Sequence Analysis, Kinetic Constants, and Anion Inhibition Profile of the Nacrein-Like Protein (CgiNAP2X1) from the Pacific Oyster Magallana gigas (Ex-Crassostrea gigas)

 ,
,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
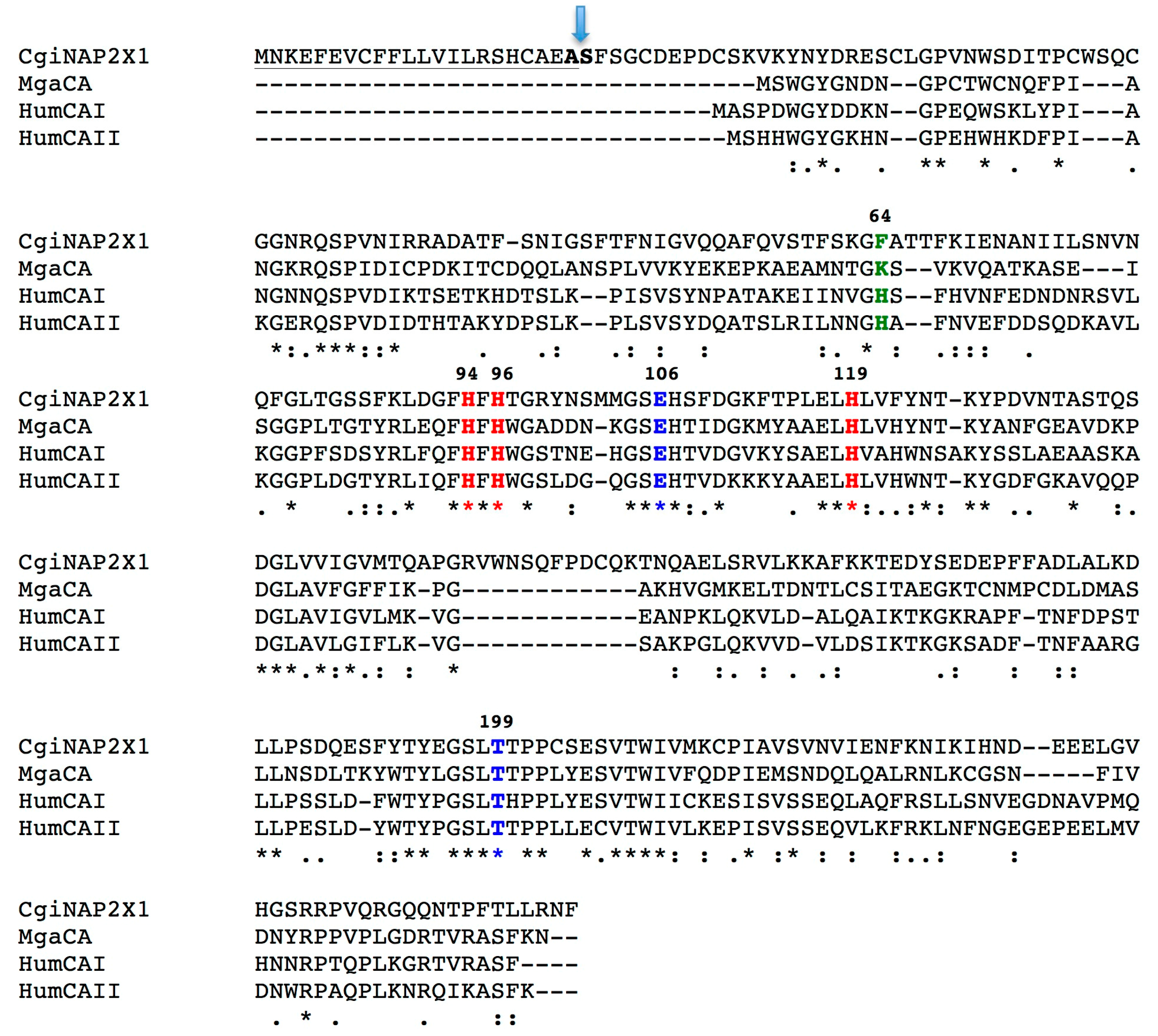
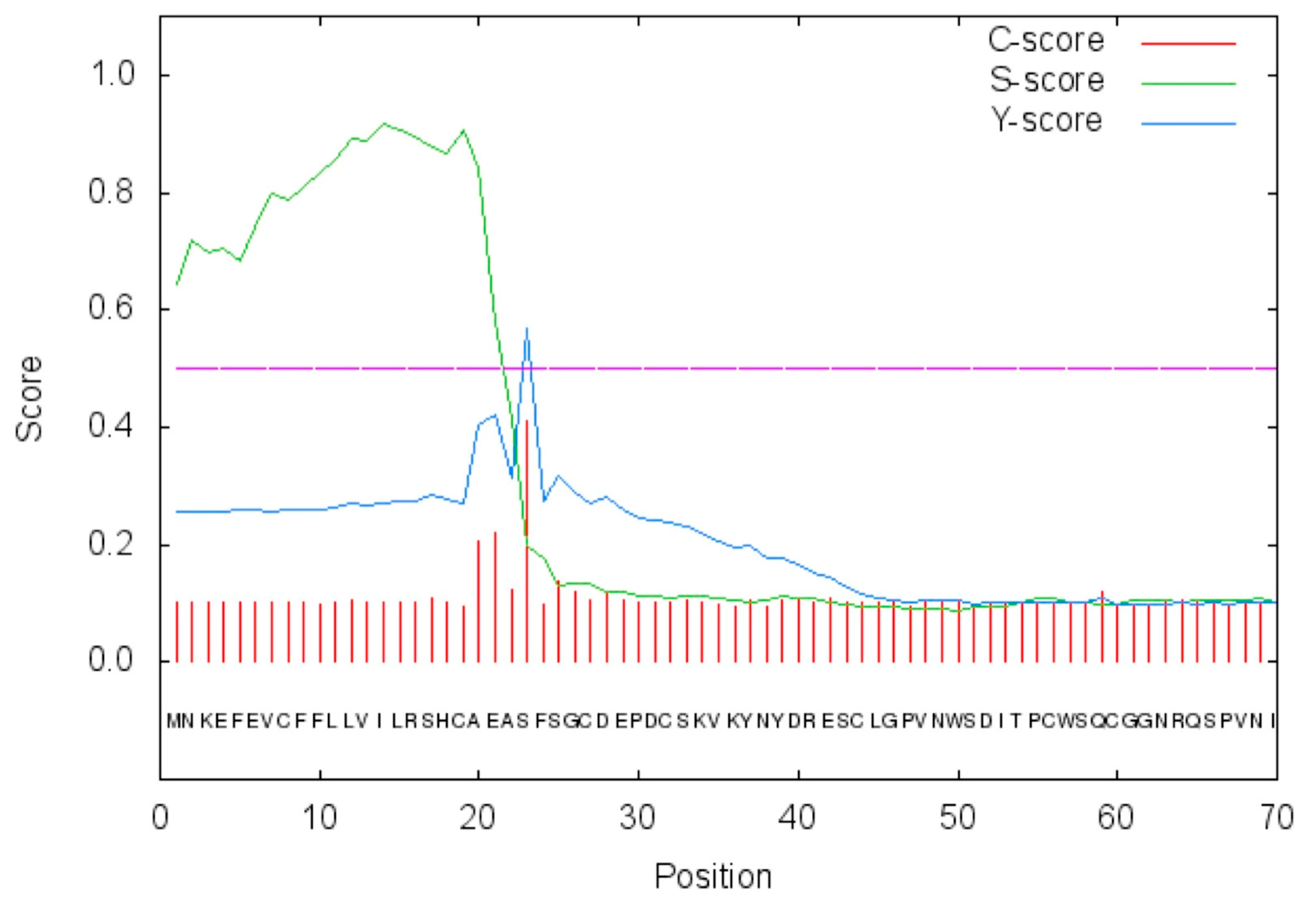
2.1. Amino Acid Sequence Analysis
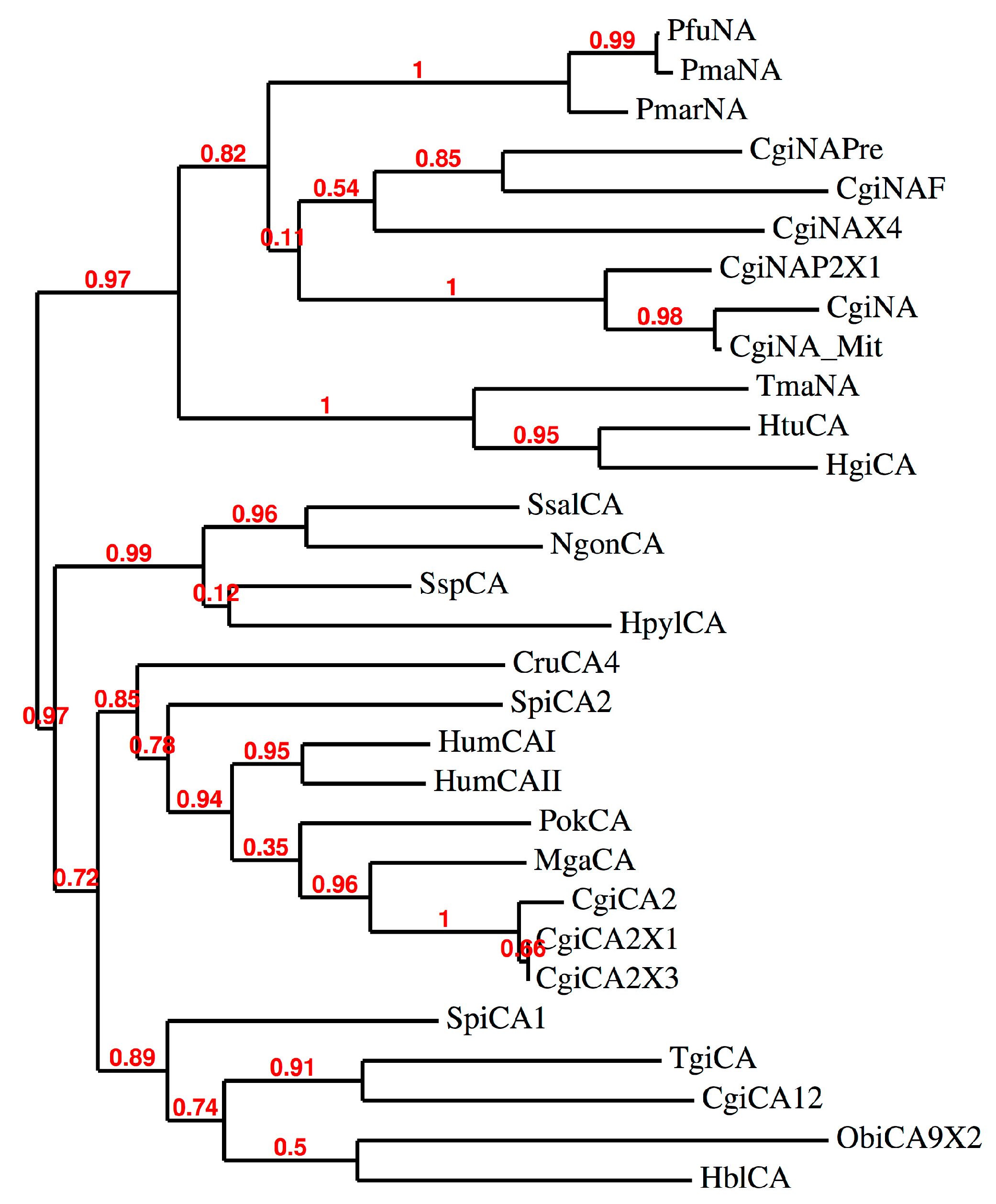
2.2. Phylogenetic Analysis
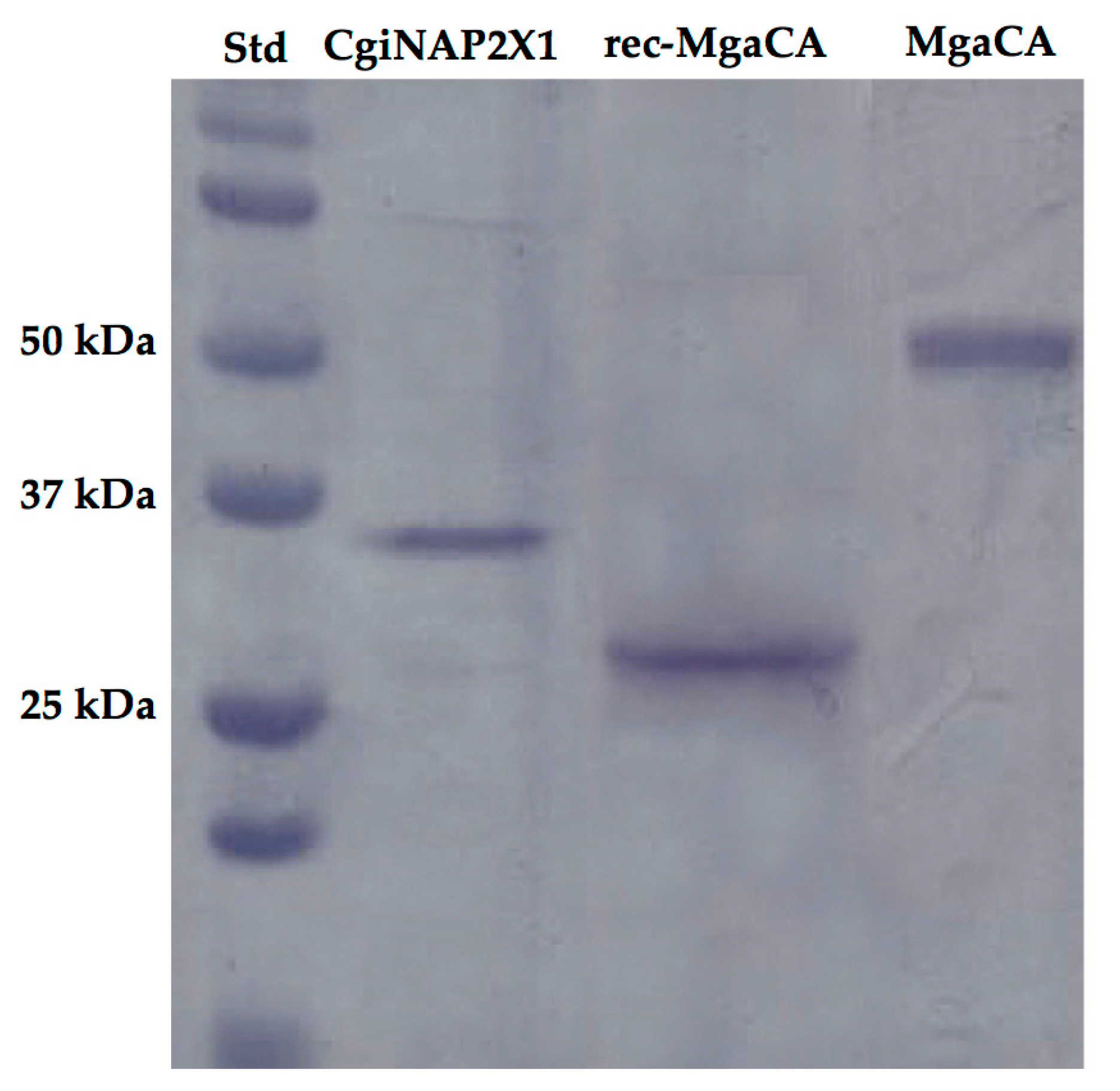
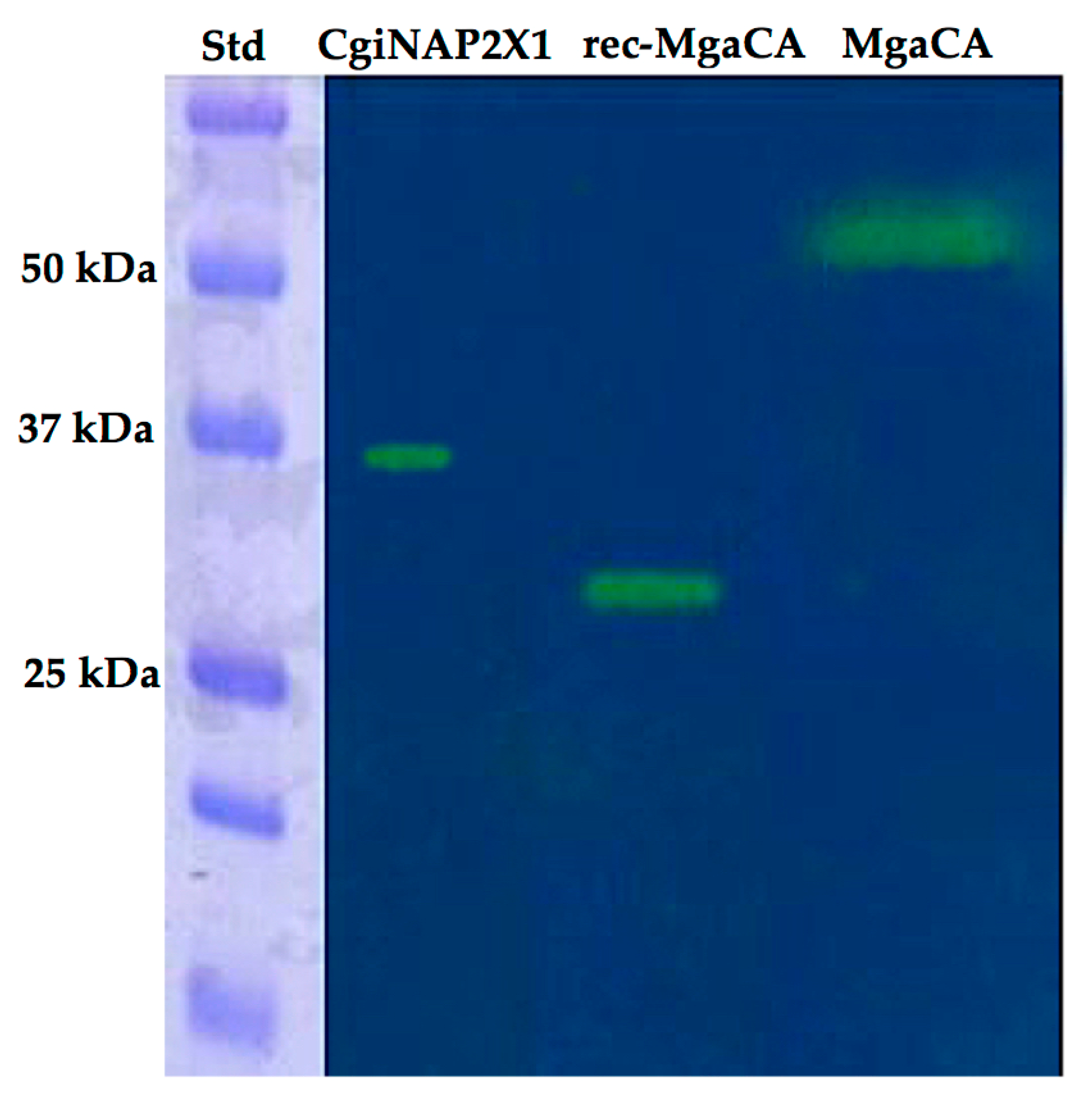
2.3. Production and Characterization of the Recombinant Enzyme (CgiNAP2X1)
2.4. Enzyme Kinetics
2.5. Anion Inhibition Profile
- The anions with the smallest propensity to coordinate metal ions, perchlorate and tetrafluoroborate, and inhibitors such as fluorosulfonate, iminodisulfonate, perrhuthenate, perrhenate, bisulfite, divanadate, tellurate, azide, and cyanide were not inhibitors of CgiNAP2X1 up to concentrations of 100 mM. With the exception of perchlorate and tetrafluoroborate, a different profile was shown for the human isoforms hCA I and II (Table 3).
- Several other anions; fluoride, chloride, bromide, iodide, cyanate, thiocyanate, bicarbonate, carbonate, nitrate, nitrite, hydrogen sulfite, stannate, selenate, peroxydisulfate, tetraborate, selenocyanate, trithiocarbonate, and trifluorometansolfonato were also weak inhibitors of CgiNAP2X1, with inhibition constants in the range of 3.1–20.7 mM (Table 3). Interesting to note is that most of these inhibitors were micro and/or submicromolar inhibitors for the human enzymes. It should be noted that the weak affinity of CgiNAP2X1 for bicarbonate and carbonate might be a positive feature for the physiological role of this enzyme in the deposition of the molluskan shell, since the enzyme is not appreciably inhibited in the presence of rather high concentrations of these two anions. Moreover, for the same reason, the enzyme could have also a potential biotechnological application in the CO2 capture processes.
- The best anion inhibitors of CgiNAP2X1 were diethyldithiocarbamate, sulfamide, sulfamate, phenylboronic acid, and phenylarsonic acid, which showed a micromolar affinity for this enzyme, with KIs in the range of 76–87 μM (Table 3). Diethyldithiocarbamate was recently shown to lead to highly potent inhibitors directed against many α- or β-CAs, and some X-ray crystal structures also revealed their binding mode, which is achieved by coordination to the metal ion via a negatively charged sulfur atom [47,48,49,50,51]. It is conceivable that a similar binding mode is achieved also within the CgiNAP2X1 active site. Sulfamide and sulfamate also coordinate (via an ionized nitrogen atom) to the Zn(II) ion within the hCA II active site, and probably the same inhibition mechanism is valid against the oyster enzyme [47,48,49,50,51].
- Most of these anions are weak, millimolar CgiNAP2X1 inhibitors probably because the enzyme is in contact with sea water, rich in many inorganic anions (chloride, sulfate, etc.) which may interfere with the enzyme’s activity. Similar behavior was observed earlier for other marine organism CAs [37].
3. Materials and Methods
3.1. Gene Identification, Synthesis, and Cloning
3.2. Protein Expression and Purification
3.3. SDS-PAGE and Protonography
3.4. Amino Acid Sequence and Phylogenetic Analysis
3.5. Enzyme Kinetics
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Supuran, C.T. Structure and function of carbonic anhydrases. Biochem. J. 2016, 473, 2023–2032. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, S.; Vullo, D.; Fisher, G.M.; Andrews, K.T.; Poulsen, S.A.; Capasso, C.; Supuran, C.T. Discovery of a new family of carbonic anhydrases in the malaria pathogen plasmodium falciparum-the eta-carbonic anhydrases. Bioorg. Med. Chem. Lett. 2014, 24, 4389–4396. [Google Scholar] [CrossRef] [PubMed]
- Kikutani, S.; Nakajima, K.; Nagasato, C.; Tsuji, Y.; Miyatake, A.; Matsuda, Y. Thylakoid luminal theta-carbonic anhydrase critical for growth and photosynthesis in the marine diatom phaeodactylum tricornutum. Proc. Natl. Acad. Sci. USA 2016, 113, 9828–9833. [Google Scholar] [CrossRef] [PubMed]
- Capasso, C.; Supuran, C.T. Bacterial, fungal and protozoan carbonic anhydrases as drug targets. Expert Opin. Ther. Targets 2015, 19, 1689–1704. [Google Scholar] [CrossRef]
- Capasso, C.; Supuran, C.T. An overview of the selectivity and efficiency of the bacterial carbonic anhydrase inhibitors. Curr. Med. Chem. 2015, 22, 2130–2139. [Google Scholar] [CrossRef] [PubMed]
- Capasso, C.; Supuran, C.T. An overview of the alpha-, beta- and gamma-carbonic anhydrases from bacteria: Can bacterial carbonic anhydrases shed new light on evolution of bacteria? J. Enzyme Inhib. Med. Chem. 2015, 30, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Capasso, C.; Supuran, C.T. An overview of the carbonic anhydrases from two pathogens of the oral cavity: Streptococcus mutans and porphyromonas gingivalis. Curr. Top. Med. Chem. 2016, 16, 2359–2368. [Google Scholar] [CrossRef] [PubMed]
- Ozensoy Guler, O.; Capasso, C.; Supuran, C.T. A magnificent enzyme superfamily: Carbonic anhydrases, their purification and characterization. J. Enzyme Inhib. Med. Chem. 2016, 31, 689–694. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T.; Capasso, C. New light on bacterial carbonic anhydrases phylogeny based on the analysis of signal peptide sequences. J. Enzyme Inhib. Med. Chem. 2016, 31, 1254–1260. [Google Scholar] [CrossRef]
- Del Prete, S.; Vullo, D.; De Luca, V.; Carginale, V.; Ferraroni, M.; Osman, S.M.; AlOthman, Z.; Supuran, C.T.; Capasso, C. Sulfonamide inhibition studies of the beta-carbonic anhydrase from the pathogenic bacterium vibrio cholerae. Bioorg. Med. Chem 2016, 24, 1115–1120. [Google Scholar] [CrossRef] [PubMed]
- Briganti, F.; Pierattelli, R.; Scozzafava, A.; Supuran, C.T. Carbonic anhydrase inhibitors. Part 37. Novel classes of carbonic anhydrase inhibitors and their interaction with the native and cobalt-substituted enzyme: Kinetic and spectroscopic investigations. Eur. J. Med. Chem. 1996, 31, 1001–1010. [Google Scholar] [CrossRef]
- Supuran, C.T.; Capasso, C. The eta-class carbonic anhydrases as drug targets for antimalarial agents. Expert Opin. Ther. Targets 2015, 19, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, S.; De Luca, V.; Supuran, C.T.; Capasso, C. Protonography, a technique applicable for the analysis of eta-carbonic anhydrase activity. J. Enzyme Inhib. Med. Chem. 2015, 30, 920–924. [Google Scholar] [CrossRef] [PubMed]
- De Simone, G.; Di Fiore, A.; Capasso, C.; Supuran, C.T. The zinc coordination pattern in the eta-carbonic anhydrase from plasmodium falciparum is different from all other carbonic anhydrase genetic families. Bioorg. Med. Chem. Lett. 2015, 25, 1385–1389. [Google Scholar] [CrossRef] [PubMed]
- De Luca, V.; Del Prete, S.; Supuran, C.T.; Capasso, C. Protonography, a new technique for the analysis of carbonic anhydrase activity. J. Enzyme Inhib. Med. Chem. 2015, 30, 277–282. [Google Scholar] [CrossRef] [PubMed]
- Alafeefy, A.M.; Abdel-Aziz, H.A.; Vullo, D.; Al-Tamimi, A.M.; Awaad, A.S.; Mohamed, M.A.; Capasso, C.; Supuran, C.T. Inhibition of human carbonic anhydrase isozymes I, II, IX and XII with a new series of sulfonamides incorporating aroylhydrazone-, [1,2,4]triazolo[3,4-b][1,3,4]thiadiazinyl- or 2-(cyanophenylmethylene)-1,3,4-thiadiazol-3(2H)-yl moieties. J. Enzyme Inhib. Med. Chem. 2015, 30, 52–56. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, S.; Vullo, D.; De Luca, V.; Carginale, V.; Osman, S.M.; AlOthman, Z.; Supuran, C.T.; Capasso, C. Cloning, expression, purification and sulfonamide inhibition profile of the complete domain of the eta-carbonic anhydrase from plasmodium falciparum. Bioorg. Med. Chem. Lett. 2016, 26, 4184–4190. [Google Scholar] [CrossRef] [PubMed]
- Buzas, G.M.; Supuran, C.T. The history and rationale of using carbonic anhydrase inhibitors in the treatment of peptic ulcers. In memoriam ioan puscas (1932–2015). J. Enzyme Inhib. Med. Chem. 2016, 31, 527–533. [Google Scholar] [CrossRef] [PubMed]
- Carta, F.; Supuran, C.T.; Scozzafava, A. Sulfonamides and their isosters as carbonic anhydrase inhibitors. Future Med. Chem. 2014, 6, 1149–1165. [Google Scholar] [CrossRef] [PubMed]
- Pinard, M.A.; Lotlikar, S.R.; Boone, C.D.; Vullo, D.; Supuran, C.T.; Patrauchan, M.A.; McKenna, R. Structure and inhibition studies of a type II beta-carbonic anhydrase psca3 from pseudomonas aeruginosa. Bioorg. Med. Chem. 2015, 23, 4831–4838. [Google Scholar] [CrossRef] [PubMed]
- Ferraroni, M.; Del Prete, S.; Vullo, D.; Capasso, C.; Supuran, C.T. Crystal structure and kinetic studies of a tetrameric type II beta-carbonic anhydrase from the pathogenic bacterium vibrio cholerae. Acta Crystallogr. D Biol. Crystallogr. 2015, 71, 2449–2456. [Google Scholar] [CrossRef] [PubMed]
- De Simone, G.; Monti, S.M.; Alterio, V.; Buonanno, M.; De Luca, V.; Rossi, M.; Carginale, V.; Supuran, C.T.; Capasso, C.; Di Fiore, A. Crystal structure of the most catalytically effective carbonic anhydrase enzyme known, sazca from the thermophilic bacterium sulfurihydrogenibium azorense. Bioorg. Med. Chem. Lett. 2015, 25, 2002–2006. [Google Scholar] [CrossRef] [PubMed]
- Zolnowska, B.; Slawinski, J.; Pogorzelska, A.; Chojnacki, J.; Vullo, D.; Supuran, C.T. Carbonic anhydrase inhibitors. Synthesis, and molecular structure of novel series n-substituted n′-(2-arylmethylthio-4-chloro-5-methylbenzenesulfonyl)guanidines and their inhibition of human cytosolic isozymes I and II and the transmembrane tumor-associated isozymes IX and XII. Eur. J. Med. Chem. 2014, 71, 135–147. [Google Scholar] [PubMed]
- De Luca, L.; Ferro, S.; Damiano, F.M.; Supuran, C.T.; Vullo, D.; Chimirri, A.; Gitto, R. Structure-based screening for the discovery of new carbonic anhydrase vii inhibitors. Eur. J. Med. Chem. 2014, 71, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Di Fiore, A.; Capasso, C.; De Luca, V.; Monti, S.M.; Carginale, V.; Supuran, C.T.; Scozzafava, A.; Pedone, C.; Rossi, M.; De Simone, G. X-ray structure of the first ‘extremo-alpha-carbonic anhydrase’, a dimeric enzyme from the thermophilic bacterium sulfurihydrogenibium yellowstonense yo3aop1. Acta Crystallogr. D Biol. Crystallogr. 2013, 69, 1150–1159. [Google Scholar] [CrossRef]
- Supuran, C.T. Structure-based drug discovery of carbonic anhydrase inhibitors. J. Enzyme Inhib. Med. Chem. 2012, 27, 759–772. [Google Scholar] [CrossRef]
- Supuran, C.T. Carbonic anhydrases—An overview. Curr. Pharm. Des. 2008, 14, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T. Advances in structure-based drug discovery of carbonic anhydrase inhibitors. Expert Opin. Drug Discov. 2017, 12, 61–88. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, A.; Mahon, B.P.; Cruzeiro, V.W.; Cornelio, B.; Laronze-Cochard, M.; Ceruso, M.; Sapi, J.; Rance, G.A.; Khlobystov, A.N.; Fontana, A.; et al. Structure-activity relationships of benzenesulfonamide-based inhibitors towards carbonic anhydrase isoform specificity. Chembiochem 2017, 18, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Alterio, V.; Langella, E.; Viparelli, F.; Vullo, D.; Ascione, G.; Dathan, N.A.; Morel, F.M.; Supuran, C.T.; De Simone, G.; Monti, S.M. Structural and inhibition insights into carbonic anhydrase cdca1 from the marine diatom thalassiosira weissflogii. Biochimie 2012, 94, 1232–1241. [Google Scholar] [CrossRef] [PubMed]
- Lomelino, C.L.; Mahon, B.P.; McKenna, R.; Carta, F.; Supuran, C.T. Kinetic and X-ray crystallographic investigations on carbonic anhydrase isoforms I, II, IX and XII of a thioureido analog of SLC-0111. Bioorg. Med. Chem. 2016, 24, 976–981. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T. How many carbonic anhydrase inhibition mechanisms exist? J. Enzyme Inhib. Med. Chem. 2016, 31, 345–360. [Google Scholar] [CrossRef] [PubMed]
- Aspatwar, A.; Tolvanen, M.E.; Ortutay, C.; Parkkila, S. Carbonic anhydrase related proteins: Molecular biology and evolution. Subcell. Biochem. 2014, 75, 135–156. [Google Scholar] [PubMed]
- Supuran, C.T. Carbonic anhydrases as drug targets—An overview. Curr. Top. Med. Chem. 2007, 7, 825–833. [Google Scholar] [CrossRef]
- Capasso, C.; Supuran, C.T. Sulfa and trimethoprim-like drugs—Antimetabolites acting as carbonic anhydrase, dihydropteroate synthase and dihydrofolate reductase inhibitors. J. Enzyme Inhib. Med. Chem. 2014, 29, 379–387. [Google Scholar] [CrossRef] [PubMed]
- Capasso, C.; Supuran, C.T. Anti-infective carbonic anhydrase inhibitors: A patent and literature review. Expert Opin Ther. Pat. 2013, 23, 693–704. [Google Scholar] [CrossRef]
- Perfetto, R.; Del Prete, S.; Vullo, D.; Sansone, G.; Barone, C.; Rossi, M.; Supuran, C.T.; Capasso, C. Biochemical characterization of the native alpha-carbonic anhydrase purified from the mantle of the mediterranean mussel, mytilus galloprovincialis. J. Enzyme Inhib. Med. Chem. 2017, 32, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, S.; Vullo, D.; Zoccola, D.; Tambutte, S.; Capasso, C.; Supuran, C.T. Kinetic properties and affinities for sulfonamide inhibitors of an alpha-carbonic anhydrase (cruca4) involved in coral biomineralization in the mediterranean red coral corallium rubrum. Bioorg. Med. Chem. 2017, 25, 3525–3530. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, H.; Miyashita, T.; Okushima, M.; Nakano, S.; Morita, T.; Matsushiro, A. A carbonic anhydrase from the nacreous layer in oyster pearls. Proc. Natl. Acad. Sci. USA 1996, 93, 9657–9660. [Google Scholar] [CrossRef]
- Yu, Z.; Xie, L.; Lee, S.; Zhang, R. A novel carbonic anhydrase from the mantle of the pearl oyster (pinctada fucata). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2006, 143, 190–194. [Google Scholar] [CrossRef] [PubMed]
- Petersen, T.N.; Brunak, S.; von Heijne, G.; Nielsen, H. Signalp 4.0: Discriminating signal peptides from transmembrane regions. Nat. Methods 2011, 8, 785–786. [Google Scholar] [CrossRef] [PubMed]
- Nishimori, I.; Minakuchi, T.; Onishi, S.; Vullo, D.; Scozzafava, A.; Supuran, C.T. Carbonic anhydrase inhibitors. DNA cloning, characterization, and inhibition studies of the human secretory isoform VI, a new target for sulfonamide and sulfamate inhibitors. J. Med. Chem. 2007, 50, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Tanc, M.; Carta, F.; Scozzafava, A.; Supuran, C.T. 6-substituted 1,2-benzoxathiine-2,2-dioxides are isoform-selective inhibitors of human carbonic anhydrases IX, XII and VA. Org. Biomol. Chem. 2015, 13, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, S.; De Luca, V.; De Simone, G.; Supuran, C.T.; Capasso, C. Cloning, expression and purification of the complete domain of the eta-carbonic anhydrase from plasmodium falciparum. J. Enzyme Inhib. Med. Chem. 2016, 31, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, S.; De Luca, V.; Iandolo, E.; Supuran, C.T.; Capasso, C. Protonography, a powerful tool for analyzing the activity and the oligomeric state of the gamma-carbonic anhydrase identified in the genome of porphyromonas gingivalis. Bioorg. Med. Chem. 2015, 23, 3747–3750. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, H.; Miyoshi, F.; Kohno, J. The carbonic anhydrase domain protein nacrein is expressed in the epithelial cells of the mantle and acts as a negative regulator in calcification in the mollusc pinctada fucata. Zool. Sci. 2005, 22, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Vullo, D.; Del Prete, S.; Osman, S.M.; Scozzafava, A.; Alothman, Z.; Supuran, C.T.; Capasso, C. Anion inhibition study of the beta-class carbonic anhydrase (pgicab) from the oral pathogen porphyromonas gingivalis. Bioorg. Med. Chem. Lett. 2014, 24, 4402–4406. [Google Scholar] [CrossRef] [PubMed]
- Nishimori, I.; Vullo, D.; Minakuchi, T.; Scozzafava, A.; Osman, S.M.; AlOthman, Z.; Capasso, C.; Supuran, C.T. Anion inhibition studies of two new beta-carbonic anhydrases from the bacterial pathogen legionella pneumophila. Bioorg. Med. Chem. Lett. 2014, 24, 1127–1132. [Google Scholar] [CrossRef] [PubMed]
- Maresca, A.; Vullo, D.; Scozzafava, A.; Supuran, C.T. Inhibition of the alpha- and beta-carbonic anhydrases from the gastric pathogen helycobacter pylori with anions. J. Enzyme Inhib. Med. Chem 2013, 28, 388–391. [Google Scholar] [CrossRef] [PubMed]
- Innocenti, A.; Scozzafava, A.; Supuran, C.T. Carbonic anhydrase inhibitors. Inhibition of transmembrane isoforms IX, XII, and XIV with less investigated anions including trithiocarbonate and dithiocarbamate. Bioorg. Med. Chem. Lett. 2010, 20, 1548–1550. [Google Scholar] [CrossRef] [PubMed]
- Scozzafava, A.; Mastrolorenzo, A.; Supuran, C.T. Arylsulfonyl-n,n-diethyl-dithiocarbamates: A novel class of antitumor agents. Bioorg. Med. Chem. Lett. 2000, 10, 1887–1891. [Google Scholar] [CrossRef]
- Khalifah, R.G. The carbon dioxide hydration activity of carbonic anhydrase I. Stop-flow kinetic studies on the native human isoenzymes B and C. J. Biol. Chem. 1971, 246, 2561–2573. [Google Scholar]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Supuran, C.T. Carbonic anhydrase inhibitors. Bioorg. Med. Chem. Lett. 2010, 20, 3467–3474. [Google Scholar] [CrossRef] [PubMed]
- Vullo, D.; De Luca, V.; Del Prete, S.; Carginale, V.; Scozzafava, A.; Capasso, C.; Supuran, C.T. Sulfonamide inhibition studies of the gamma-carbonic anhydrase from the antarctic bacterium pseudoalteromonas haloplanktis. Bioorg. Med. Chem. Lett. 2015, 25, 3550–3555. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Muscle: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed]
- Guindon, S.; Gascuel, O. A simple, fast, and accurate algorithm to estimate large phylogenies by maximum likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, S.; Vullo, D.; De Luca, V.; Carginale, V.; di Fonzo, P.; Osman, S.M.; AlOthman, Z.; Supuran, C.T.; Capasso, C. Anion inhibition profiles of alpha-, beta- and gamma-carbonic anhydrases from the pathogenic bacterium vibrio cholerae. Bioorg. Med. Chem. 2016, 24, 3413–3417. [Google Scholar] [CrossRef] [PubMed]
- Del Prete, S.; Vullo, D.; De Luca, V.; Carginale, V.; di Fonzo, P.; Osman, S.M.; AlOthman, Z.; Supuran, C.T.; Capasso, C. Anion inhibition profiles of the complete domain of the eta-carbonic anhydrase from plasmodium falciparum. Bioorg. Med. Chem 2016, 24, 4410–4414. [Google Scholar] [CrossRef] [PubMed]
- De Luca, V.; Vullo, D.; Del Prete, S.; Carginale, V.; Osman, S.M.; AlOthman, Z.; Supuran, C.T.; Capasso, C. Cloning, characterization and anion inhibition studies of a gamma-carbonic anhydrase from the antarctic bacterium colwellia psychrerythraea. Bioorg. Med. Chem. 2016, 24, 835–840. [Google Scholar] [CrossRef]
- Drake, J.L.; Mass, T.; Haramaty, L.; Zelzion, E.; Bhattacharya, D.; Falkowski, P.G. Proteomic analysis of skeletal organic matrix from the stony coral stylophora pistillata. Proc. Natl. Acad. Sci. USA 2013, 110, 3788–3793. [Google Scholar] [CrossRef] [PubMed]
- Le Goff, C.; Ganot, P.; Zoccola, D.; Caminiti-Segonds, N.; Allemand, D.; Tambutte, S. Carbonic anhydrases in cnidarians: Novel perspectives from the octocorallian corallium rubrum. PLoS ONE 2016, 11, e0160368. [Google Scholar] [CrossRef] [PubMed]
- Mass, T.; Drake, J.L.; Peters, E.C.; Jiang, W.; Falkowski, P.G. Immunolocalization of skeletal matrix proteins in tissue and mineral of the coral stylophora pistillata. Proc. Natl. Acad. Sci. USA 2014, 111, 12728–12733. [Google Scholar] [CrossRef] [PubMed]
- Bertucci, A.; Moya, A.; Tambutte, S.; Allemand, D.; Supuran, C.T.; Zoccola, D. Carbonic anhydrases in anthozoan corals—A review. Bioorg. Med. Chem 2013, 21, 1437–1450. [Google Scholar] [CrossRef] [PubMed]
- Bertucci, A.; Tambutte, S.; Supuran, C.T.; Allemand, D.; Zoccola, D. A new coral carbonic anhydrase in stylophora pistillata. Mar. Biotechnol. (NY) 2011, 13, 992–1002. [Google Scholar] [CrossRef] [PubMed]
- Le Roy, N.; Jackson, D.J.; Marie, B.; Ramos-Silva, P.; Marin, F. The evolution of metazoan α-carbonic anhydrases and their roles in calcium carbonate biomineralization. Front. Zool. 2014, 11, 75. [Google Scholar] [CrossRef] [Green Version]
- Bertucci, A.; Innocenti, A.; Zoccola, D.; Scozzafava, A.; Allemand, D.; Tambutte, S.; Supuran, C. Carbonic anhydrase inhibitors: Inhibition studies of a coral secretory isoform with inorganic anions. Bioorg. Med. Chem. Lett. 2009, 19, 650–653. [Google Scholar] [CrossRef] [PubMed]
- Bertucci, A.; Innocenti, A.; Zoccola, D.; Scozzafava, A.; Tambutte, S.; Supuran, C.T. Carbonic anhydrase inhibitors. Inhibition studies of a coral secretory isoform by sulfonamides. Bioorg. Med. Chem. 2009, 17, 5054–5058. [Google Scholar] [CrossRef] [PubMed]
- Zoccola, D.; Innocenti, A.; Bertucci, A.; Tambutté, E.; Supuran, C.T.; Tambutté, S. Coral Carbonic Anhydrases: Regulation by Ocean Acidification. Mar Drugs. 2016, 14, 109. [Google Scholar] [CrossRef] [PubMed]
- Moya, A.; Tambutté, S.; Bertucci, A.; Tambutté, E.; Lotto, S.; Vullo, D.; Supuran, C.T.; Allemand, D.; Zoccola, D. Carbonic anhydrase in the scleractinian coral Stylophora pistillata: Characterization, localization, and role in biomineralization. J. Biol. Chem. 2008, 283, 25475–25484. [Google Scholar] [CrossRef] [PubMed]
- Bertucci, A.; Zoccola, D.; Tambutte, S.; Vullo, D.; Supuran, C. Carbonic anhydrase activators. The first activation study of a coral secretory isoform with amino acids and amines. Bioorg. Med. Chem. 2010, 18, 2300–2303. [Google Scholar] [CrossRef] [PubMed]
- Allemand, D.; Tambutte, E.; Zoccola, D.; Tambutte, S. Coral calcification, cells to reefs. In Coral Reefs: An Ecosystem in Transition; Wiley: New York, NY, USA, 2011; pp. 119–150. [Google Scholar]
- Müller, W.E.; Wang, X.; Grebenjuk, V.A.; Korzhev, M.; Wiens, M.; Schlossmacher, U.; Schröder, H.C. Common genetic denominators for Ca++-based skeleton in Metazoa: Role of osteoclast-stimulating factor and of carbonic anhydrase in a calcareous sponge. PLoS ONE 2012, 7, e34617. [Google Scholar]
- Müller, W.E.; Schröder, H.C.; Schlossmacher, U.; Neufurth, M.; Geurtsen, W.; Korzhev, M.; Wang, X. The enzyme carbonic anhydrase as an integral component of biogenic Ca-carbonate formation in sponge spicules. FEBS Open Bio 2013, 3, 357–362. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Acronym | Enzyme Type | Organism | Accession Number |
|---|---|---|---|
| PfuNA | Nacrein | Pinctada fucata | BAA11940.1 |
| PmaNA | Nacrein-like | Pinctada maxima | BAF42330.1 |
| PmarNA | Nacrein | Pinctada margaritifera | AEC03970.1 |
| CgiNAPre | Nacrein precursor | Crassostrea gigas | NP_001292235.1 |
| CgiNAF | Nacrein isoform F | Crassostrea gigas | EKC37961.1 |
| CgiNAX4 | Nacrein isoform X4 | Crassostrea gigas | XP_011426380.1 |
| CgiNAP2X1 | Nacrein-like isoform P2X1 | Crassostrea gigas | XP_011428406.1 |
| CgiNA | Nacrein-like | Crassostrea gigas | XP_011428408.1 |
| CgiNA_Mit | Mitochondrial nacrein | Crassostrea gigas | EKC36446.1 |
| TmaNA | Nacrein | Turbo marmoratus | BAB91157.1 |
| HtuCA | α-CA | Haliotis tuberculata | AEL22200.1 |
| HgiCA | α-CA | Haliotis gigantea | BAH58350.1 |
| SsalCA | α-CA | Streptococcus salivarius] | WP_002888224.1 |
| NgonCA | α-CA | Neisseria gonorrhoeae | WP_003688976.1 |
| SspCA | α-CA | Sulfurihydrogenibium yellowstonense | WP_012459296.1 |
| HpylCA | α-CA | Helicobacter pylori | WP_010882609.1 |
| CruCA4 | α-CA isoform 4 | Corallium rubrum | KU557746.1 |
| SpiCA2 | α-CA isoform 2 | Stylophora pistillata | ACE95141.1 |
| HumCAI | α-CA isoform 1 | Homo sapiens | NP_001729.1 |
| HumCAII | α-CA isoform 2 | Homo sapiens | AAH11949.1 |
| PokCA | α-CA | Phreagena okutanii | BAO01209.1 |
| MgaCA | α-CA | Mytilus galloprovincialis | ALF62133.1 |
| CgiCA2 | α-CA isoform 2 | Crassostrea gigas | EKC31880.1 |
| CgiCA2X1 | α-CA isoform 2X1 | Crassostrea gigas | XP_011447892.1 |
| CgiCA2X3 | α-CA isoform 2X3 | Crassostrea gigas | XP_011447895.1 |
| SpCA1 | α-CA isoform 1 | Stylophora pistillata | ACA53457.1 |
| TgiCA | α-CA | Tridacna gigas | AAX16122.1 |
| CgiCA12 | α-CA isoform 12 | Crassostrea gigas | EKC34762.1 |
| ObiCA9X | α-CA isoform 9X | Octopus bimaculoides | XP_014786584.1 |
| HblCA | α-CA | Heterololigo bleekeri | BAL42244.1 |
| Enzyme | Organism | Class | kcat 1 (s−1) | kcat/KM1 (M−1 × s−1) | KIs (acetazolamide) 1 (nM) |
|---|---|---|---|---|---|
| HumCAI | Homo sapiens | α | 2.0 × 105 | 5.0 × 107 | 250 |
| HumCAII | Homo sapiens | α | 1.4 × 106 | 1.5 × 108 | 12 |
| CgiNAP2X1 | Crassotrea gigas | Nacrein-like | 1.0 × 106 | 1.2 × 108 | 495 |
| SpiCA1 | Stylophora pistillata | α | 3.1 × 105 | 4.6 × 107 | 16 |
| SpiCA2 | Stylophora pistillata | α | 5.6 × 105 | 8.3 × 107 | 74 |
| CruCA4 | Corallium rubrum | α | 2.4 × 105 | 5.2 × 107 | 450 |
| MgaCA | Mytilus galloprovincialis | α | 4.1 × 105 | 3.6 × 107 | 380 |
| rec-MgaCA | Mytilus galloprovincialis | α | 4.2 × 105 | 3.5 × 107 | 361 |
| Inhibitor 1 | KIs [mM] 2 | ||
|---|---|---|---|
| hCA I | hCA II | CgiNAP2X1 | |
| F− | >100 | >100 | 12.1 |
| Cl− | 6 | >100 | 10.8 |
| Br− | 4 | 63 | 16.6 |
| I− | 0.3 | 26 | 8.86 |
| CNO− | 0.0007 | 0.03 | 3.11 |
| SCN− | 0.2 | 1.60 | 5.88 |
| CN− | 0.0005 | 0.02 | >100 |
| N3− | 0.0012 | 1.51 | >100 |
| HCO3− | 12 | 85 | 1.98 |
| CO32− | 15 | 73 | 6.62 |
| NO3− | 7 | 35 | 10.3 |
| NO2− | 8.4 | 63 | 7.14 |
| HS− | 0.0006 | 0.04 | 8.19 |
| HSO3− | 18 | 89 | >100 |
| SnO32− | 0.57 | 0.83 | 3.47 |
| SeO42− | >100 | >100 | 8.74 |
| TeO42− | 0.66 | 0.92 | >100 |
| OsO52− | 0.92 | 0.95 | >100 |
| P2O74− | 25.8 | 48.5 | 9.47 |
| V2O74− | 0.54 | 0.57 | >100 |
| B4O72− | 0.64 | 0.95 | 8.85 |
| ReO4− | 0.11 | 0.75 | >100 |
| RuO4− | 0.101 | 0.69 | >100 |
| S2O82− | 0.107 | 0.084 | 20.7 |
| SeCN− | 0.085 | 0.086 | 8.21 |
| CS32− | 0.0087 | 0.0088 | 31.7 |
| Et2NCS2− | 0.00079 | 0.0031 | 0.76 |
| CF3SO3− | >100 | >100 | 8.68 |
| PF6− | >100 | >100 | >100 |
| SO42− | 63 | >100 | 14.5 |
| ClO4− | >100 | >100 | >100 |
| BF4− | >100 | >100 | >100 |
| FSO3− | 0.79 | 0.46 | >100 |
| NH(SO3)22− | 0.31 | 0.76 | >100 |
| H2NSO2NH2 | 0.31 | 1.13 | 0.076 |
| H2NSO3H | 0.021 | 0.39 | 0.073 |
| Ph-B(OH)2 | 58.6 | 23.1 | 0.072 |
| Ph-AsO3H2 | 31.7 | 49.2 | 0.087 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Perfetto, R.; Del Prete, S.; Vullo, D.; Sansone, G.; Barone, C.M.A.; Rossi, M.; Supuran, C.T.; Capasso, C. Sequence Analysis, Kinetic Constants, and Anion Inhibition Profile of the Nacrein-Like Protein (CgiNAP2X1) from the Pacific Oyster Magallana gigas (Ex-Crassostrea gigas). Mar. Drugs 2017, 15, 270. https://doi.org/10.3390/md15090270
Perfetto R, Del Prete S, Vullo D, Sansone G, Barone CMA, Rossi M, Supuran CT, Capasso C. Sequence Analysis, Kinetic Constants, and Anion Inhibition Profile of the Nacrein-Like Protein (CgiNAP2X1) from the Pacific Oyster Magallana gigas (Ex-Crassostrea gigas). Marine Drugs. 2017; 15(9):270. https://doi.org/10.3390/md15090270
Chicago/Turabian StylePerfetto, Rosa, Sonia Del Prete, Daniela Vullo, Giovanni Sansone, Carmela M. A. Barone, Mosè Rossi, Claudiu T. Supuran, and Clemente Capasso. 2017. "Sequence Analysis, Kinetic Constants, and Anion Inhibition Profile of the Nacrein-Like Protein (CgiNAP2X1) from the Pacific Oyster Magallana gigas (Ex-Crassostrea gigas)" Marine Drugs 15, no. 9: 270. https://doi.org/10.3390/md15090270
APA StylePerfetto, R., Del Prete, S., Vullo, D., Sansone, G., Barone, C. M. A., Rossi, M., Supuran, C. T., & Capasso, C. (2017). Sequence Analysis, Kinetic Constants, and Anion Inhibition Profile of the Nacrein-Like Protein (CgiNAP2X1) from the Pacific Oyster Magallana gigas (Ex-Crassostrea gigas). Marine Drugs, 15(9), 270. https://doi.org/10.3390/md15090270