Identification and Molecular Docking Study of a Novel Angiotensin-I Converting Enzyme Inhibitory Peptide Derived from Enzymatic Hydrolysates of Cyclina sinensis

 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
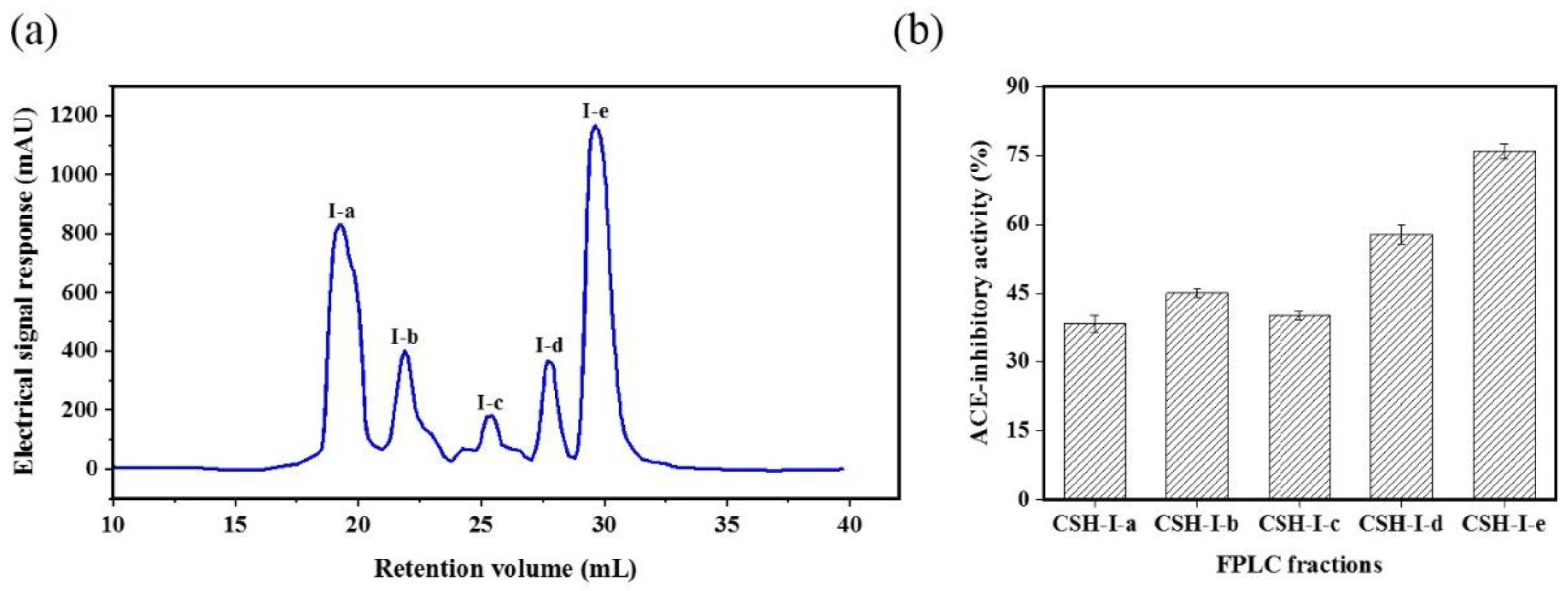
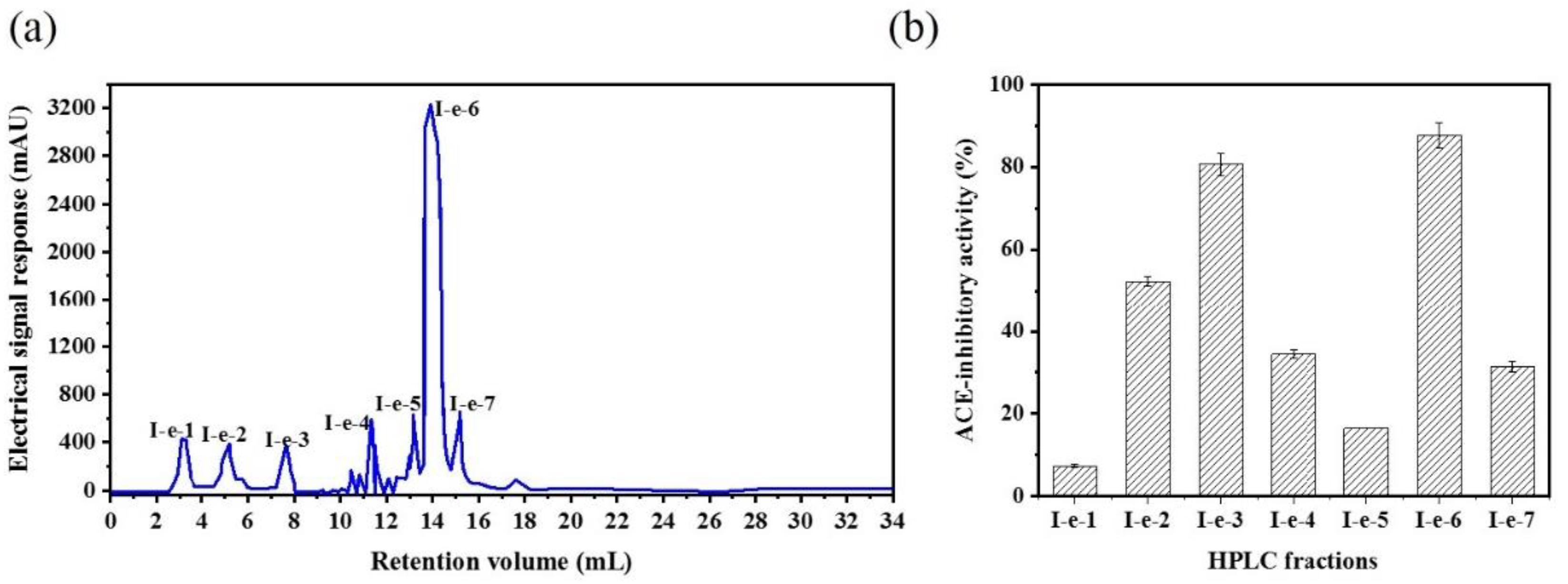
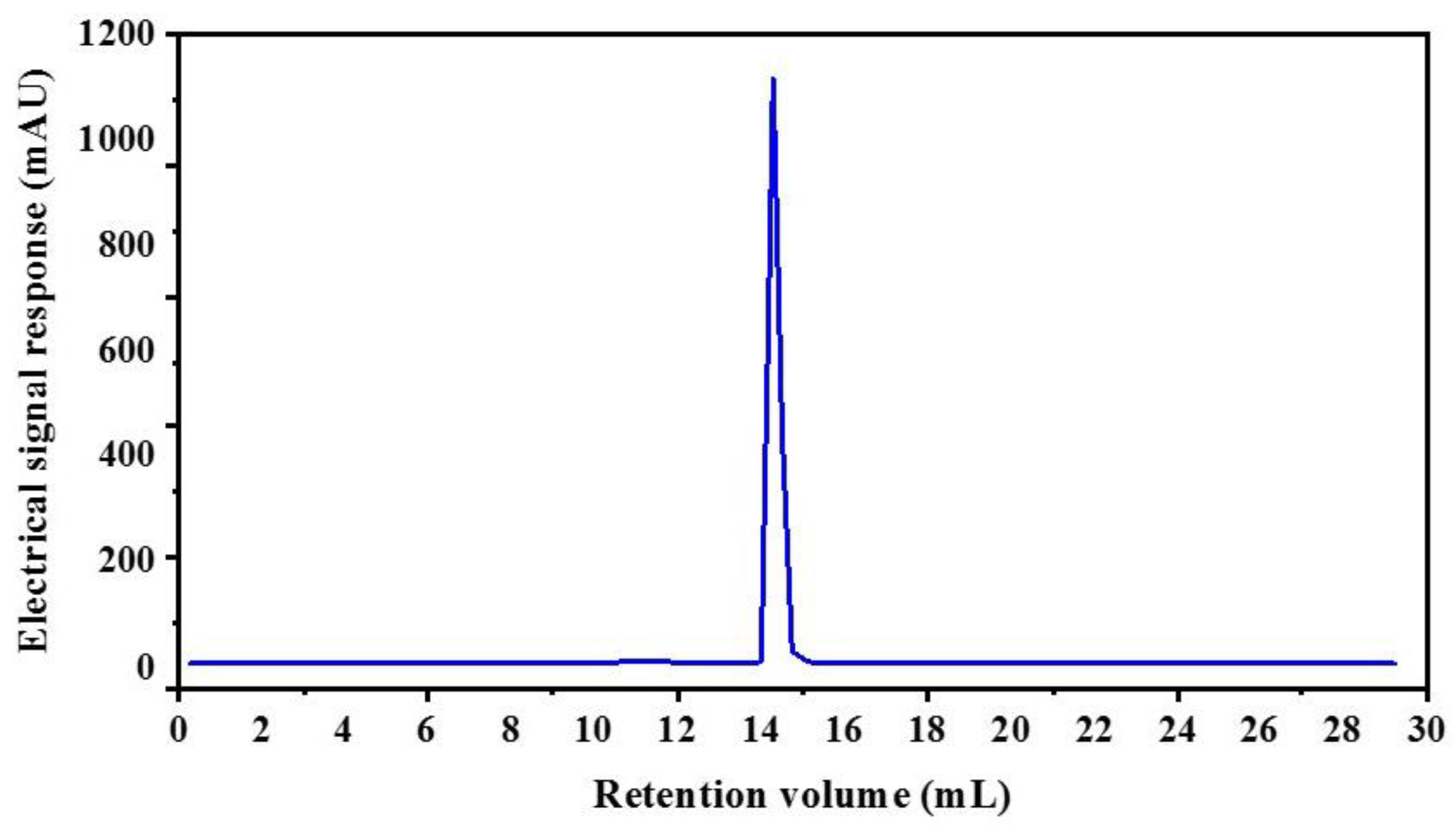
2.1. Isolation and Purification of an Angiotensin-I Converting Enzyme (ACE) Inhibitory Peptide
2.2. Amino Acid Sequence and IC50 Value of the Purified Peptide
2.3. Analysis of the ACE Inhibitory Characteristics of Peptide WPMGF
2.4. Stability of Peptide WPMGF
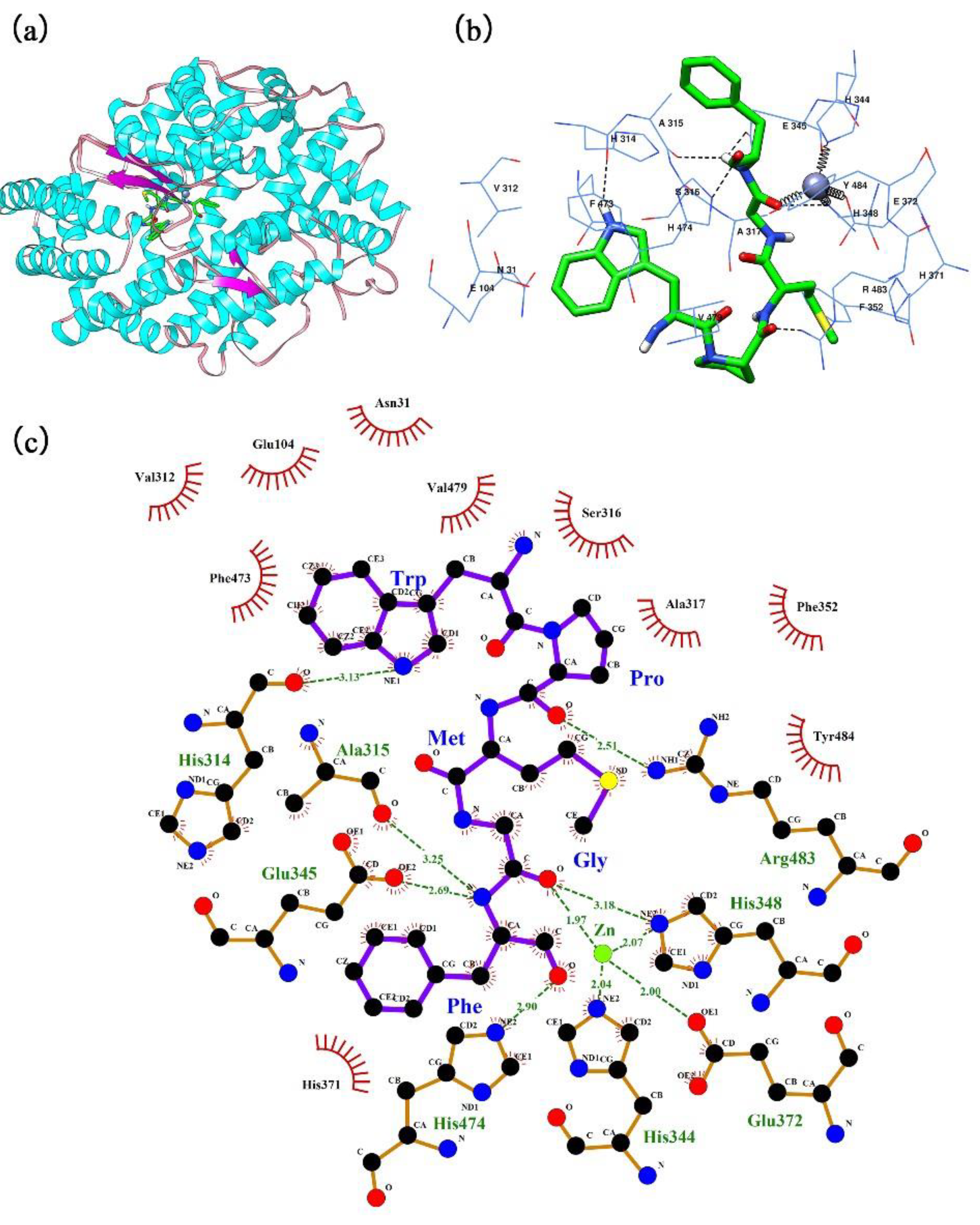
2.5. Insights into the Molecular Docking Mechanism
3. Materials and Methods
3.1. Materials and Chemicals
3.2. Production of Cyclina Sinensis (CSH)
3.3. Determination of ACE Inhibitory Activity
3.4. Isolation and Purification of ACE-Inhibitory Peptides
3.4.1. Ultrafiltration (UF)
3.4.2. Fast protein liquid chromatography (FPLC)
3.4.3. Reversed-phase high-performance liquid chromatography (RP-HPLC)
3.5. N-Terminal Sequencing of the ACE Inhibitory Peptide
3.6. Synthesis of ACE Inhibitory Peptide
3.7. Determination of the ACE-Inhibitory Characteristics of the Purified Peptide
3.8. Stability Studies
3.8.1. Thermal and pH Stability of the ACE-Inhibitory Peptide
3.8.2. Effects of Simulated Gastroenteric Environments on the ACE-Inhibitory Activity of the Peptide
3.9. Molecular Docking of the Peptide in the ACE Binding Site
3.10. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- World Health Organization. A Global Brief on Hypertension: Silent Killer, Global Public Health Crisis: World Health Day 2013. Available online: http://www.who.int/cardiovascular_diseases/publications/global_brief_hypertension/en (accessed on 26 October 2018).
- Lim, S.S.; Vos, T.; Flaxman, A.D.; Danaei, G.; Shibuya, K.; Adair-Rohani, H.; AlMazroa, M.A.; Amann, M.; Anderson, H.R.; Andrew, K.G.; et al. A comparative risk assessment of burden of disease and injury attributable to 67 risk factors and risk factor clusters in 21 regions, 1990–2010: A systematic analysis for the Global Burden of Disease Study 2010. Lancet 2012, 380, 2224–2260. [Google Scholar] [CrossRef]
- Liu, L.S.; Writing Group of Chinese Guidelines for the Management of Hypertension. [2010 Chinese guidelines for the management of hypertension]. Zhonghua Xin Xue Guan Bing Za Zhi 2011, 39, 579–615. [Google Scholar] [PubMed]
- Hooper, N.M. Angiotensin I-Converting Enzyme (ACE). In xPharm: The Comprehensive Pharmacology Reference; Enna, S.J., Bylund, D.B., Eds.; Elsevier: New York, NY, USA, 2007; pp. 1–7. [Google Scholar]
- Heidari, F.; Vasudevan, R.; Mohd Ali, S.Z.; Ismail, P.; Arkani, M. RAS Genetic Variants in Interaction with ACE Inhibitors Drugs Influences Essential Hypertension Control. Arch. Med. Res. 2017, 48, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Regoli, D.; Gobeil, F. Critical insights into the beneficial and protective actions of the kallikrein–kinin system. Vasc. Pharmacol. 2015, 64, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Roytrakul, S.; Sutheerawattananonda, M. Production and purification of glucosamine and angiotensin-I converting enzyme (ACE) inhibitory peptides from mushroom hydrolysates. J. Funct. Foods. 2017, 36, 72–83. [Google Scholar] [CrossRef]
- Patchett, A.A.; Harris, E.; Tristram, E.W.; Wyvratt, M.J.; Wu, M.T.; Taub, D.; Peterson, E.R.; Ikeler, T.J.; ten Broeke, J.; Payne, L.G.; et al. A new class of angiotensin-converting enzyme inhibitors. Nature 1980, 288, 280–283. [Google Scholar] [CrossRef] [PubMed]
- Bylund, D.B. Captopril: Reference Module in Biomedical Sciences. Elsevier 2017, 1–7. [Google Scholar]
- Aronson, J.K. Enalapril: Meyler′s Side Effects of Drugs. Elsevier 2016, 16, 41–46. [Google Scholar]
- Kim, S.-K.; Wijesekara, I. Development and biological activities of marine-derived bioactive peptides: A review. J. Funct. Foods 2010, 2, 1–9. [Google Scholar] [CrossRef]
- Chen, J.; Liu, Y.; Wang, G.; Sun, S.; Liu, R.; Hong, B.; Gao, R.; Bai, K. Processing Optimization and Characterization of Angiotensin-Ι-Converting Enzyme Inhibitory Peptides from Lizardfish (Synodus macrops) Scale Gelatin. Mar. Drugs 2018, 16, 228. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Chen, Y.; Zhao, W.; Li, J.; Liu, J.; Chen, F. Identification and molecular docking study of novel angiotensin-converting enzyme inhibitory peptides from Salmo salar using in silico methods. J. Sci. Food Agric. 2018, 98, 3907–3914. [Google Scholar] [CrossRef] [PubMed]
- Krichen, F.; Sila, A.; Caron, J.; Kobbi, S.; Nedjar, N.; Miled, N.; Blecker, C.; Besbes, S.; Bougatef, A. Identification and molecular docking of novel ACE inhibitory peptides from protein hydrolysates of shrimp waste. Eng. Life Sci. 2018, 18, 682–691. [Google Scholar] [CrossRef]
- Yu, Y.; Fan, F.; Wu, D.; Yu, C.; Wang, Z.; Du, M. Antioxidant and ACE Inhibitory Activity of Enzymatic Hydrolysates from Ruditapes philippinarum. Molecules 2018, 23, 1189. [Google Scholar] [CrossRef] [PubMed]
- Fujita, H.; Yoshikawa, M. LKPNM: A prodrug-type ACE-inhibitory peptide derived from fish protein. Immunopharmacology 1999, 44, 123–127. [Google Scholar] [CrossRef]
- Hartmann, R.; Meisel, H. Food-derived peptides with biological activity: From research to food applications. Curr. Opin. Biotechnol. 2007, 18, 163–169. [Google Scholar] [CrossRef] [PubMed]
- Chinese Animal Science Editorial Board of Chinese Academy of Sciences. Chinese Zoology, Mollusc, Bivalve, Curcuma; Science Press: Beijing, China, 2001. [Google Scholar]
- Jiang, C.; Wang, M.; Liu, J.; Gan, D.; Zeng, X. Extraction, preliminary characterization, antioxidant and anticancer activities in vitro of polysaccharides from Cyclina sinensis. Carbohydr. Polym. 2011, 84, 851–857. [Google Scholar] [CrossRef]
- Jiang, C.; Xiong, Q.; Li, S.; Zhao, X.; Zeng, X. Structural characterization, sulfation and antitumor activity of a polysaccharide fraction from Cyclina sinensis. Carbohydr. Polym. 2015, 115, 200–206. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Xiong, Q.; Gan, D.; Jiao, Y.; Liu, J.; Ma, L.; Zeng, X. Antioxidant activity and potential hepatoprotective effect of polysaccharides from Cyclina sinensis. Carbohydr. Polym. 2013, 91, 262–268. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Zhu, W.; Xu, F.; Liu, M.; Xie, Y.; Zhang, J. Optimized ultrasonic-assisted extraction of polysaccharides from Cyclina sinensis and evaluation of antioxidant activities in vitro. CyTA-J. Food 2014, 12, 32–39. [Google Scholar] [CrossRef]
- Ren, Y.; Pan, H.; Yang, Y.; Pan, B.; Bu, W. Molecular cloning, characterization and functional analysis of a heat shock protein 70 gene in Cyclina sinensis. Fish Shellfish Immunol. 2016, 58, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Ren, Y.; Zhang, H.; Pan, B.; Gao, H. Identification and expression analysis of IκB and NF-κB genes from Cyclina sinensis. Fish Shellfish Immunol. 2016, 56, 427–435. [Google Scholar] [CrossRef] [PubMed]
- Daliri, E.B.-M.; Lee, B.H.; Oh, D.H. Current trends and perspectives of bioactive peptides. Crit. Rev. Food Sci. 2017, 2, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Eslie, W.D.; Cheryan, M. Continuous Enzymatic Modification of Proteins in an Ultrafiltration Reactor. J. Food Sci. 1981, 46, 1035–1042. [Google Scholar] [CrossRef]
- Picot, L.; Ravallec, R.; Fouchereau-Peron, M.; Vandanjon, L.; Jaouen, P.; Chaplain-Derouiniot, M.; Guerard, F.; Chabeaud, A.; Legal, Y.; Alvarez, O.M.; et al. Impact of ultrafiltration and nanofiltration of an industrial fish protein hydrolysate on its bioactive properties. J. Sci. Food Agric. 2010, 90, 1819–1826. [Google Scholar] [CrossRef] [PubMed]
- Abdelhedi, O.; Nasri, R.; Mora, L.; Jridi, M.; Toldrá, F.; Nasri, M. In silico analysis and molecular docking study of angiotensin I-converting enzyme inhibitory peptides from smooth-hound viscera protein hydrolysates fractionated by ultrafiltration. Food Chem. 2018, 239, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Xia, S.; Zhang, Y.; Li, X. Optimization of ACE inhibitory peptides from black soybean by microwave-assisted enzymatic method and study on its stability. LWT Food Sci. Technol. 2018, 98, 358–365. [Google Scholar] [CrossRef]
- Kumar, R.; Chaudhary, K.; Sharma, M.; Nagpal, G.; Chauhan, J.S.; Singh, S.; Gautam, A.; Raghava, G.P. AHTPDB: A comprehensive platform for analysis and presentation of antihypertensive peptides. Nucleic Acids Res. 2015, 43, D956–D962. [Google Scholar] [CrossRef] [PubMed]
- Minkiewicz, P.; Dziuba, J.; Iwaniak, A.; Dziuba, M.; Darewicz, M. BIOPEP database and other programs for processing bioactive peptide sequences. J. AOAC Int. 2008, 91, 965–980. [Google Scholar] [PubMed]
- Zamyatnin, A.A.; Borchikov, A.S.; Vladimirov, M.G.; Voronina, O.L. The EROP-Moscow oligopeptide database. Nucleic Acids Res. 2006, 34, D261–D266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sagardia, I.; Roa-Ureta, R.H.; Bald, C. A new QSAR model, for angiotensin I-converting enzyme inhibitory oligopeptides. Food Chem. 2013, 136, 1370–1376. [Google Scholar] [CrossRef] [PubMed]
- Pripp, A.H.; Isaksson, T.; Stepaniak, L.; Sørhaug, T. Quantitative structure-activity relationship modelling of ACE-inhibitory peptides derived from milk proteins. Eur. Food Res. Technol. 2004, 219, 579–583. [Google Scholar] [CrossRef]
- Pan, D.; Cao, J.; Guo, H.; Zhao, B. Studies on purification and the molecular mechanism of a novel ACE inhibitory peptide from whey protein hydrolysate. Food Chem. 2012, 130, 121–126. [Google Scholar] [CrossRef]
- He, R.; Malomo, S.A.; Alashi, A.; Girgih, A.T.; Ju, X.; Aluko, R.E. Purification and hypotensive activity of rapeseed protein-derived renin and angiotensin converting enzyme inhibitory peptides. J. Funct. Foods 2013, 5, 781–789. [Google Scholar] [CrossRef]
- Jing, P.; Qian, B.; He, Y.; Zhao, X.; Zhang, J.; Zhao, D.; Lv, Y.; Deng, Y. Screening milk-derived antihypertensive peptides using quantitative structure activity relationship (QSAR) modelling and in vitro/in vivo studies on their bioactivity. Int. Dairy J. 2014, 35, 95–101. [Google Scholar] [CrossRef]
- Wu, Q.; Jia, J.; Yan, H.; Du, J.; Gui, Z. A novel angiotensin-І converting enzyme (ACE) inhibitory peptide from gastrointestinal protease hydrolysate of silkworm pupa (Bombyx mori) protein: Biochemical characterization and molecular docking study. Peptides 2015, 68, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Du, J.; Jia, J.; Kuang, C. Production of ACE inhibitory peptides from sweet sorghum grain protein using alcalase: Hydrolysis kinetic, purification and molecular docking study. Food Chem. 2016, 199, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Moskowitz, D.W. Is “somatic” angiotensin I-converting enzyme a mechanosensor? Diabetes Technol. Ther. 2002, 4, 841–858. [Google Scholar] [CrossRef] [PubMed]
- Serrano, G.R. Binding of peptides substrates and inhibitorsof angiotensin-converting enzyme. J. Biol. Chem. 1980, 255, 401–407. [Google Scholar]
- Engelking, L.R. Chapter 6—Enzyme Kinetics. In Textbook of Veterinary Physiological Chemistry (Third Edition); Engelking, L.R., Ed.; Academic Press: Boston, MA, USA, 2015; pp. 32–38. [Google Scholar]
- Jao, C.L.; Huang, S.L.; Hsu, K.C. Angiotensin I-converting enzyme inhibitory peptides: Inhibition mode, bioavailability, and antihypertensive effects. BioMedicine 2012, 2, 130–136. [Google Scholar] [CrossRef]
- Xue, L.; Wang, X.; Hu, Z.; Wu, Z.; Wang, L.; Wang, H.; Yang, M. Identification and characterization of an angiotensin-converting enzyme inhibitory peptide derived from bovine casein. Peptides 2018, 99, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Wei, Y.N.; Chang, Q.; Sun, H.J.; Chai, K.G.; Huang, Z.Q.; Zhao, Z.X. Ultrafast screening of a novel, moderately hydrophilic angiotensin-converting-enzyme-inhibitory peptide, RYL, from silkworm pupa using an Fe-doped-silkworm-excrement-derived biocarbon: Waste conversion by waste. J. Agric. Food Chem. 2017, 65, 11202–11211. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, K.A.A.; David, S.M.; Murugesh, E.; Thandeeswaran, M.; Kiran, K.G.; Mahendran, R.; Palaniswamy, M.; Angayarkanni, J. Identification and in silico characterization of a novel peptide inhibitor of angiotensin converting enzyme from pigeon pea (Cajanus cajan). Phytomedicine 2017, 36, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Mancia, G. Angiotensin-converting enzyme inhibitors in the treatment of hypertension. J. Cardiovasc. Pharm. 1991, 18, S1–S3. [Google Scholar]
- Chel-Guerrero, L.; Domínguez-Magaña, M.; Martínez-Ayala, A.; Dávila-Ortiz, G.; Betancur-Ancona, D. Lima bean (Phaseolus lunatus) protein hydrolysates with ACE-I inhibitory activity. Food Nutr. Sci. 2012, 3, 11. [Google Scholar]
- Ketnawa, S.; Benjakul, S.; Martínez-Alvarez, O.; Rawdkuen, S. Fish skin gelatin hydrolysates produced by visceral peptidase and bovine trypsin: Bioactivity and stability. Food Chem. 2017, 215, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wang, X.W.; Zhai, J.; Li, X.; Huang, H.; Li, C.; Zheng, J.; Sun, H. Improvement to flooding risk assessment of storm surges by residual interpolation in the coastal areas of Guangdong Province, China. Quat. Int. 2017, 453, 1–14. [Google Scholar] [CrossRef]
- Singh, B.P.; Vij, S. In vitro stability of bioactive peptides derived from fermented soy milk against heat treatment, pH and gastrointestinal enzymes. LWT-Food Sci. Technol. 2018, 91, 303–307. [Google Scholar] [CrossRef]
- Plessas, S.; Nouska, C.; Karapetsas, A.; Kazakos, S.; Alexopoulos, A.; Mantzourani, I.; Chondrou, P.; Fournomiti, M.; Galanis, A.; Bezirtzoglou, E. Isolation, characterization and evaluation of the probiotic potential of a novel Lactobacillus strain isolated from Feta-type cheese. Food Chem. 2017, 226, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Vermeirssen, V.; Camp, J.V.; Verstraete, W. Bioavailability of angiotensin I converting enzyme inhibitory peptides. Br. J. Nutr. 2007, 92, 357–366. [Google Scholar] [CrossRef]
- Sheih, I.C.; Fang, T.J.; Wu, T.-K. Isolation and characterisation of a novel angiotensin I-converting enzyme (ACE) inhibitory peptide from the algae protein waste. Food Chem. 2009, 115, 279–284. [Google Scholar] [CrossRef]
- Chiang, W.D.; Tsou, M.J.; Tsai, Z.Y.; Tsai, T.C. Angiotensin I-converting enzyme inhibitor derived from soy protein hydrolysate and produced by using membrane reactor. Food Chem. 2006, 98, 725–732. [Google Scholar] [CrossRef]
- Tsai, J.-S.; Chen, J.-L.; Pan, B.S. ACE-inhibitory peptides identified from the muscle protein hydrolysate of hard clam (Meretrix lusoria). Process. Biochem. 2008, 43, 743–747. [Google Scholar] [CrossRef]
- Schulten, V.; Lauer, I.; Scheurer, S.; Thalhammer, T.; Bohle, B. A food matrix reduces digestion and absorption of food allergens in vivo. Mol. Nutr. Food Res. 2011, 55, 1484–1491. [Google Scholar] [CrossRef] [PubMed]
- Mouécoucou, J.; Frémont, S.; Sanchez, C.; Villaume, C.; Méjean, L. In vitro allergenicity of peanut after hydrolysis in the presence of polysaccharides. Clin. Exp. Allergy 2004, 34, 1429–1437. [Google Scholar] [CrossRef] [PubMed]
- Mouecoucou, J.; Villaume, C.; Sanchez, C.; Mejean, L. Beta-lactoglobulin/polysaccharide interactions during in vitro gastric and pancreatic hydrolysis assessed in dialysis bags of different molecular weight cut-offs. Bba-Gen. Subj. 2004, 1670, 105–112. [Google Scholar] [CrossRef]
- Nacer, S.A.; Sanchez, C.; Villaume, C.; Mejean, L.; Mouecoucou, J. Interactions between β-Lactoglobulin and Pectins during in Vitro Gastric Hydrolysis. J. Agric. Food Chem. 2004, 52, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.W.; Yang, Z.S.; Zhang, Y.R.; Wang, F.; Yu, F.M.; Ding, G.F. Process Optimization for Preparation of ACE-inhibitory Peptides from Cyclina sinensis by Enzymatic Hydrolysis. Food Ind. 2016, 37, 56–59. [Google Scholar]
- Cushman, D.W.; Cheung, H.S. Spectrophotometric assay and properties of the angiotensin-converting enzyme of rabbit lung. Biochem. Pharmacol. 1971, 20, 1637–1648. [Google Scholar] [CrossRef]
- Paiva, L.; Lima, E.; Neto, A.I.; Baptista, J. Isolation and characterization of angiotensin I-converting enzyme (ACE) inhibitory peptides from Ulva rigida C. Agardh protein hydrolysate. J. Funct. Foods 2016, 26, 65–76. [Google Scholar] [CrossRef]
- Forghani, B.; Zarei, M.; Ebrahimpour, A.; Philip, R.; Bakar, J.; Abdul, H.A.; Saari, N. Purification and characterization of angiotensin converting enzyme-inhibitory peptides derived from Stichopus horrens: Stability study against the ACE and inhibition kinetics. J. Funct. Foods 2016, 20, 276–290. [Google Scholar] [CrossRef]
- Lineweaver, H.; Burk, D. The Determination of Enzyme Dissociation Constants. J. Am. Chem. Soc. 1934, 56, 658–666. [Google Scholar] [CrossRef]
- Kittiphattanabawon, P.; Benjakul, S.; Visessanguan, W.; Shahidi, F. Gelatin hydrolysate from blacktip shark skin prepared using papaya latex enzyme: Antioxidant activity and its potential in model systems. Food Chem. 2012, 135, 1118–1126. [Google Scholar] [CrossRef] [PubMed]
- Commission, N.P. Pharmacopoeia of People’s Republic of China; China Medical Science Press: Beijing, China, 2015. [Google Scholar]
- Yu, Z.P.; Zhao, W.Z.; Yu, Y.D.; Liu, B.Q.; Liu, J.B. Stability of Antihypertensive Peptides from Egg White Protein to Chemicals and Enzymes. Food Sci. 2010, 31, 23–26. [Google Scholar]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.M.; Huey, R.; Lindstrom, W.; Sanner, M.F.; Belew, R.K.; Goodsell, D.S.; Olson, A.J. AutoDock4 and AutoDockTools4: Automated docking with selective receptor flexibility. J. Comput. Chem. 2009, 30, 2785–2791. [Google Scholar] [CrossRef] [PubMed] [Green Version]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, F.; Zhang, Z.; Luo, L.; Zhu, J.; Huang, F.; Yang, Z.; Tang, Y.; Ding, G. Identification and Molecular Docking Study of a Novel Angiotensin-I Converting Enzyme Inhibitory Peptide Derived from Enzymatic Hydrolysates of Cyclina sinensis. Mar. Drugs 2018, 16, 411. https://doi.org/10.3390/md16110411
Yu F, Zhang Z, Luo L, Zhu J, Huang F, Yang Z, Tang Y, Ding G. Identification and Molecular Docking Study of a Novel Angiotensin-I Converting Enzyme Inhibitory Peptide Derived from Enzymatic Hydrolysates of Cyclina sinensis. Marine Drugs. 2018; 16(11):411. https://doi.org/10.3390/md16110411
Chicago/Turabian StyleYu, Fangmiao, Zhuangwei Zhang, Liwang Luo, Junxiang Zhu, Fangfang Huang, Zuisu Yang, Yunping Tang, and Guofang Ding. 2018. "Identification and Molecular Docking Study of a Novel Angiotensin-I Converting Enzyme Inhibitory Peptide Derived from Enzymatic Hydrolysates of Cyclina sinensis" Marine Drugs 16, no. 11: 411. https://doi.org/10.3390/md16110411
APA StyleYu, F., Zhang, Z., Luo, L., Zhu, J., Huang, F., Yang, Z., Tang, Y., & Ding, G. (2018). Identification and Molecular Docking Study of a Novel Angiotensin-I Converting Enzyme Inhibitory Peptide Derived from Enzymatic Hydrolysates of Cyclina sinensis. Marine Drugs, 16(11), 411. https://doi.org/10.3390/md16110411