H.; Aristizabal, F.; Duque, C.; Kerr, R. Cytotoxic and antimicrobial activity of pseudopterosins and seco-pseudopterosins isolated from the octocoral Pseudopterogorgia elisabethae of san andrés and providencia islands (Southwest Caribbean Sea). Mar. Drugs 2011, 9, 334–344. [Google Scholar] [CrossRef] [PubMed]
- Ata, A.; Win, H.Y.; Holt, D.; Holloway, P.; Segstro, E.P.; Jayatilake, G.S. New antibacterial diterpenes from Pseudopterogorgia elisabethae. Helv. Chim. Acta 2004, 87, 1090–1098. [Google Scholar] [CrossRef]
- Yamamoto, M.; Taguchi, Y.; Ito-kureha, T.; Semba, K.; Yamaguchi, N.; Inoue, J. NF-κB non-cell-autonomously regulates cancer stem cell populations in the basal-like breast cancer subtype. Nat. Commun. 2013, 4, 2299. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, N.; Ito, T.; Azuma, S.; Ito, E.; Honma, R.; Yanagisawa, Y.; Nishikawa, A.; Kawamura, M.; Imai, J.; Watanabe, S.; et al. Constitutive activation of nuclear factor-kappaB is preferentially involved in the proliferation of basal-like subtype breast cancer cell lines. Cancer Sci. 2009, 100, 1668–1674. [Google Scholar] [CrossRef] [PubMed]
- Ling, J.; Kumar, R. Crosstalk between NFkB and glucocorticoid signaling: A potential target of breast cancer therapy. Cancer Lett. 2012, 322, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Pires, B.R.B.; Mencalha, A.L.; Ferreira, G.M.; de Souza, W.F.; Morgado-Díaz, J.A.; Maia, A.M.; Corrêa, S.; Abdelhay, E.S.F.W. NF-kappaB is involved in the regulation of EMT genes in breast cancer cells. PLoS ONE 2017, 12, e0169622. [Google Scholar] [CrossRef] [PubMed]
- Gordon, A.H.; O’Keefe, R.J.; Schwarz, E.M.; Rosier, R.N.; Puzas, J.E. Nuclear factor-kB-dependent mechanisms in breast cancer cells regulate tumor burden and osteolysis in bone. Am. Assoc. Cancer Res. 2005, 65, 3209–3218. [Google Scholar] [CrossRef] [PubMed]
- Rutz, H.P. Effects of corticosteroid use on treatment of solid tumours. Lancet 2002, 360, 1969–1970. [Google Scholar] [CrossRef]
- Pan, D.; Kocherginsky, M.; Conzen, S.D. Activation of the glucocorticoid receptor is associated with poor prognosis in estrogen receptor-negative breast cancer. Cancer Res. 2011, 71, 6360–6370. [Google Scholar] [CrossRef] [PubMed]
- Mondal, S.K.; Jinka, S.; Pal, K.; Nelli, S.; Dutta, S.K.; Wang, E.; Ahmad, A.; AlKharfy, K.M.; Mukhopadhyay, D.; Banerjee, R. Glucocorticoid receptor-targeted liposomal codelivery of lipophilic drug and Anti-Hsp90 gene: Strategy to induce drug-sensitivity, EMT-reversal, and reduced malignancy in aggressive tumors. Mol. Pharm. 2016, 13, 2507–2523. [Google Scholar] [CrossRef] [PubMed]
- Radisky, D.C.; Bissell, M.J. NF-kappaB links oestrogen receptor signalling and EMT. Nat. Cell Biol. 2007, 9, 361–363. [Google Scholar] [CrossRef] [PubMed]
- Abduljabbar, R.; Negm, O.H.; Lai, C.-F.; Jerjees, D.A.; Al-Kaabi, M.; Hamed, M.R.; Tighe, P.J.; Buluwela, L.; Mukherjee, A.; Green, A.R.; et al. Clinical and biological significance of glucocorticoid receptor (GR) expression in breast cancer. Breast Cancer Res. Treat. 2015, 150, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Chrousos, G.P.; Kino, T. Intracellular glucocorticoid signaling: A formerly simple system turns stochastic. Sci. Signal. 2005, 2005, 48. [Google Scholar] [CrossRef] [PubMed]
- De Bosscher, K.; Vanden Berghe, W.; Haegeman, G. The interplay between the glucocorticoid receptor and nuclear factor-kB or activator protein-1: Molecular mechanisms for gene repression. Endocr. Rev. 2003, 24, 488–522. [Google Scholar] [CrossRef] [PubMed]
- Kino, T.; De Martino, M.U.; Charmandari, E.; Mirani, M.; Chrousos, G.P. Tissue glucocorticoid resistance/hypersensitivity syndromes. J. Steroid Biochem. Mol. Biol. 2003, 85, 457–467. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- McCloy, R.A.; Rogers, S.; Caldon, C.E.; Lorca, T.; Castro, A.; Burgess, A. Partial inhibition of Cdk1 in G2 phase overrides the SAC and decouples mitotic events. Cell Cycle 2014, 13, 1400–1412. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MDA-MB-231 | Control (pg/mL) | +LPS 1 µg/mL | +PsA-D 30 µM | p-Value | % Inhibition |
|---|---|---|---|---|---|
| IL-6 | 1626.3 ± 144 | 4666.7 ± 307 | 2874.8 ± 610 | <0.0002 | 38.3 |
| TNFα | 1.9 ± 0.6 | 29.1 ± 5.5 | 7.17 ± 3.4 | <0.0005 | 75.3 |
| MCP-1 | 325.3 ± 260 | 1625.6 ± 540.6 | 241.3 ± 100.9 | 0.0082 | 85.2 |
| THP-1 | Control (pg/mL) | +LPS 1 µg/mL | +PsA-D 30 µM | p-Value | % Inhibition |
| IL-6 | 2.8 ± 1 | 66.7 ± 9.8 | 33 ± 1.98 | 0.0089 | 50.0 |
| TNFα | 13.4 ± 4.5 | 439.4 ± 28 | 232.0 ± 100 | 0.1138 | 47.2 |
| MCP-1 | 182.9 ± 65.3 | 4436.7 ± 2098 | 1208.9 ± 762.3 | 0.0552 | 72.8 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sperlich, J.; Kerr, R.; Teusch, N. The Marine Natural Product Pseudopterosin Blocks Cytokine Release of Triple-Negative Breast Cancer and Monocytic Leukemia Cells by Inhibiting NF-κB Signaling. Mar. Drugs 2017, 15, 262. https://doi.org/10.3390/md15090262
Sperlich J, Kerr R, Teusch N. The Marine Natural Product Pseudopterosin Blocks Cytokine Release of Triple-Negative Breast Cancer and Monocytic Leukemia Cells by Inhibiting NF-κB Signaling. Marine Drugs. 2017; 15(9):262. https://doi.org/10.3390/md15090262
Chicago/Turabian StyleSperlich, Julia, Russell Kerr, and Nicole Teusch. 2017. "The Marine Natural Product Pseudopterosin Blocks Cytokine Release of Triple-Negative Breast Cancer and Monocytic Leukemia Cells by Inhibiting NF-κB Signaling" Marine Drugs 15, no. 9: 262. https://doi.org/10.3390/md15090262
APA StyleSperlich, J., Kerr, R., & Teusch, N. (2017). The Marine Natural Product Pseudopterosin Blocks Cytokine Release of Triple-Negative Breast Cancer and Monocytic Leukemia Cells by Inhibiting NF-κB Signaling. Marine Drugs, 15(9), 262. https://doi.org/10.3390/md15090262