Effect of Omega-3 Fatty Acid on STAMP2 Expression in the Heart and Kidney of 5/6 Nephrectomy Rat Model



Abstract
:1. Introduction
2. Results
2.1. Laboratory Data
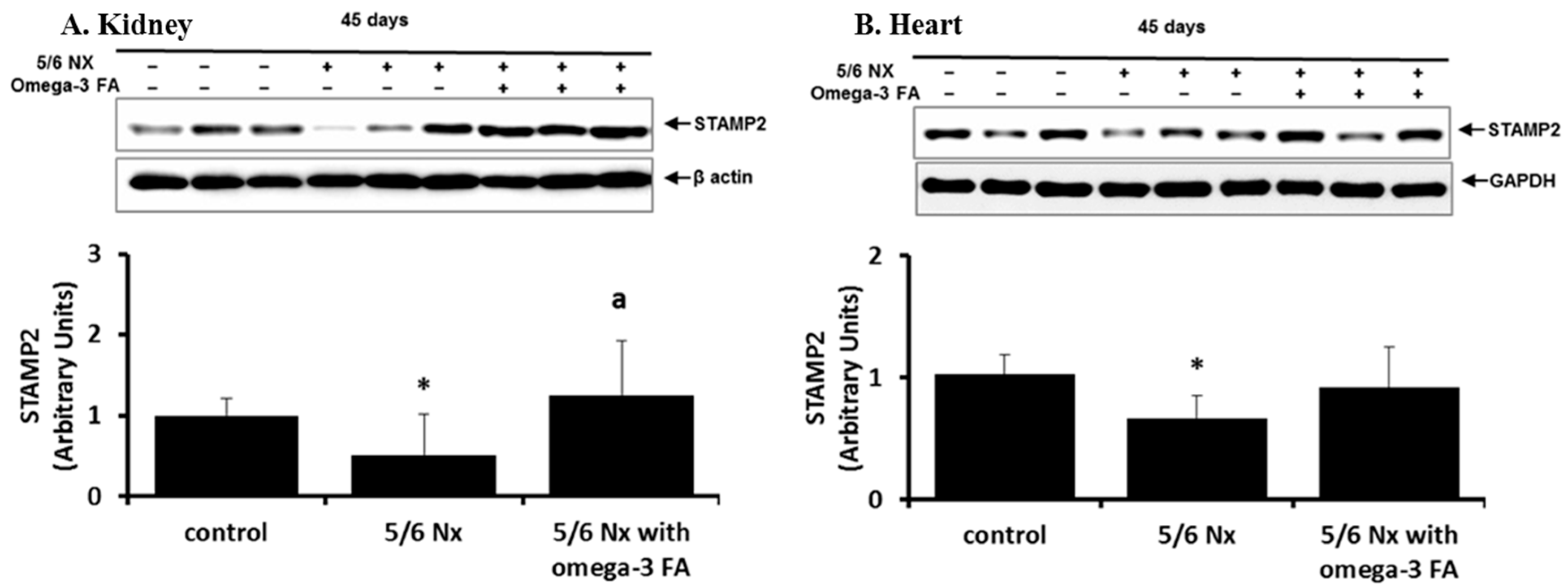
2.2. STAMP2 Data
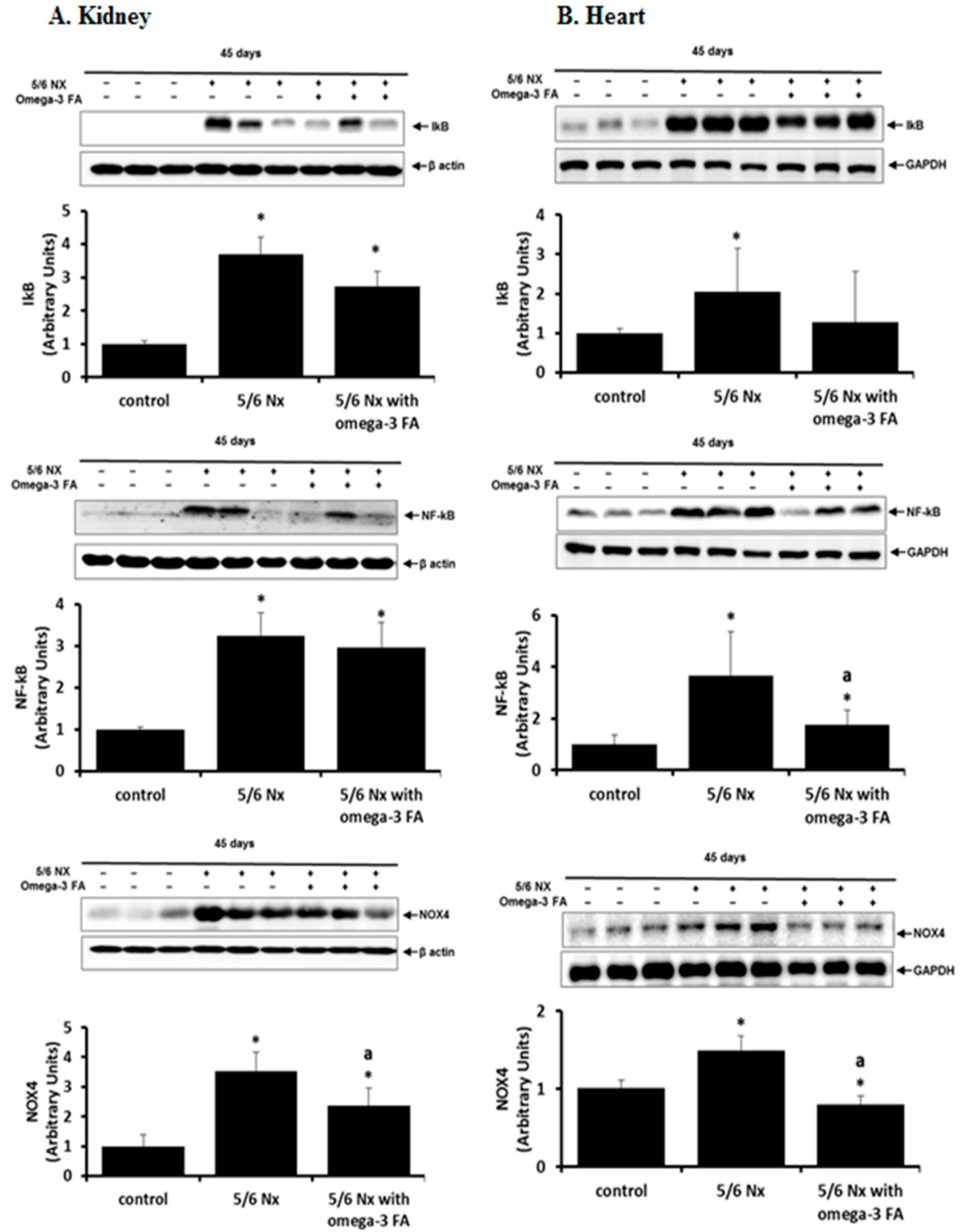
2.3. IκB, NF-κB, and NOX4 Data
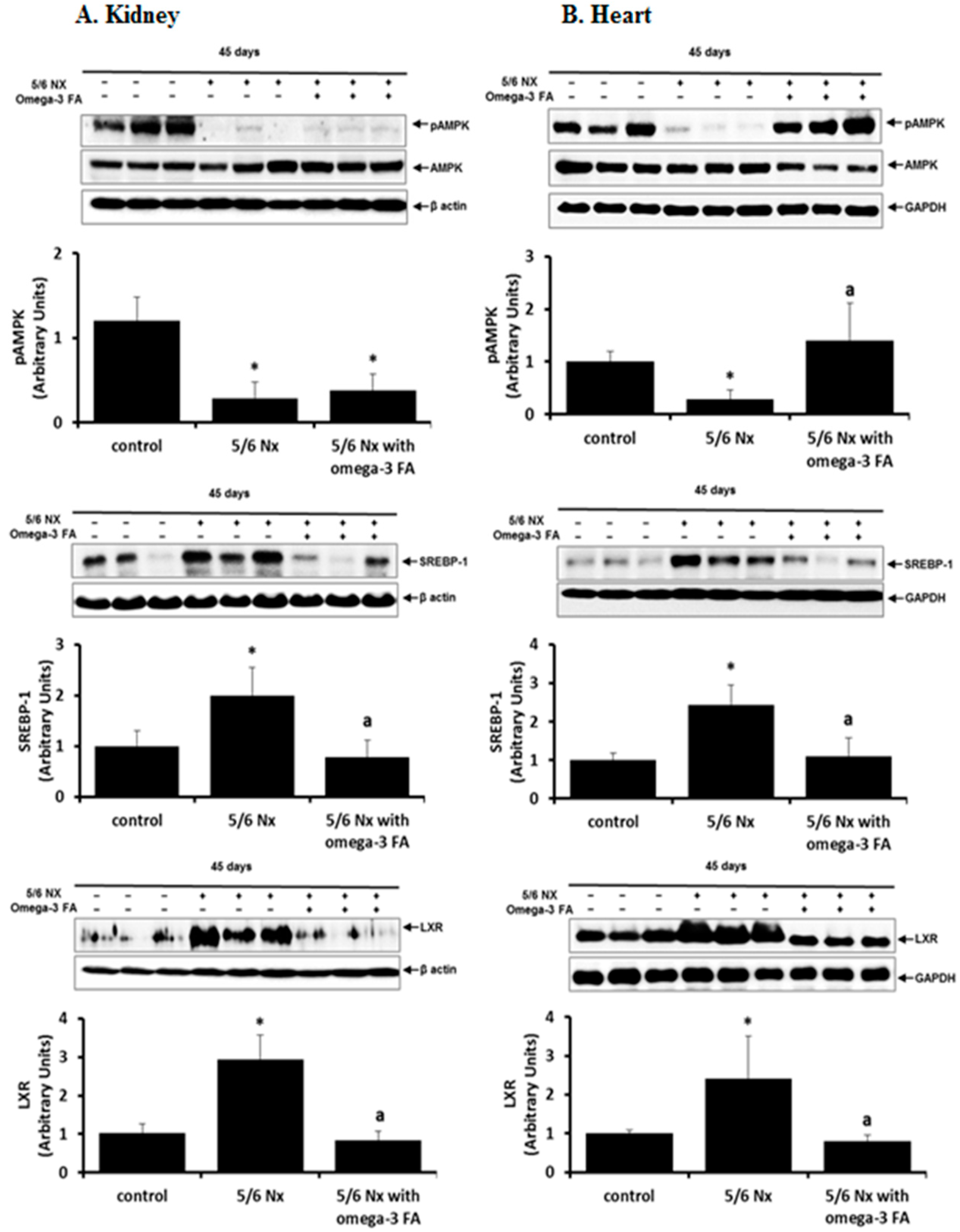
2.4. AMPK, SREBP-1, and LXR Data
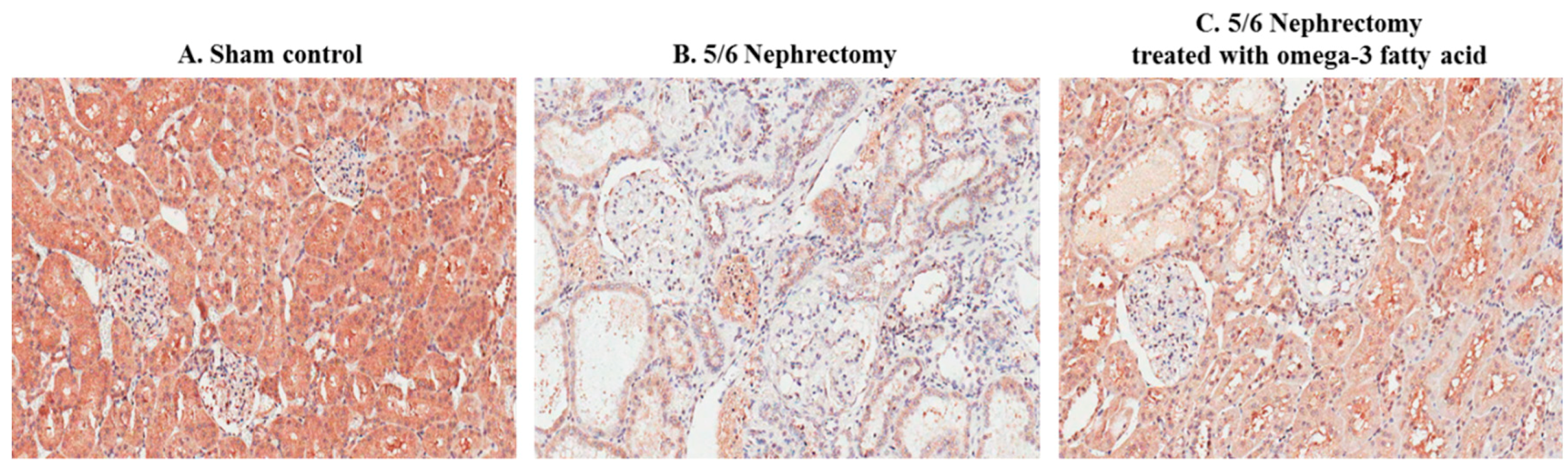
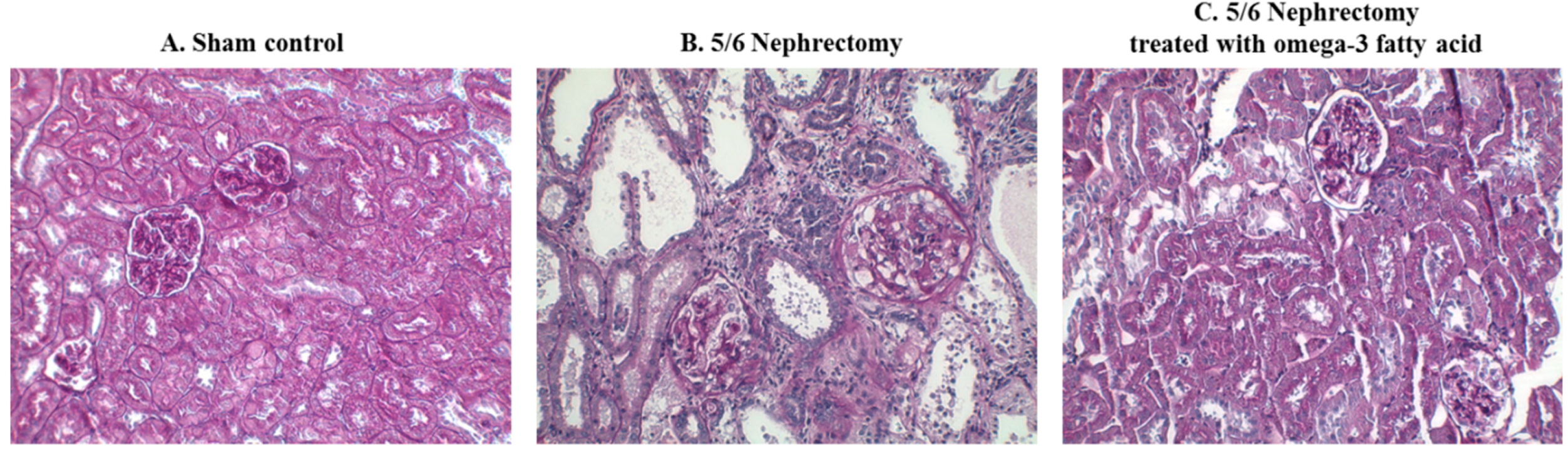
2.5. Renal Pathology
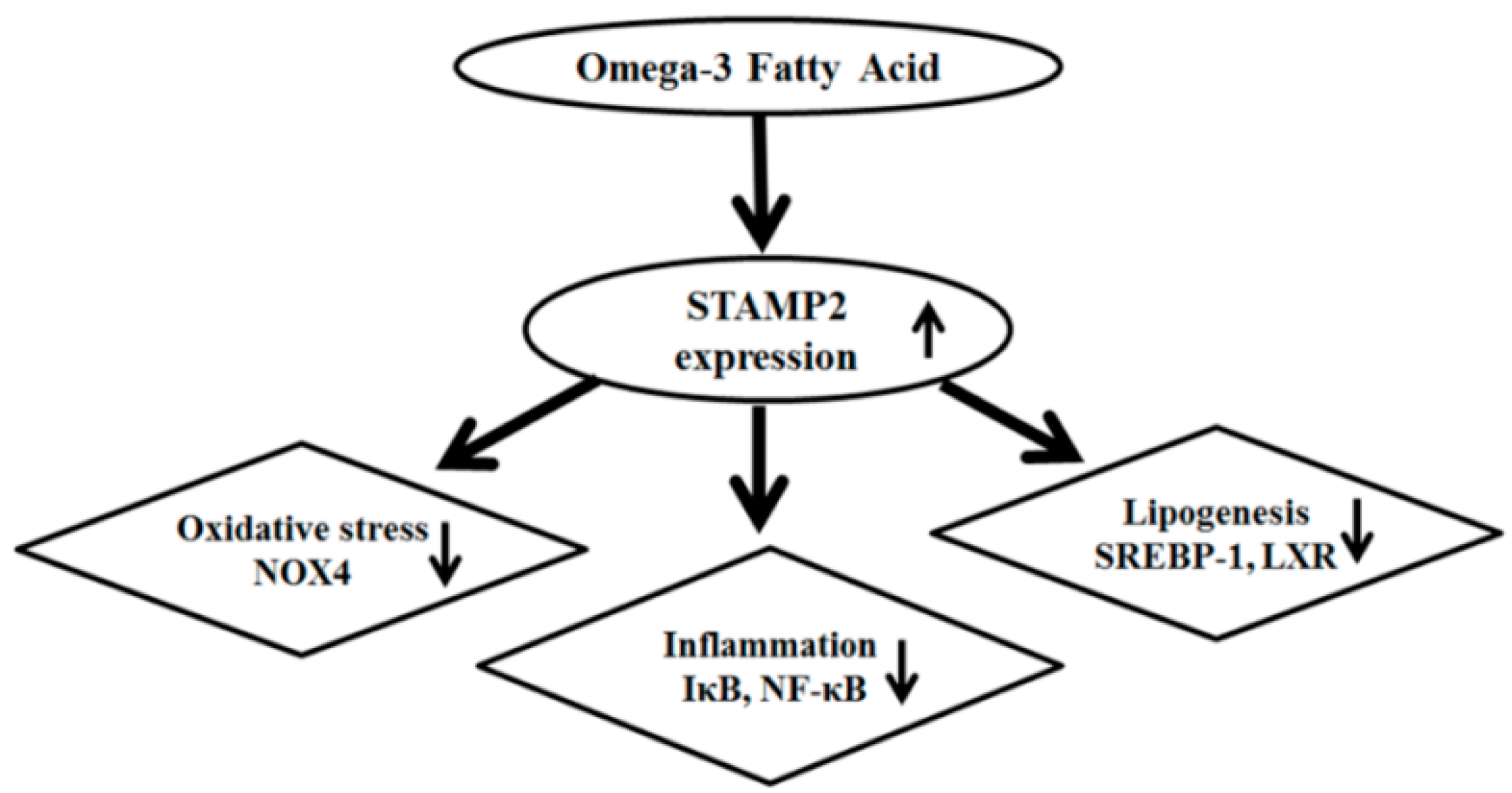
3. Discussion
4. Materials and Methods
4.1. Animals and Experimental Design
4.2. Histopathologic Evaluation
4.3. Immunohistochemical Staining
4.4. Western Blotting
4.5. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Foley, R.N.; Parfrey, P.S.; Sarnak, M.J. Clinical epidemiology of cardiovascular disease in chronic renal disease. Am. J. Kidney Dis. 1998, 32, S112–S119. [Google Scholar] [CrossRef] [PubMed]
- Stenvinkel, P.; Heimburger, O.; Paultre, F.; Diczfalusy, U.; Wang, T.; Berglund, L.; Jogestrand, T. Strong association between malnutrition, inflammation, and atherosclerosis in chronic renal failure. Kidney Int. 1999, 55, 1899–1911. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, C.G.; Korkmaz, K.S.; Kurys, P.; Elbi, C.; Wang, L.; Klokk, T.I.; Hammarstrom, C.; Troen, G.; Svindland, A.; Hager, G.L.; et al. Molecular cloning and characterization of stamp2, an androgen-regulated six transmembrane protein that is overexpressed in prostate cancer. Oncogene 2005, 24, 4934–4945. [Google Scholar] [CrossRef] [PubMed]
- Ohgami, R.S.; Campagna, D.R.; McDonald, A.; Fleming, M.D. The steap proteins are metalloreductases. Blood 2006, 108, 1388–1394. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.M.; Chi, X.; Wang, B.; Zhang, M.; Ni, Y.H.; Chen, R.H.; Li, X.N.; Guo, X.R. Downregulation of steap4, a highly-expressed tnf-alpha-inducible gene in adipose tissue, is associated with obesity in humans. Acta Pharmacol. Sin. 2008, 29, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Moldes, M.; Lasnier, F.; Gauthereau, X.; Klein, C.; Pairault, J.; Feve, B.; Chambaut-Guerin, A.M. Tumor necrosis factor-alpha-induced adipose-related protein (tiarp), a cell-surface protein that is highly induced by tumor necrosis factor-alpha and adipose conversion. J. Biol. Chem. 2001, 276, 33938–33946. [Google Scholar] [CrossRef] [PubMed]
- Wellen, K.E.; Fucho, R.; Gregor, M.F.; Furuhashi, M.; Morgan, C.; Lindstad, T.; Vaillancourt, E.; Gorgun, C.Z.; Saatcioglu, F.; Hotamisligil, G.S. Coordinated regulation of nutrient and inflammatory responses by stamp2 is essential for metabolic homeostasis. Cell 2007, 129, 537–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.Y.; Park, S.Y.; Lee, M.H.; Rho, J.H.; Oh, Y.J.; Jung, H.U.; Yoo, S.H.; Jeong, N.Y.; Lee, H.J.; Suh, S.; et al. Hepatic stamp2 alleviates high fat diet-induced hepatic steatosis and insulin resistance. J. Hepatol. 2015, 63, 477–485. [Google Scholar] [CrossRef] [PubMed]
- Waki, H.; Tontonoz, P. Stamping out inflammation. Cell 2007, 129, 451–452. [Google Scholar] [CrossRef] [PubMed]
- Han, L.; Tang, M.X.; Ti, Y.; Wang, Z.H.; Wang, J.; Ding, W.Y.; Wang, H.; Zhang, Y.; Zhang, W.; Zhong, M. Overexpressing STAMP2 improves insulin resistance in diabetic ApoE−/−/LDLR−/− mice via macrophage polarization shift in adipose tissues. PLoS ONE 2013, 8, e78903. [Google Scholar] [CrossRef]
- Ruan, X.Z.; Moorhead, J.F.; Fernando, R.; Wheeler, D.C.; Powis, S.H.; Varghese, Z. Regulation of lipoprotein trafficking in the kidney: Role of inflammatory mediators and transcription factors. Biochem. Soc. Trans. 2004, 32, 88–91. [Google Scholar] [CrossRef] [PubMed]
- Hardie, D.G. The AMP-activated protein kinase pathway—New players upstream and downstream. J. Cell Sci. 2004, 117, 5479–5487. [Google Scholar] [CrossRef] [PubMed]
- Finucane, O.M.; Lyons, C.L.; Murphy, A.M.; Reynolds, C.M.; Klinger, R.; Healy, N.P.; Cooke, A.A.; Coll, R.C.; McAllan, L.; Nilaweera, K.N.; et al. Monounsaturated fatty acid-enriched high-fat diets impede adipose NLRP3 inflammasome-mediated IL-1β secretion and insulin resistance despite obesity. Diabetes 2015, 64, 2116–2128. [Google Scholar] [CrossRef] [PubMed]
- An, W.S.; Kim, H.J.; Cho, K.H.; Vaziri, N.D. Omega-3 fatty acid supplementation attenuates oxidative stress, inflammation, and tubulointerstitial fibrosis in the remnant kidney. Am. J. Physiol. Renal. Physiol. 2009, 297, F895–F903. [Google Scholar] [CrossRef] [PubMed]
- Svensson, M.; Schmidt, E.B.; Jorgensen, K.A.; Christensen, J.H. N-3 fatty acids as secondary prevention against cardiovascular events in patients who undergo chronic hemodialysis: A randomized, placebo-controlled intervention trial. Clin. J. Am. Soc. Nephrol. 2006, 1, 780–786. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; An, W.S. Cardioprotective effects of omega-3 pufas in chronic kidney disease. BioMed Res. Int. 2013, 2013, 712949. [Google Scholar] [PubMed]
- Eslick, G.D.; Howe, P.R.; Smith, C.; Priest, R.; Bensoussan, A. Benefits of fish oil supplementation in hyperlipidemia: A systematic review and meta-analysis. Int. J. Cardiol. 2009, 136, 4–16. [Google Scholar] [CrossRef] [PubMed]
- Serhan, C.N.; Chiang, N.; Van Dyke, T.E. Resolving inflammation: Dual anti-inflammatory and pro-resolution lipid mediators. Nat. Rev. Immunol. 2008, 8, 349–361. [Google Scholar] [CrossRef] [PubMed]
- James, M.; Proudman, S.; Cleland, L. Fish oil and rheumatoid arthritis: Past, present and future. Proc. Nutr. Soc. 2010, 69, 316–323. [Google Scholar] [CrossRef] [PubMed]
- An, W.S.; Kim, S.E.; Kim, K.H.; Lee, S.; Park, Y.; Kim, H.J.; Vaziri, N.D. Comparison of fatty acid contents of erythrocyte membrane in hemodialysis and peritoneal dialysis patients. J. Ren. Nutr. 2009, 19, 267–274. [Google Scholar] [CrossRef] [PubMed]
- An, W.S.; Lee, S.M.; Son, Y.K.; Kim, S.E.; Kim, K.H.; Han, J.Y.; Bae, H.R.; Rha, S.H.; Park, Y. Omega-3 fatty acid supplementation increases 1,25-dihydroxyvitamin d and fetuin-a levels in dialysis patients. Nutr. Res. 2012, 32, 495–502. [Google Scholar] [CrossRef] [PubMed]
- An, W.S.; Lee, S.M.; Son, Y.K.; Kim, S.E.; Kim, K.H.; Han, J.Y.; Bae, H.R.; Park, Y. Effect of omega-3 fatty acids on the modification of erythrocyte membrane fatty acid content including oleic acid in peritoneal dialysis patients. Prostag. Leukotr. Ess. 2012, 86, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.N.; Chen, X.; Li, R.; Gao, B.; Mohammed-Ali, Z.; Lu, C.; Yum, V.; Dickhout, J.G.; Krepinsky, J.C. Srebp-1 mediates angiotensin ii-induced tgf-beta1 upregulation and glomerular fibrosis. J. Am. Soc. Nephrol. 2015, 26, 1839–1854. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Wang, T.; Uttarwar, L.; vanKrieken, R.; Li, R.; Chen, X.; Gao, B.; Ghayur, A.; Margetts, P.; Krepinsky, J.C. Srebp-1 is a novel mediator of tgfbeta1 signaling in mesangial cells. J. Mol. Cell Biol. 2014, 6, 516–530. [Google Scholar] [CrossRef] [PubMed]
- Chu, K.; Miyazaki, M.; Man, W.C.; Ntambi, J.M. Stearoyl-coenzyme a desaturase 1 deficiency protects against hypertriglyceridemia and increases plasma high-density lipoprotein cholesterol induced by liver x receptor activation. Mol. Cell Biol. 2006, 26, 6786–6798. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, M.; Dobrzyn, A.; Man, W.C.; Chu, K.; Sampath, H.; Kim, H.J.; Ntambi, J.M. Stearoyl-coa desaturase 1 gene expression is necessary for fructose-mediated induction of lipogenic gene expression by sterol regulatory element-binding protein-1c-dependent and -independent mechanisms. J. Biol. Chem. 2004, 279, 25164–25171. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Navarrete, J.M.; Ortega, F.; Serrano, M.; Perez-Perez, R.; Sabater, M.; Ricart, W.; Tinahones, F.; Peral, B.; Fernandez-Real, J.M. Decreased stamp2 expression in association with visceral adipose tissue dysfunction. J. Clin. Endocrinol. Metab. 2011, 96, E1816–E1825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Towler, M.C.; Hardie, D.G. Amp-activated protein kinase in metabolic control and insulin signaling. Circ. Res. 2007, 100, 328–341. [Google Scholar] [CrossRef] [PubMed]
- Hardie, D.G. Amp-activated/snf1 protein kinases: Conserved guardians of cellular energy. Nat. Rev. Mol. Cell Biol. 2007, 8, 774–785. [Google Scholar] [CrossRef] [PubMed]
- Sharma, K.; Ramachandrarao, S.; Qiu, G.; Usui, H.K.; Zhu, Y.; Dunn, S.R.; Ouedraogo, R.; Hough, K.; McCue, P.; Chan, L.; et al. Adiponectin regulates albuminuria and podocyte function in mice. J. Clin. Investig. 2008, 118, 1645–1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kume, S.; Uzu, T.; Araki, S.; Sugimoto, T.; Isshiki, K.; Chin-Kanasaki, M.; Sakaguchi, M.; Kubota, N.; Terauchi, Y.; Kadowaki, T.; et al. Role of altered renal lipid metabolism in the development of renal injury induced by a high-fat diet. J. Am. Soc. Nephrol. 2007, 18, 2715–2723. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, M.; Liang, B.; Xu, J.; Xie, Z.; Liu, C.; Viollet, B.; Yan, D.; Zou, M.H. Ampkalpha2 deletion causes aberrant expression and activation of nad(p)h oxidase and consequent endothelial dysfunction in vivo: Role of 26s proteasomes. Circ. Res. 2010, 106, 1117–1128. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.C.; Yun, S.R.; Lee, S.W.; Han, S.W.; Kim, W.; Park, J.; Kim, Y.K. Current characteristics of dialysis therapy in korea: 2016 registry data focusing on diabetic patients. Kidney Res. Clin. Pract. 2018, 37, 20–29. [Google Scholar] [CrossRef] [PubMed]
- Leaf, A.; Kang, J.X.; Xiao, Y.F.; Billman, G.E. Clinical prevention of sudden cardiac death by n-3 polyunsaturated fatty acids and mechanism of prevention of arrhythmias by n-3 fish oils. Circulation 2003, 107, 2646–2652. [Google Scholar] [CrossRef] [PubMed]
- Burr, M.L.; Fehily, A.M.; Gilbert, J.F.; Rogers, S.; Holliday, R.M.; Sweetnam, P.M.; Elwood, P.C.; Deadman, N.M. Effects of changes in fat, fish, and fibre intakes on death and myocardial reinfarction: Diet and reinfarction trial (dart). Lancet 1989, 2, 757–761. [Google Scholar] [CrossRef]
- Dietary supplementation with n-3 polyunsaturated fatty acids and vitamin E after myocardial infarction: Results of the gissi-prevenzione trial. Gruppo italiano per lo studio della sopravvivenza nell′infarto miocardico. Lancet 1999, 354, 447–455. [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control (n = 6) | 5/6 Nephrectomy (n = 6) | 5/6 Nephrectomy with Omega-3 Fatty Acid (n = 6) | p Value | |
|---|---|---|---|---|
| Final Body Weight (g) | 463.1 ± 8.6 | 397.7 ± 21.1 * | 429.7 ± 21.3 *,a | 0.002 |
| Body Weight Gain (g) | 91.0 ± 3.0 | 58.5 ± 22.5 * | 82.7 ± 18.6 * | 0.006 |
| Blood Urea Nitrogen (mg/dL) | 17.7 ± 1.5 | 77.7 ± 28.4 * | 63.9 ± 17.0 * | 0.007 |
| Creatinine (mg/dL) | 0.4 ± 0.0 | 1.3 ± 0.6 * | 1.0 ± 0.3 * | 0.008 |
| Calcium (mg/dL) | 6.8 ± 0.3 | 6.9 ± 0.7 | 7.2 ± 1.0 | 0.663 |
| Phosphorus (mg/dL) | 8.4 ± 0.4 | 9.7 ± 4.0 | 8.2 ± 0.6 | 0.327 |
| Total Cholesterol (mg/dL) | 61.9 ± 7.3 | 134.2 ± 32.2 * | 107.2 ± 37.9 * | 0.005 |
| Triglyceride (mg/dL) | 134.0 ± 27.2 | 266.5 ± 87.5 * | 216.8 ± 157.4 | 0.100 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.W.; Lee, S.M.; Lee, M.H.; Son, Y.K.; Kim, S.E.; An, W.S. Effect of Omega-3 Fatty Acid on STAMP2 Expression in the Heart and Kidney of 5/6 Nephrectomy Rat Model. Mar. Drugs 2018, 16, 398. https://doi.org/10.3390/md16110398
Lee HW, Lee SM, Lee MH, Son YK, Kim SE, An WS. Effect of Omega-3 Fatty Acid on STAMP2 Expression in the Heart and Kidney of 5/6 Nephrectomy Rat Model. Marine Drugs. 2018; 16(11):398. https://doi.org/10.3390/md16110398
Chicago/Turabian StyleLee, Hye Won, Su Mi Lee, Mi Hwa Lee, Young Ki Son, Seong Eun Kim, and Won Suk An. 2018. "Effect of Omega-3 Fatty Acid on STAMP2 Expression in the Heart and Kidney of 5/6 Nephrectomy Rat Model" Marine Drugs 16, no. 11: 398. https://doi.org/10.3390/md16110398
APA StyleLee, H. W., Lee, S. M., Lee, M. H., Son, Y. K., Kim, S. E., & An, W. S. (2018). Effect of Omega-3 Fatty Acid on STAMP2 Expression in the Heart and Kidney of 5/6 Nephrectomy Rat Model. Marine Drugs, 16(11), 398. https://doi.org/10.3390/md16110398