Antifouling Compounds from Marine Macroalgae


Abstract
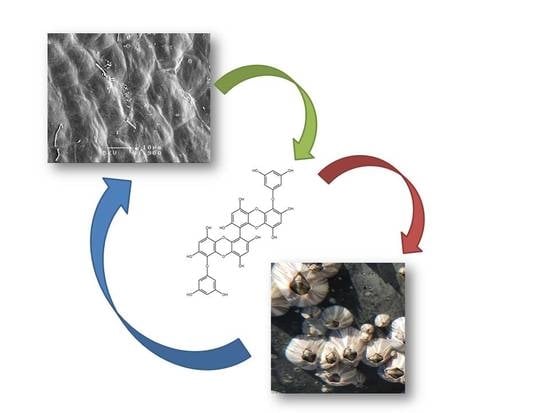
:
1. Introduction
2. Antifouling Compounds from Marine Macroalgae
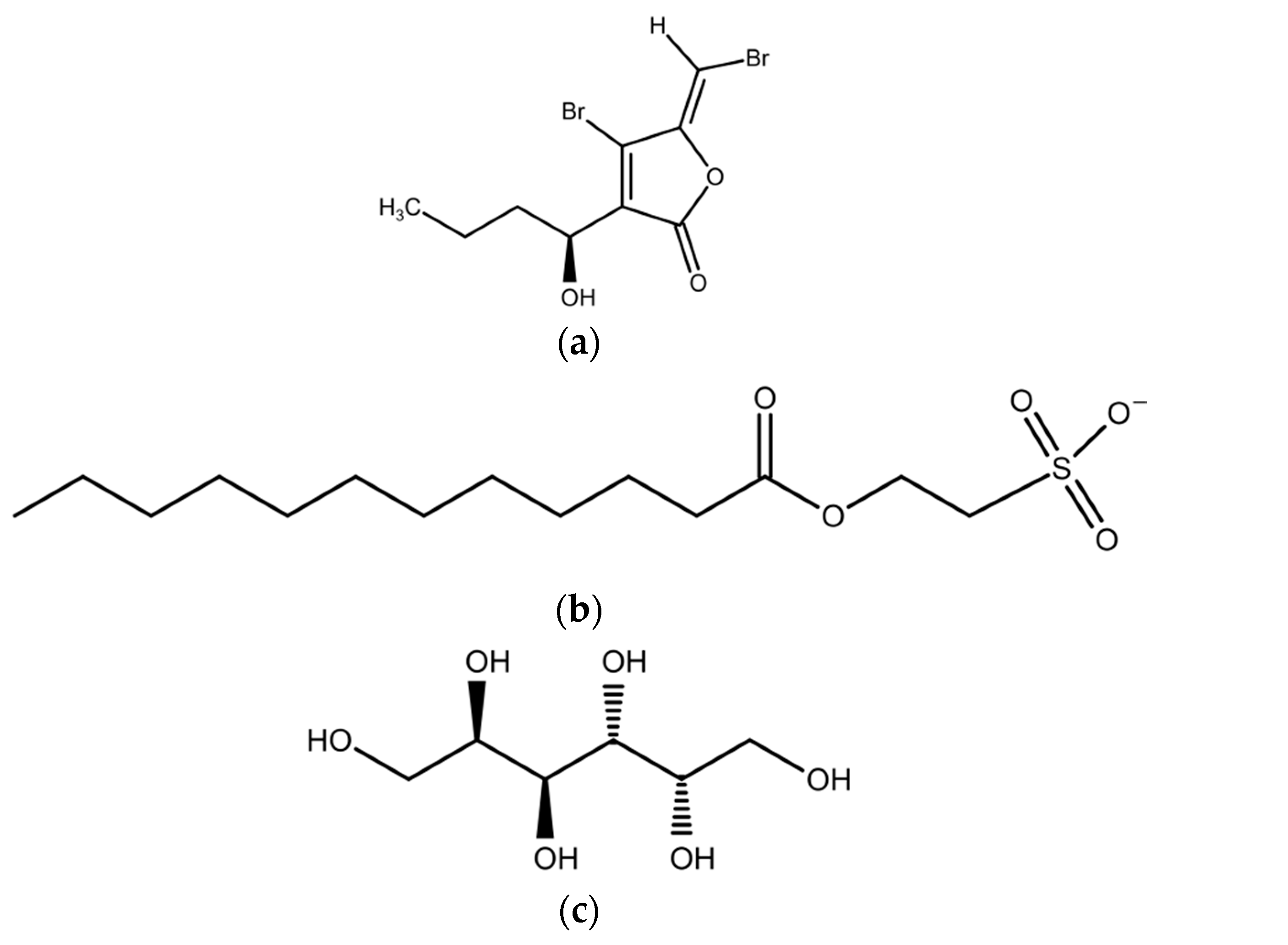
2.1. Green Macroalgae (Chlorophyta)
2.2. Brown Macroalgae (Phaeophyta)
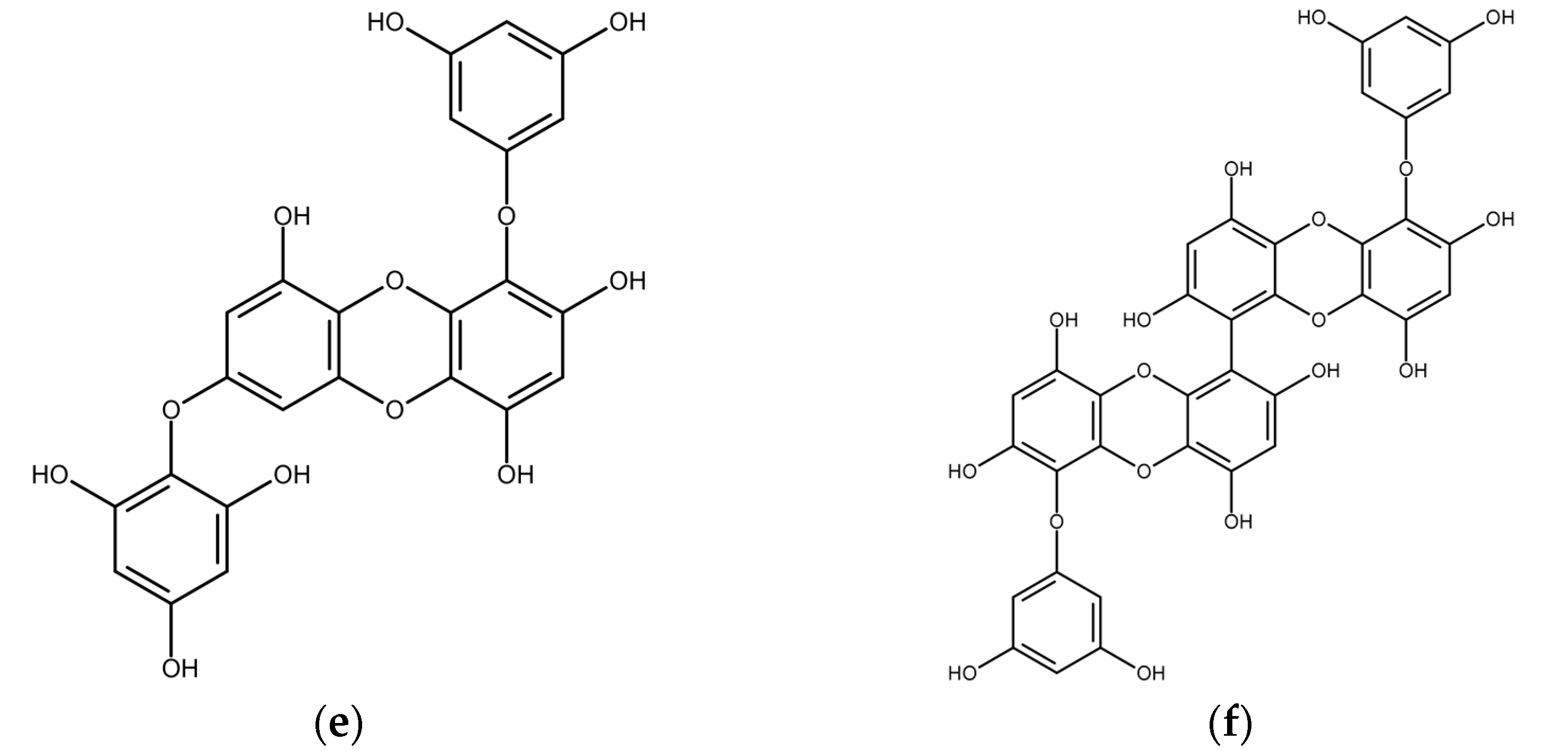
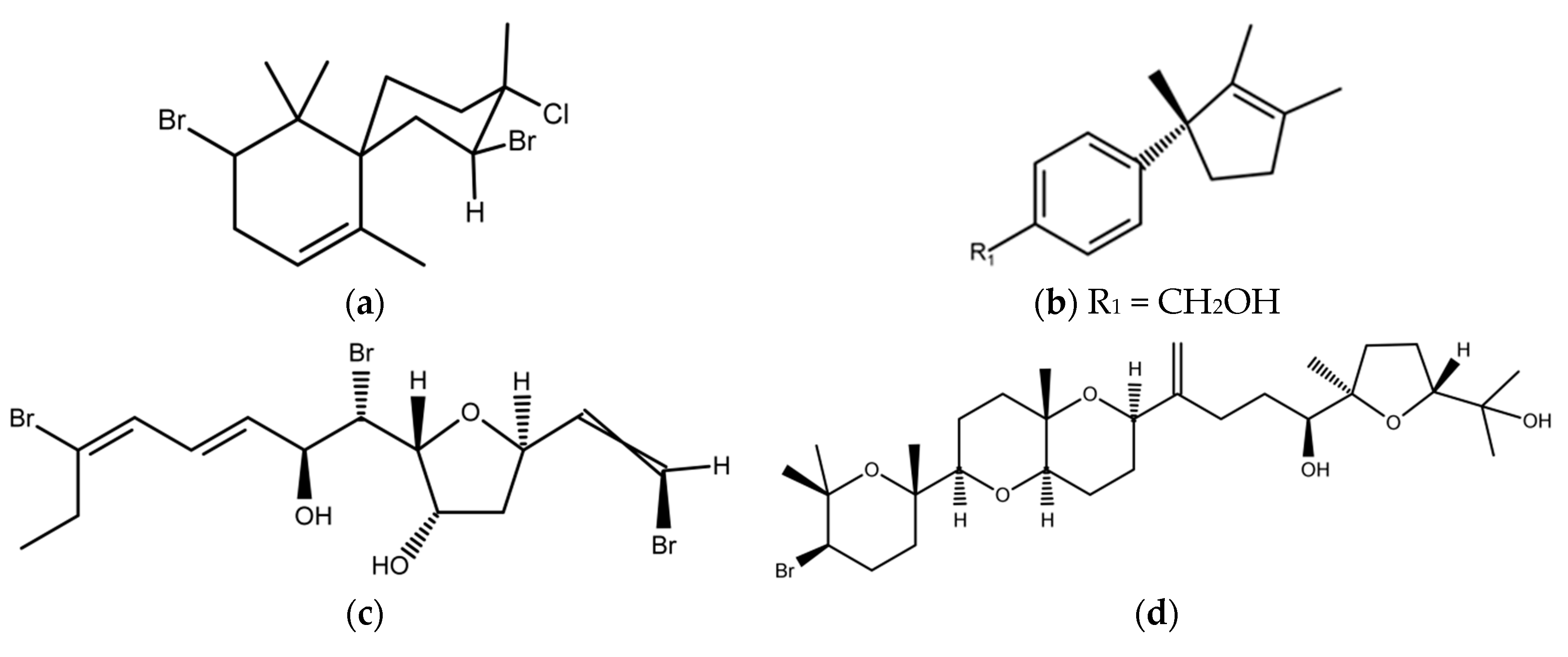
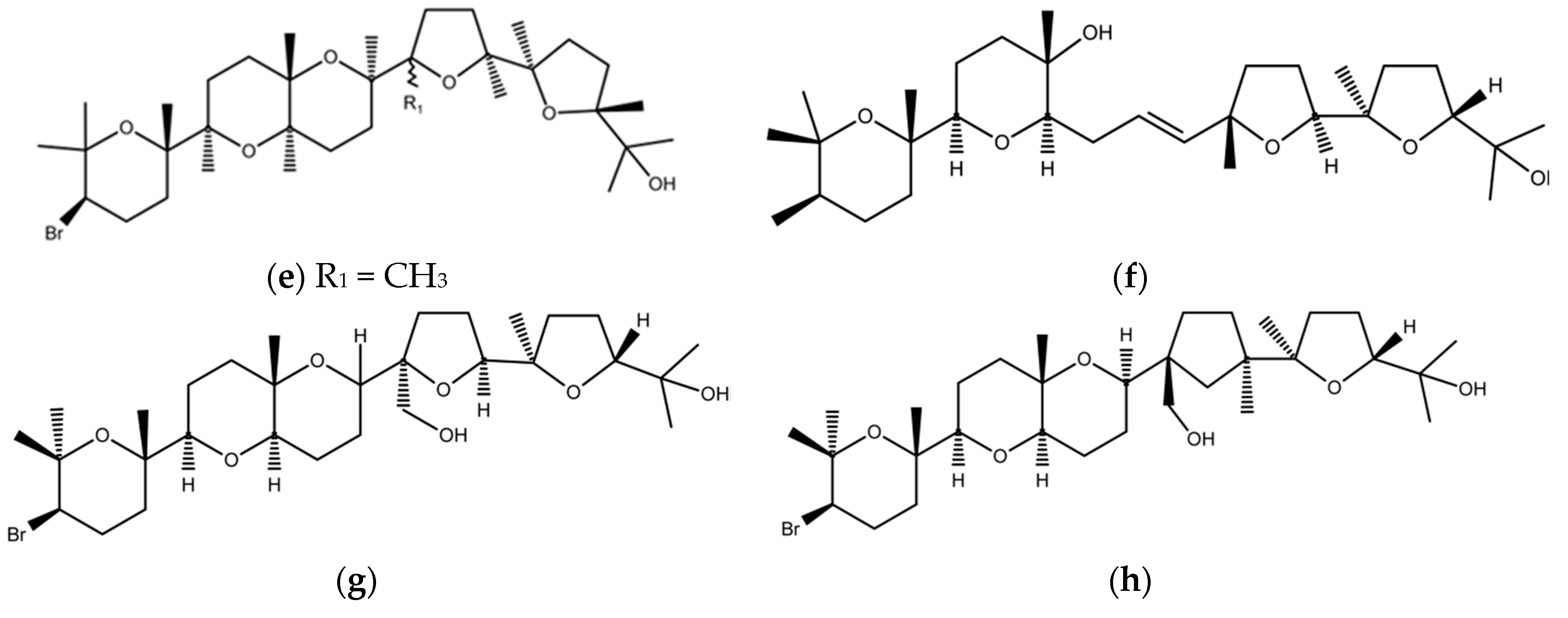
2.3. Red Macroalgae (Rhodophyta)
3. Quorum Sensing Inhibitors from Macroalgae
4. Potential of Macroalgal Extracts in Biological Synthesis of Nanoparticles
5. Antifouling Defense
6. Role of Epibiotic Organisms
7. Conclusions and Future Outlook
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Wahl, M. Marine epibiosis. I. Fouling and antifouling: Some basic aspects. Mar. Ecol. Prog. Ser. 1989, 58, 175–189. [Google Scholar] [CrossRef] [Green Version]
- Dobretsov, S.; Dahms, H.U.; Qian, P.Y. Inhibition of biofouling by marine microorganisms and their metabolites. Biofouling 2006, 22, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Wahl, M.; Goecke, F.; Labes, A.; Dobretsov, S.; Weinberger, F. The second skin: Ecological role of epibiotic biofilms on marine organisms. Front. Microbiol. 2012, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yebra, D.M.; Kiil, S.; Dam-Johansen, K. Antifouling technology—Past, present and future steps towards efficient and environmentally friendly antifouling coatings. Prog. Org. Coat. 2004, 50, 75–104. [Google Scholar] [CrossRef]
- Trepos, R.; Pinori, E.; Berglin, M.; Svenson, J.; Coutinho, R.; Lausmaa, J.; Hellio, C. Innovative approaches for the development of new copper-free marine antifouling paints. J. Ocean Technol. 2014, in press. [Google Scholar]
- Callow, M.E.; Callow, J.E. Marine biofouling: A sticky problem. Biologist 2002, 49, 10–14. [Google Scholar] [PubMed]
- Bers, A.V.; Prendergast, G.S.; Zurn, C.M.; Hansson, L.; Head, R.M.; Thomason, J.C. A comparative study of the anti-settlement properties of Mytilid shells. Biol. Lett. 2006, 2, 88–91. [Google Scholar] [CrossRef] [PubMed]
- Qian, P.-Y.; Xu, Y.; Fusetani, N. Natural products as antifouling compounds: Recent progress and future perspectives. Biofouling 2010, 25, 223–234. [Google Scholar] [CrossRef] [PubMed]
- Lee, R.E. Phycology; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Bhadury, P.; Wright, P.C. Exploitation of marine algae: Biogenic compounds for potential antifouling applications. Planta 2004, 219, 561–578. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2014, 31, 160–258. [Google Scholar] [CrossRef] [PubMed]
- Fusetani, N. Antifouling natural products. Nat. Prod. Rep. 2011, 28, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Qian, P.Y.; Chen, L.; Xu, Y. Mini-Review: Molecular mechanisms of antifouling compounds. Biofouling 2013, 29, 381–400. [Google Scholar] [CrossRef] [PubMed]
- Abed, R.; Dobretsov, S.; Kumar, S. Applications of cyanobacteria in biotechnology. J. Appl. Microbiol. 2009, 106, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Dobretsov, S.; Abed, R.; Teplitski, M. Mini-Review: Inhibition of biofouling by marine microorganisms. Biofouling 2013, 29, 423–441. [Google Scholar] [CrossRef] [PubMed]
- Satheesh, S.; Ba-akdah, M.A.; Al-Sofyani, A.A. Natural antifouling compound production by microbes associated with marine macroorganisms—A review. Electron. J. Biotechnol. 2016, 21, 26–35. [Google Scholar] [CrossRef]
- Chapman, J.; Hellio, C.; Sullivan, T.; Brown, R.; Russell, S.; Kiterringham, E.; Le Nor, L.; Regan, F. Bioinspired synthetic macroalgae: Examples from nature for antifouling applications. Int. Biodeterior. Biodegrad. 2014, 86, 6–13. [Google Scholar] [CrossRef]
- Prabhakaran, S.; Rajaram, R.; Balasubramanian, V.; Mathivanan, K. Antifouling potentials of extracts from seaweeds, seagrasses and mangroves against primary biofilm forming bacteria. Asian Pac. J. Trop. Biomed. 2012, 2, 316–322. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, H.; Guo, G.; Pu, Y.; Yan, B.; Wang, C. Green alga Ulva pertusa—A new source of bioactive compounds with antialgal activity. Environ. Sci. Pollut. Res. 2015, 22, 10351–10359. [Google Scholar]
- Águila-Ramírez, R.N.; Arenas-González, A.; Hernández-Guerrero, C.J.; González-Acosta, B.; Borges-Souza, J.M.; Véron, B.; Pope, J.; Hellio, C. Antimicrobial and antifouling activities achieved by extracts of seaweeds from Gulf of California, Mexico. Hidrobiologica 2012, 22, 8–15. [Google Scholar]
- Batista, D.; Carvalho, A.P.; Costa, R.; Coutinho, R.; Dobretsov, S. Extracts of macroalgae from the Brazilian coast inhibit bacterial quorum sensing. Bot. Mar. 2014, 57, 441–447. [Google Scholar] [CrossRef]
- Grosser, K.; Zedler, L.; Schmitt, M.; Dietzek, B.; Popp, J.; Pohnert, G. Disruption-free imaging by Raman spectroscopy reveals a chemical sphere with antifouling metabolites around macroalgae. Biofouling 2012, 28, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Smyrniotopoulos, V.; Abatis, D.; Tziveleka, L.A.; Tsitsimpikou, C.; Roussis, V.; Loukis, A.; Vagias, C. Acetylene sesquiterpenoid esters from the green alga Caulerpa prolifera. J. Nat. Prod. 2003, 66, 21–24. [Google Scholar] [CrossRef] [PubMed]
- Bhagavathy, S.; Sumathi, P.; Bell, I.J.S. Green algae Chlorococcum humicola—A new source of bioactive compounds with antimicrobial activity. Asian Pac. J. Trop. Biomed. 2011, 1, S1–S7. [Google Scholar] [CrossRef]
- Saha, M.; Wahl, M. Seasonal variation in the antifouling defence of the temperate brown alga Fucus vesiculosus. Biofouling 2013, 29, 661–668. [Google Scholar] [CrossRef] [PubMed]
- Lachnit, T.; Fischer, M.; Künzel, S.; Baines, J.F.; Harder, T. Compounds associated with algal surfaces mediate epiphytic colonization of the marine macroalga Fucus vesiculosus. FEMS Microbiol. Ecol. 2013, 84, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Lachnit, T.; Wahl, M.; Harder, T. Isolated thallus-associated compounds from the macroalga Fucus vesiculosus mediate bacterial surface colonization in the field similar to that on the natural alga. Biofouling 2010, 26, 245–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saha, M.; Rempt, M.; Gebser, B.; Grueneberg, J.; Pohnert, G.; Weinberger, F. Biofouling Dimethylsulphopropionate (DMSP) and proline from the surface of the brown alga Fucus vesiculosus inhibit bacterial attachment. Biofouling 2012, 28, 593–604. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, N.; Sugimoto, N.; Ohki, K.; Kamiya, M. Diversity of phlorotannin profiles among sargassasacean species affecting variation and abundance of epiphytes. Eur. J. Phycol. 2016, 51, 307–316. [Google Scholar] [CrossRef]
- Plouguerné, E.; Hellio, C.; Cesconetto, C.; Thabard, M.; Mason, K.; Véron, B.; Pereira, R.C.; da Gama, B.A.P. Antifouling activity as a function of population variation in Sargassum vulgare from the littoral of Rio de Janeiro (Brazil). J. Appl. Phycol. 2010, 22, 717–724. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.X.; Wu, H.X.; Xu, Y.; Shao, C.L.; Wang, C.Y.; Qian, P.Y. Antifouling activity of secondary metabolites isolated from Chinese marine organisms. Mar. Biotechnol. 2013, 15, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.Y. Antifouling chromanols isolated from brown alga Sargassum horneri. J. Appl. Phycol. 2013, 25, 299–309. [Google Scholar] [CrossRef]
- Schwartz, N.; Dobretsov, S.; Rohde, S.; Schupp, P.J. Comparison of antifouling properties of native and invasive Sargassum (Fucales, Phaeophyceae) species. Eur. J. Phycol. 2017, 52, 116–131. [Google Scholar] [CrossRef]
- Silkina, A.; Bazes, A.; Mouget, J.-L.; Bourgougnon, N. Comparative efficiency of macroalgal extracts and booster biocides as antifouling agents to control growth of three diatom species. Mar. Pollut. Bull. 2012, 64, 2039–2046. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.P.; Batista, D.; Dobretsov, S.; Coutinho, R. Extracts of seaweeds as potential inhibitors of quorum sensing and bacterial growth. J. Appl. Phycol. 2016. [Google Scholar] [CrossRef]
- Othmani, A.; Briand, J.F.; Ayé, M.; Molmeret, M.; Culioli, G. Surface metabolites of the brown alga Taonia atomaria have the ability to regulate epibiosis. Biofouling 2016, 32, 801–813. [Google Scholar] [CrossRef] [PubMed]
- Othmani, A.; Bunet, R.; Bonnefont, J.-L.; Briand, J.-F.; Culioli, G. Settlement inhibition of marine biofilm bacteria and barnacle larvae by compounds isolated from the Mediterranean brown alga Taonia atomaria. J. Appl. Phycol. 2016, 28, 1975–1986. [Google Scholar] [CrossRef]
- Suresh, M.; Iyapparaj, P.; Anantharaman, P. Optimization, characterization and partial purification of bacteriocin produced by Staphylococcus haemolyticus MSM an isolate from seaweed. Biocatal. Agric. Biotechnol. 2014, 3, 161–166. [Google Scholar] [CrossRef]
- El Hattab, M.; Genta-jouve, G.; Bouzidi, N.; Orthalo-magn, M.; Hellio, C.; Marechal, J.P. Cystophloroketals A-E, unusual phloroglucinol-Meroterpenoid hybrids from the brown alga Cystoseira tamariscifolia. J. Nat. Prod. 2015, 78, 1663–1670. [Google Scholar] [CrossRef] [PubMed]
- Busetti, A.; Thompson, T.P.; Tegazzini, D.; Megaw, J.; Maggs, C.A.; Gilmore, B.F. Antibiofilm activity of the brown alga Halidrys siliquosa against clinically relevant human pathogens. Mar. Drugs 2015, 13, 3581–3605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Othmani, A.; Bouzidi, N.; Viano, Y.; Alliche, Z.; Seridi, H.; Blache, Y.; El Hattab, M.; Briand, J.-F.; Culioli, G. Anti-microfouling properties of compounds isolated from several Mediterranean Dictyota spp. J. Appl. Phycol. 2014, 26, 1573–1584. [Google Scholar] [CrossRef]
- Ktari, L.; Ismail-ben, A.A.; Ben, R.Y.; Langar, H.; El Bour, M. Antifouling activity and chemical investigation of the brown alga Dictyota fasciola (Dictyotales) from Tunisian coast. Cah. Biol. Mar. 2010, 51, 109–115. [Google Scholar]
- Muñoz, J.; Culioli, G.; Köck, M. Linear diterpenes from the marine brown alga Bifurcaria bifurcata: A chemical perspective. Phytochem. Rev. 2013, 12, 407–424. [Google Scholar] [CrossRef]
- Paradas, W.C.; Salgado, L.T.; Pereira, R.C.; Hellio, C.; Atella, G.C.; de Lima Moreira, D.; do Carmo, A.P.B.; Soares, A.R.; Amado-Filho, G.M. A Novel Antifouling Defense Strategy from Red Seaweed: Exocytosis and Deposition of Fatty Acid Derivatives at the Cell Wall Surface. Plant Cell Physiol. 2016, 57, 1008–1019. [Google Scholar] [CrossRef] [PubMed]
- Al-Lihaibi, S.S.; Abdel-Lateff, A.; Alarif, W.M.; Nogata, Y.; Ayyad, S.E.N.; Okino, T. Potent Antifouling Metabolites from Red Sea Organisms. Asian J. Chem. 2015, 27, 2252–2256. [Google Scholar] [CrossRef]
- Umezawa, T.; Oguri, Y.; Matsuura, H.; Yamazaki, S.; Suzuki, M.; Yoshimura, E.; Furuta, T.; Nogata, Y.; Serisawa, Y.; Matsuyama-Serisawa, K.; et al. Omaezallene from red alga Laurencia sp.: Structure elucidation, total synthesis, and antifouling activity. Angew. Chem. Int. Ed. 2014, 53, 3909–3912. [Google Scholar] [CrossRef] [PubMed]
- Cen-Pacheco, F.; Santiago-Benítez, A.J.; García, C.; Álvarez-Méndez, S.J.; Martín-Rodríguez, A.J.; Norte, M.; Martín, V.S.; Gavín, J.A.; Fernández, J.J.; Daranas, A.H. Oxasqualenoids from Laurencia viridis: Combined Spectroscopic–Computational Analysis and Antifouling Potential. J. Nat. Prod. 2015, 78, 712–721. [Google Scholar] [CrossRef] [PubMed]
- Gomez-Lemos, L.A.; Diaz-Pulido, G. Crustose coralline algae and associated microbial biofilms deter seaweed settlement on coral reefs. Coral Reefs 2017, 36, 453–462. [Google Scholar] [CrossRef]
- Martinez-Garcia, M.; van der Maarel, M.J.E.C. Floridoside production by the red microalga Galdieria sulphuraria under different conditions of growth and osmotic stress. AMB Express 2016, 6, 2–8. [Google Scholar] [CrossRef] [PubMed]
- Jha, B.; Kavita, K.; Westphal, J.; Hartmann, A.; Schmitt-Kopplin, P. Quorum sensing inhibition by Asparagopsis taxiformis, a marine macroalga: Separation of the compound that interrupts bacterial communication. Mar. Drugs 2013, 11, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Manilal, A.; Sujith, S.; Sabarathnam, B.; Kiran, G.S.; Selvin, J.; Shakir, C.; Lipton, A. Antifouling potentials of seaweeds collected from the Southwest Coast of India. World J. Agric. Sci. 2010, 6, 243–248. [Google Scholar]
- Chambers, L.D.; Stokes, K.R.; Walsh, F.C.; Wood, R.J.K. Modern approaches to marine antifouling coatings. Surf. Coat. Technol. 2006, 201, 3642–3652. [Google Scholar] [CrossRef]
- Williams, P.; Camara, M.; Hardman, A.; Swift, S.; Milton, D.; Hope, V.J.; Winzer, K.; Middleton, B.; Pritchard, D.I.; Bycroft, B.W. Quorum sensing and the population-dependent control virulence. Philos. Trans. Biol. Sci. 2000, 355, 667–680. [Google Scholar] [CrossRef] [PubMed]
- Dobretsov, S.; Teplitski, M.; Paul, V.J. Mini-Review: Quorum sensing in the marine environment and its relationship to biofouling. Biofouling 2009, 25, 413–427. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Kim, Y.H.; Seo, Y.W.; Park, S. Quorum sensing inhibitors from the red alga, Ahnfeltiopsis flabelliformis. Biotechnol. Bioprocess Eng. 2007, 12, 308–311. [Google Scholar] [CrossRef]
- Goecke, F.; Labes, A.; Wiese, J.; Imhoff, J.F. Chemical interactions between marine macroalgae and bacteria. Mar. Ecol. Prog. Ser. 2010, 409, 267–300. [Google Scholar] [CrossRef]
- Saurav, K.; Costantino, V.; Venturi, V.; Steindler, L. Quorum Sensing Inhibitors from the Sea Discovered Using Bacterial N-acyl-homoserine Lactone-Based Biosensors. Mar. Drugs 2017, 15, 53. [Google Scholar] [CrossRef] [PubMed]
- Borchardt, S.A.; Allian, E.J.; Michels, J.J.; Stearns, G.W.; Kelly, R.F.; McCoy, W.F. Reaction of acylated homoserine lactone bacterial signaling molecules with oxidized halogen antimicrobials. Appl. Environ. Microbiol. 2001, 67, 3174–3179. [Google Scholar] [CrossRef] [PubMed]
- Dobretsov, S.; Teplitski, M.; Bayer, M.; Gunasekera, S.; Proksch, P.; Paul, V. Inhibition of marine biofouling by bacterial quorum sensing inhibitors. Biofouling 2011, 27, 893–905. [Google Scholar] [CrossRef] [PubMed]
- Kanagasabhapathy, M.; Yamazaki, G.; Ishida, A.; Sasaki, H.; Nagata, S. Presence of quorum-sensing inhibitor-like compounds from bacteria isolated from the brown alga Colpomenia sinuosa. Lett. Appl. Microbiol. 2009, 49, 573–579. [Google Scholar] [CrossRef] [PubMed]
- Pugazhendhi, E.; Kirubha, P.; Palanisamy, K.; Gopalakrishnan, R. Synthesis and characterization of silver nanoparticles from Alpinia calcarata by Green approach and its applications in bactericidal and nonlinear optics. Appl. Surf. Sci. 2015, 357, 1801–1808. [Google Scholar] [CrossRef]
- Shankar, P.D.; Shobana, S.; Karuppusamy, I.; Pugazhendhi, A.; Ramkumar, V.C.; Arvindnarayan, S.; Kumar, G. A review on the biosynthesis of metallic nanoparticles (gold and silver) using bio-components of microalgae: Formation mechanism and applications. Enzyme Microb. Technol. 2016, 95, 28–44. [Google Scholar] [CrossRef] [PubMed]
- Al Naamani, L.; Dobretsov, S.; Dutta, J.; Burgess, G. Chitosan-ZnO nanocomposite coatings for the prevention of marine fouling. Chemosphere 2017, 168, 408–417. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-L.; Li, Y.-F.; Liang, X.; Guo, X.-P.; Ding, D.-W.; Zhang, D.; Zhou, S.; Bao, W.-Y.; Bellou, N.; Dobretsov, S. Silver nanoparticles impact biofilm communities and mussel settlement. Sci. Rep. 2016, 6, 37406. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.-L.; Li, Y.-F.; Guo, X.-P.; Liang, X.; Xu, Y.F.; Ding, D.W.; Bao, W.-Y.; Dobretsov, S. The effect of carbon nanotubes and titanium dioxide incorporated in PDMS on biofilm community composition and subsequent mussel plantigrade settlement. Biofouling 2016, 32, 763–777. [Google Scholar] [CrossRef] [PubMed]
- Sathe, P.; Myint, M.T.Z.; Dobretsov, S.; Dutta, J. Self-decontaminating photocatalytic zinc oxide nanorod coatings for prevention of marine microfouling: A mesocosm study. Biofouling 2016, 32, 383–395. [Google Scholar] [CrossRef] [PubMed]
- Muthukumar, K.; Vignesh, S.; Dahms, H.-U.; Gokul, M.S.; Palanichamy, X.; Subramian, G.; James, R.A. Antifouling assessments on biogenic nanoparticles: A field study from polluted offshore platform. Mar. Pollut. Bull. 2015, 101, 816–825. [Google Scholar]
- Asmathunisha, N.; Kathiresan, K. A review on biosynthesis of nanoparticles by marine organisms. Coll. Surf. B Biointerfaces 2013, 103, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Thakkar, K.N; Mhatre, S.S.; Parikh, R.Y. Biological synthesis of metal nanoparticles. Nanomed. Nanotechnol. Biol. Med. 2010, 6, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Ramkumar, V.R.; Pugazhendhi, A.; Gopalakrishnan, K.; Sivagurunathan, P.; Saratale, G.D. Biofabrication and characterization of silver nanoparticles using aqueous extracts of seaweed Enteromorpha compressa and its biomedical properties. Biotechnol. Rep. 2017. [Google Scholar] [CrossRef] [PubMed]
- Stachowicz, J.J. Mutualism, facilitation and the structure of ecological communities. BioScience 2001, 51, 235–246. [Google Scholar] [CrossRef]
- Dworjanyn, S.A.; Wright, J.T.; Paul, N.A.; De Nys, R.; Steinberg, P.D. Cost of chemical defence in the red alga Delisea pulchra. Oikos 2006, 113, 13–22. [Google Scholar] [CrossRef]
- Harizani, M.; Ioannou, E.; Roussis, V. The Laurencia paradox: An endless source of chemodiversity. In Progress in the Chemistry of Organic Natural Products; Kinghorn, A.D., Galk, H., Gibbons, S., Kobayashi, J., Eds.; Springer: Berlin, Germany, 2016; Volume 102, pp. 91–252. [Google Scholar]
- Paradas, W.C.; Crespo, T.M.; Saldagi, L.T.; de Andrade, L.R.; Soares, A.R.; Hellio, C.; Paranhos, R.R.; Hill, L.J.; de Souza, G.M.; Kelecom, A.G.; et al. Mevalonosomes: Specific vacuoles containing the mevalonate pathway in Plocamium brasiliense cortical cells (Rhodophyta). J. Phycol. 2015, 51, 225–235. [Google Scholar] [CrossRef] [PubMed]
- Hellio, C.; Yebra, D.M. Advances in Marine Antifouling Coating and Technologies, 1st ed.; Woodhead publishing Limited: Boca Raton, FL, USA, 2009; p. 784. [Google Scholar]
- Da Gama, B.A.; Plouguerne, E.; Pereira, R.C. The antifouling defence mechanisms of marine macroalgae. Adv. Bot. Res. 2014, 71, 413–440. [Google Scholar]
- Dobretsov, S. Marine Biofilms. In Biofouling, 1st ed.; Dürr, S., Thomason, J.C., Eds.; Wiley-Blackwell: Oxford, UK, 2010; pp. 123–136. [Google Scholar]
- Wade, W. Unculturable bacteria—The uncharacterized organisms that cause oral infections. J. R. Soc. Med. 2002, 95, 81–83. [Google Scholar] [CrossRef] [PubMed]
- Nylund, G.M.; Gribben, P.E.; de Nys, R.; Steinberg, P.; Pavia, H. Surface chemistry versus whole-cell extracts: Antifouling tests with seaweeds metabolites. Mar. Ecol. Prog. Ser. 2007, 329, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Cirri, E.; Grosser, K.; Pohnert, G. A solid phase extraction based non-disruptive sampling technique to investigate the surface chemistry of macroalgae. Biofouling 2016, 32, 145–153. [Google Scholar] [CrossRef] [PubMed]
- Holmstrøm, C.; Kjelleberg, S. Marine Pseudoalteromonas species are associated with higher organisms and produce active extracellular compounds. FEMS Microbiol. Ecol. 1999, 30, 285–293. [Google Scholar] [CrossRef]
- Burgess, J.G.; Boyd, K.G.; Armstrong, E.; Jiang, Z.; Yan, L.; Berggren, M.; May, U.; Pisacane, T.; Granmo, A.; Adams, D.R. The development of a marine natural product-based antifouling paint. Biofouling 2003, 19, 197–205. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, A.L.L.; de Felício, R.; Debonsi, H.M. Marine natural products: Chemical and biological potential of seaweeds and their endophytic fungi. Braz. J. Pharm. 2012, 22, 906–920. [Google Scholar] [CrossRef]
- Dobretsov, S.; Qian, P.Y. Effect of bacteria associated with the green alga Ulva reticulata on marine micro- and macrofouling. Biofouling 2002, 18, 217–228. [Google Scholar] [CrossRef]
- Harder, T.; Dobretsov, S.; Qian, P.Y. Waterborne polar macromolecules act as algal antifoulants in the seaweed Ulva reticulata. Mar. Ecol. Prog. Ser. 2004, 274, 133–141. [Google Scholar] [CrossRef]
- Kientz, B.; Thabard, M.; Cragg, S.M.; Pope, J.; Hellio, C. A new method for removing microflora from macroalgal surfaces: An important step for natural product discovery. Bot. Mar. 2011, 54, 2655–2661. [Google Scholar] [CrossRef]
- Yan, L.; Boyd, K.G.; Burgess, J.G. Surface attachment induced production of antimicrobial compounds by marine epiphytic bacteria using modified roller bottle cultivation. Mar. Biotechnol. 2009, 4, 356–366. [Google Scholar] [CrossRef] [PubMed]
- Hellio, C.; Berge, J.P.; Beapoil, C.; Le Gal, Y.; Bourgougnon, N. Screening of marine algal extracts for anti-settlement activities against microalgae and macroalgae. Biofouling 2002, 18, 205–215. [Google Scholar] [CrossRef]
- Da Gama, B.A.P.; Carvalho, A.; Weidner, K.; Soared, A.R.; Coutinho, R.; Fleury, B.G.; Texeira, V.L.; Pereira, R.C. Antifouling activity of natural products from Brazilian seaweeds. Bot. Mar. 2008, 51, 191–201. [Google Scholar] [CrossRef]
- Kremb, S.; Müller, C.; Schmitt-Kopplin, P.; Voolstra, C.R. Bioactive Potential of Marine Macroalgae from the Central Red Sea (Saudi Arabia) Assessed by High-Throughput Imaging-Based Phenotypic Profiling. Mar. Drugs 2017, 15, 18. [Google Scholar] [CrossRef] [PubMed]
- Dahms, H.-U.; Won, E.-J.; Kim, H.-S.; Han, J.H.; Park, H.G.; Souissi, S.; Raisuddin, S.; Lee, J.-S. Potential of the small cyclopoid copepod Paracyclopina nana as an invertebrate model for ecotoxicity testing. Aquat. Toxicol. 2016, 180, 282–294. [Google Scholar] [CrossRef] [PubMed]
- Dahms, H.-U.; Huang, D.-J.; Lee, S.H.; Chen, W.-Y.; Soong, K.; Hwang, J.-S. The challenging role of life cycle monitoring—Evidence from bisphenol A on the copepod Tigriopus japonicus. Hydrobiologia 2017, 784, 81–91. [Google Scholar] [CrossRef]
- Dahms, H.-U. Traditional microbiological methods. In Biofouling Methods, 1st ed.; Dobretsov, S., Thomason, J.C., Williams, D.N., Eds.; John Wiley & Sons: Oxford, UK, 2014; pp. 123–136. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
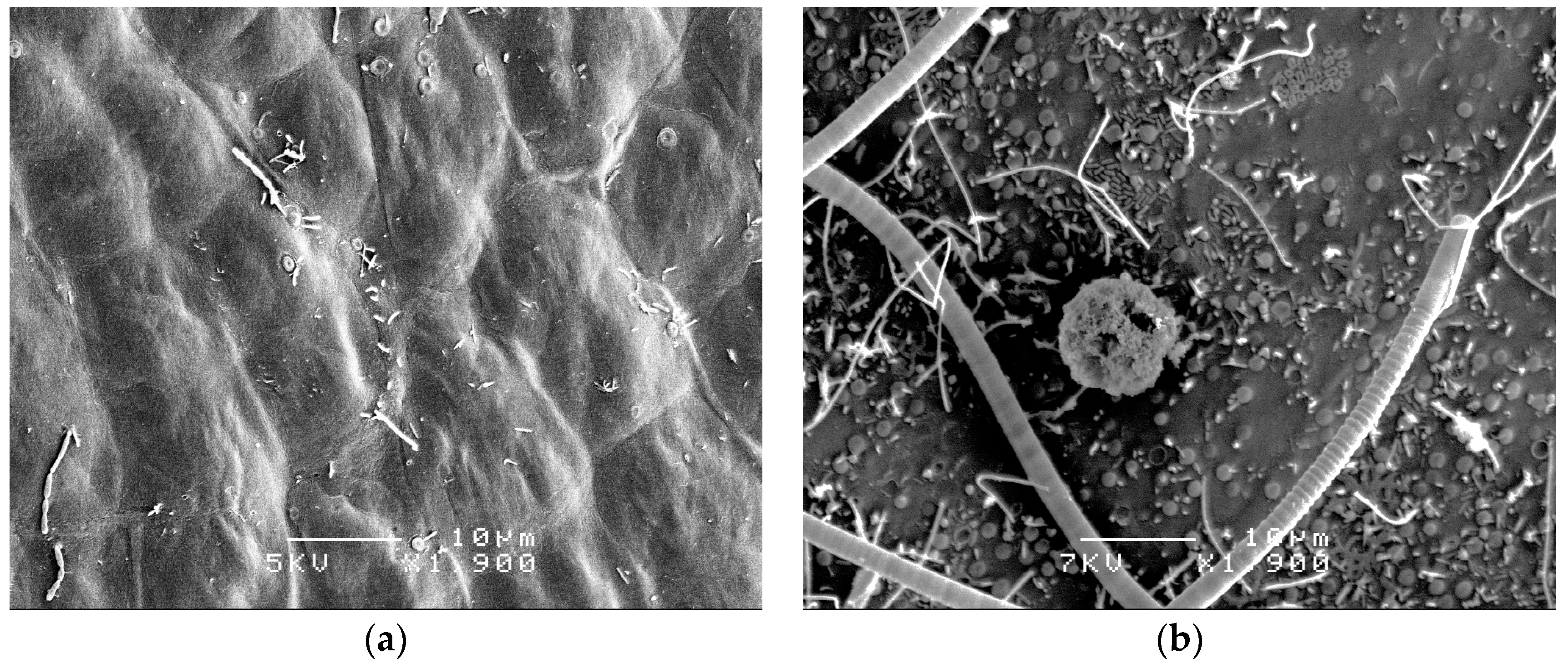{kind=link}
| Algae | Type of Activity | Compound | Reference 1 |
|---|---|---|---|
| Ulva rigida | Antifouling | 3-bromo-5-(diphenylene)-2(5H)-furanone | [17] |
| Ulva pertusa | Anti-algal | Alkaloid phenolic acid | [19] |
| Ulva reticulate Halimeda macroloba | Anti-bacterial | Organic extract 2 | [18] |
| Ulva lactuca | Anti-bacterial | Organic extract 2 | |
| Codium fragile | Anti-algal | Organic extract 2 | [20] |
| Caulerpa racemosa Codium spp. Ulva (Enteromorpha) fasciata | Anti-QS, Anti-bacterial | Polar and non-polar extracts 2 | [21] |
| Ulva sp. | Antifouling | β-carotene | [22] |
| Caulerpa prolifera | Antibacterial, antialgal | Acetylene sesquiterpenoid esters | [23] |
| Chlorococcum humicola | Antibacterial and anti Aspergillus | Chlorophyll a and b | [24] |
| Algae | Type of Activity | Compound | Reference 1 |
|---|---|---|---|
| Native and invasive Sargassum spp. | Anti-QS Anti-larval Anti-diatom | Non-polar extracts 2 | [33] |
| Sargassum spp. | Anti-algal | Phlorotannin | [29] |
| Sargassum muticum | Anti-diatom | Ethanol and Dichlormethane extracts 2 | [34] |
| S. muticum | Anti-bacterial | Galactoglycerolipids | [30] |
| S. thunbergii | Anti-larval | Stigmasta-5,22-E-,28-triene-3β,24α-diol | [31] |
| S. horneri | Anti-bacterial Anti-larval Anti-diatom | Chromanols | [32] |
| S. vulgare Padina sp. | Anti-QS Anti-bacterial | Polar and non-polar extracts 2 | [35] |
| S. furcatum Dyctiota sp. | Anti-QS Anti-bacterial | Polar and non-polar extracts 2 | [21] |
| Taonia atomaria | Anti-bacterial | Sesquiterpenoids | [36] |
| Taonia atomaria | Anti-bacterial Anti-larval | Sesquiterpenoid (−)-gleenol sn-3-O-(geranylgeranyl)glycerol | [37] |
| Padina tetrastromatica | Anti-bacterial Anti-diatom Anti-mussel | Methanol extracts (fatty acids) 2 | [38] |
| Cystoseira tamariscifolia | Anti-bacterial Anti-fungal, Anti-larval, Anti-algal | Cystophloroketals A-E monocyclic meroditerpenoid | [39] |
| Halidrys siliquosa | Anti-bacterial | Methanolic extracts 2 | [40] |
| Dyctiota spp. | Anti-bacterial | Diterpenes 1-Ο-octadecenoylglycerol sn-3-Ο-(geranylgeranyl)glycerol | [41] |
| Dyctiota sp. | Anti-bacterial Anti-algal | Extract 2 | [42] |
| Bifurcaria bifurcata | Anti-bacterial Anti-fouling | Acyclic linear diterpenoids | [43] |
| Fucus vesiculosus | Anti-bacterial | Surface extracts 2 | [25] |
| F.vesiculosus | Anti-bacterial | Surface extracts 2 | [26] |
| F.vesiculosus | Anti-bacterial | Dimethylsulphopropionate Proline | [28] |
| Laurencia johnstonii | Anti-bacterial | Organic extract 2 | [20] |
| Algae | Type of Activity | Compound | Reference 1 |
|---|---|---|---|
| Crustose coralline algae | Anti-algal | Methanol extract 2 | [48] |
| Galdieria sulphuraria | Antifouling | Floridoside | [49] |
| Laurencia translucida | Anti-bacterial | Fatty acid derivates | [44] |
| Laurencia obtusa | Anti-larval | 2,10-dibromo-3-chloro-7-chamigrene 12-hydroxyisolaurene | [45] |
| Asparagopsis taxiformis | Anti-bacterial Anti-QS | Methanol extract 2-dodecanoyloxyethanesulfonate | [50] |
| A. taxiformis | Anti-bacterial Anti-algal | Extract 2 | [51] |
| Ceramium botryocarpum | Anti-diatom | Ethanol and Dichlormethane extracts 2 | [34] |
| Chondrus crispus | Anti-algal Anti-bacterial | Crude extracts 2 | [52] |
| Pterocladiella capillacea Spyridia aculeata | Anti-QS Anti-bacterial | Polar and non-polar extracts 2 | [35] |
| P. capillacea Peyssonnelia capensis Spyridia spp. | Anti-QS Anti-bacterial | Polar and non-polar extracts 2 | [21] |
| Laurencia sp. | Antifouling | Omaezallene | [46] |
| Laurencia translucida | Antifouling | Fatty acid | [44] |
| Laurencia viridis | Anti-diatom | Dehydrothyrsiferol | [47] |
| L. viridis | Anti-diatom | Saiyacenols B | [47] |
| L. viridis | Anti-diatom | Saiyacenols C | [47] |
| L. viridis | Anti-algal | 28-hydroxysaiyacenols B and A | [47] |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dahms, H.U.; Dobretsov, S. Antifouling Compounds from Marine Macroalgae. Mar. Drugs 2017, 15, 265. https://doi.org/10.3390/md15090265
Dahms HU, Dobretsov S. Antifouling Compounds from Marine Macroalgae. Marine Drugs. 2017; 15(9):265. https://doi.org/10.3390/md15090265
Chicago/Turabian StyleDahms, Hans Uwe, and Sergey Dobretsov. 2017. "Antifouling Compounds from Marine Macroalgae" Marine Drugs 15, no. 9: 265. https://doi.org/10.3390/md15090265
APA StyleDahms, H. U., & Dobretsov, S. (2017). Antifouling Compounds from Marine Macroalgae. Marine Drugs, 15(9), 265. https://doi.org/10.3390/md15090265