Heterologous Expression of a Thermostable β-1,3-Galactosidase and Its Potential in Synthesis of Galactooligosaccharides

Abstract
:1. Introduction
2. Results
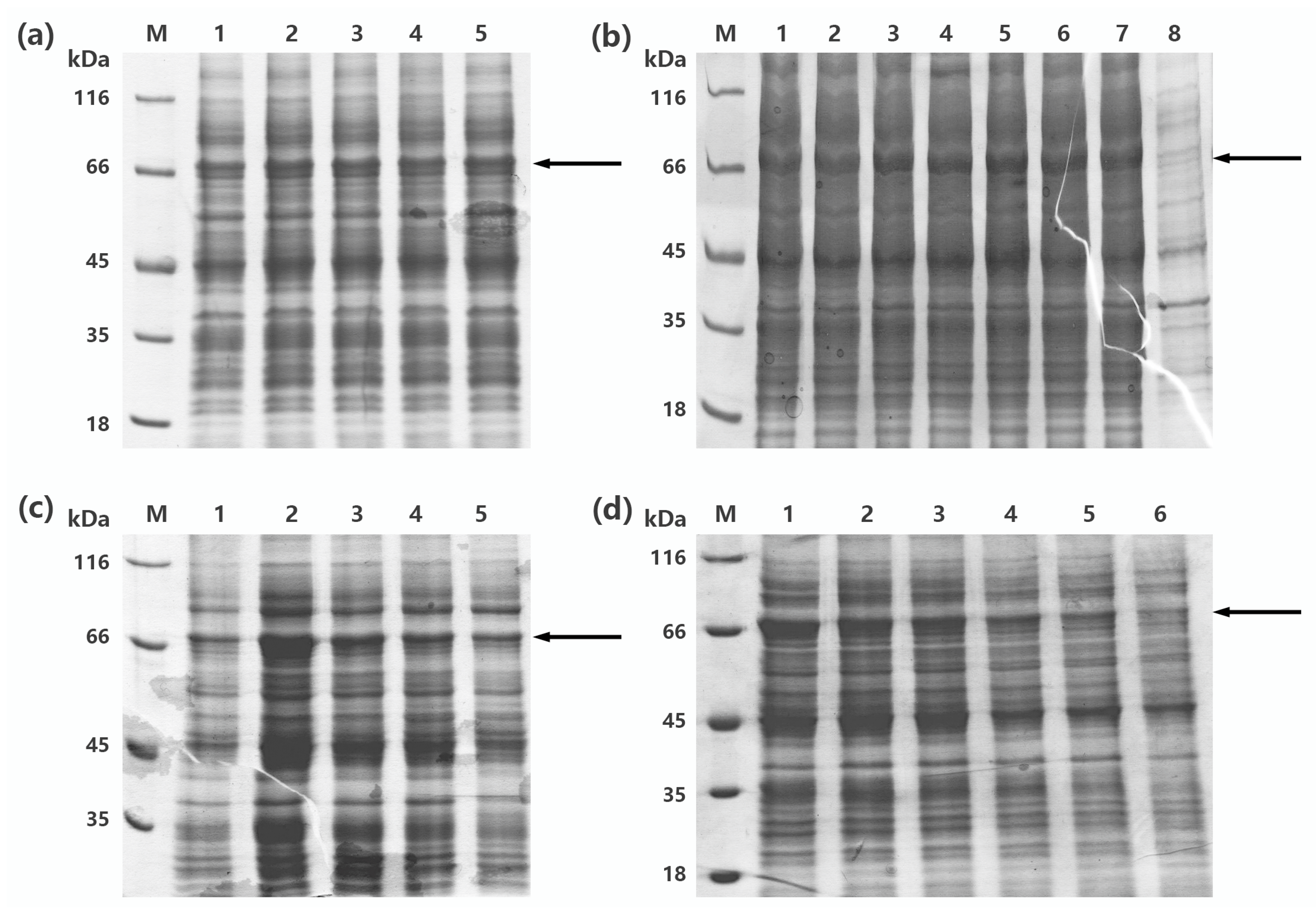
2.1. Construction of Recombinant Cells
2.2. Optimization of the Expression Condition of rMaBGA
2.3. Purification of rMaBGA
2.4. Enzymatic Characterization of MaBGA
2.4.1. Effect of pH and Temperature on the Activity of rMaBGA
2.4.2. Thermal Denaturation Kinetic of rMaBGA
2.4.3. Steady-State Kinetic of rMaBGA
2.5. Synthesis of Galactooligosaccharides
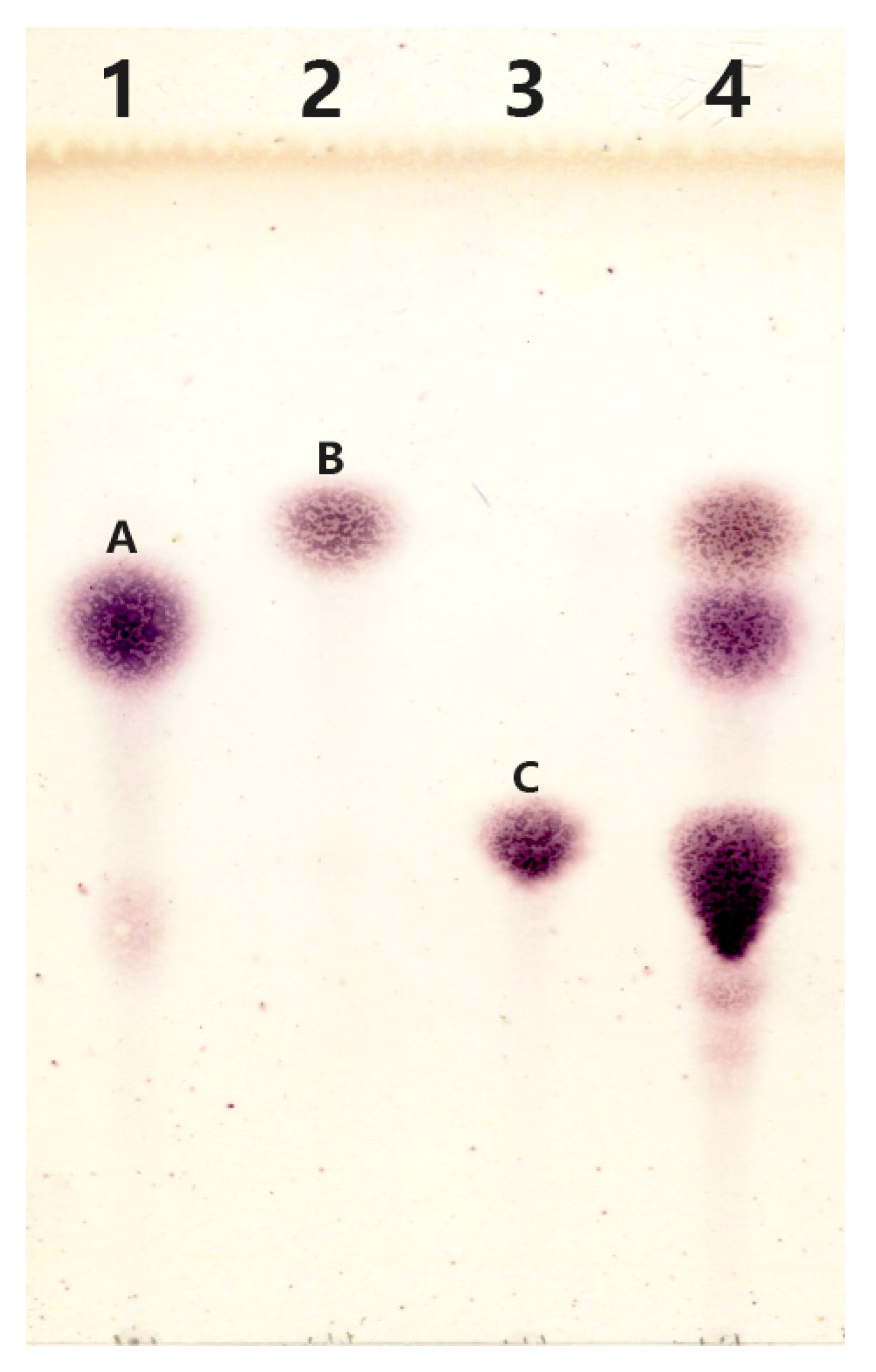
2.5.1. Thin-Layer Chromatography Analysis
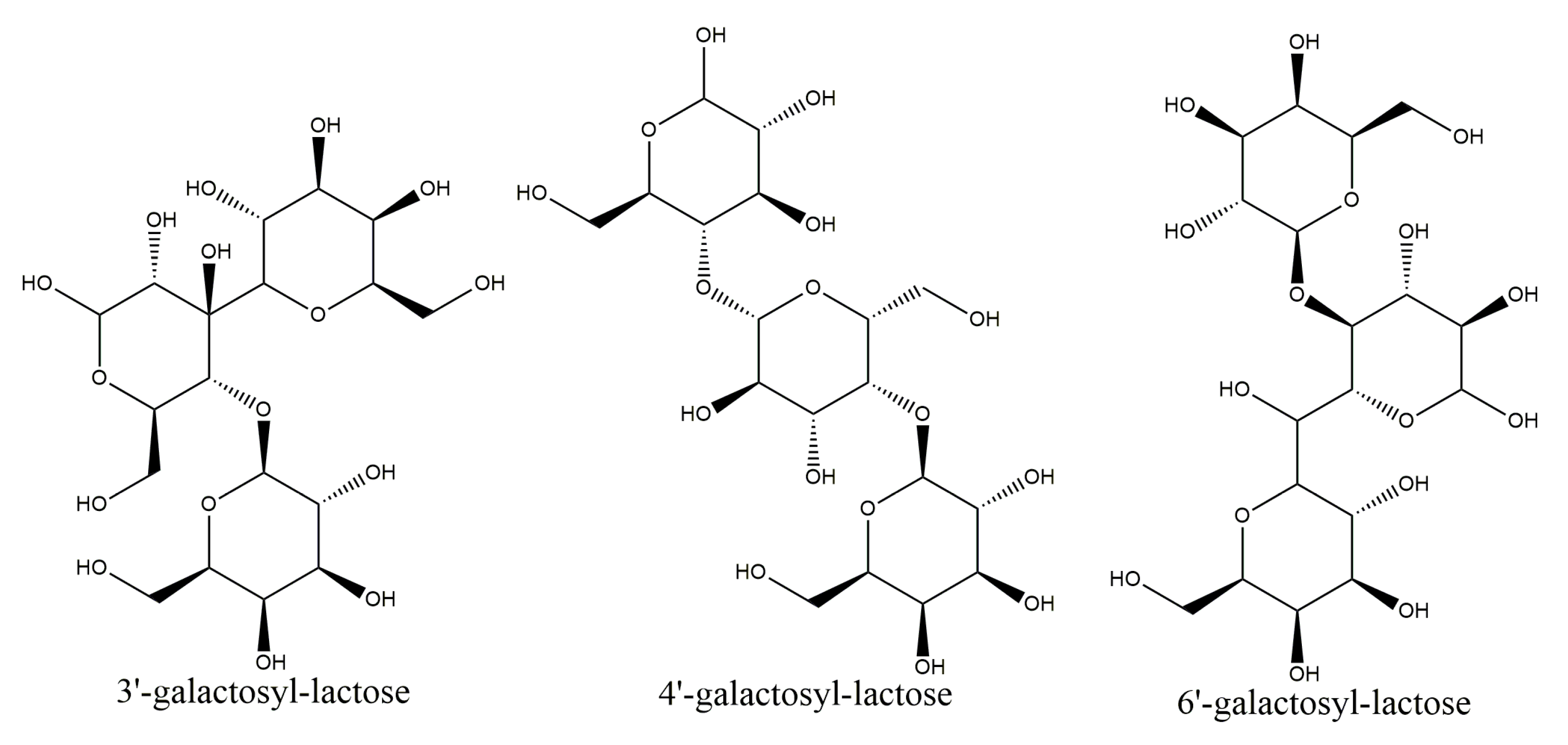
2.5.2. Characterization of the Products of Transglycosylation
3. Discussion
4. Materials and Methods
4.1. Expression and Purification of rMaBGA
4.1.1. Strains, Plasmids, and Culture Conditions
4.1.2. Construction of Recombinant Strains
4.1.3. Optimization of the Production of rMaBGA
4.1.4. Expression and Purification of rMaBGA
4.1.5. SDS-PAGE Analysis
4.2. Enzymatic Characterization of rMaBGA
4.2.1. Enzyme Activity Assay
4.2.2. Effect of pH and Temperature on the Activity of rMaBGA
4.2.3. Thermal Denaturation Kinetic of rMaBGA
4.2.4. Steady-State Kinetic of rMaBGA
4.3. Synthesis of Galactooligosaccharides
4.3.1. Thin-Layer Chromatography Analysis
4.3.2. Characterization of the Products of Transglycosylation
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Panesar, P.S.; Kaur, R.; Singh, R.S.; Kennedy, J.F. Biocatalytic strategies in the production of galacto-oligosaccharides and its global status. Int. J. Biol. Macromol. 2018, 111, 667–679. [Google Scholar] [CrossRef] [PubMed]
- Holscher, H.D. Dietary fiber and prebiotics and the gastrointestinal microbiota. Gut Microbes 2017, 8, 172–184. [Google Scholar] [CrossRef] [PubMed]
- So, S.S.; Wan, M.L.; El-Nezami, H. Probiotics-mediated suppression of cancer. Curr. Opin. Oncol. 2017, 29, 62–72. [Google Scholar] [CrossRef] [PubMed]
- Intanon, M.; Arreola, S.L.; Pham, N.H.; Kneifel, W.; Haltrich, D.; Nguyen, T.H. Nature and biosynthesis of galacto-oligosaccharides related to oligosaccharides in human breast milk. Fems. Microbiol. Lett. 2014, 353, 89–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vera, C.; Cordova, A.; Aburto, C.; Guerrero, C.; Suarez, S.; Illanes, A. Synthesis and purification of galacto-oligosaccharides: State of the art. World J. Microbiol. Biotechnol. 2016, 32, 197. [Google Scholar] [CrossRef] [PubMed]
- Park, A.R.; Oh, D.K. Galacto-oligosaccharide production using microbial beta-galactosidase: Current state and perspectives. Appl. Microbiol. Biotechnol. 2010, 85, 1279–1286. [Google Scholar] [CrossRef] [PubMed]
- Rastall, R.A. Functional oligosaccharides: Application and manufacture. Annu. Rev. Food Sci. Technol. 2010, 1, 305–339. [Google Scholar] [CrossRef] [PubMed]
- Saqib, S.; Akram, A.; Halim, S.A.; Tassaduq, R. Sources of beta-galactosidase and its applications in food industry. Biotechnology 2017, 7, 79. [Google Scholar]
- Oliveira, C.; Guimaraes, P.M.; Domingues, L. Recombinant microbial systems for improved beta-galactosidase production and biotechnological applications. Biotechnol. Adv. 2011, 29, 600–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bras, N.F.; Fernandes, P.A.; Ramos, M.J. QM/MM Studies on the beta-Galactosidase Catalytic Mechanism: Hydrolysis and Transglycosylation Reactions. J. Chem. Theory Comput. 2010, 6, 421–433. [Google Scholar] [CrossRef] [PubMed]
- Chanalia, P.; Gandhi, D.; Attri, P.; Dhanda, S. Purification and characterization of beta-galactosidase from probiotic Pediococcus acidilactici and its use in milk lactose hydrolysis and galactooligosaccharide synthesis. Bioorg. Chem. 2018, 77, 176–189. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Pijning, T.; Meng, X.; Dijkhuizen, L.; van Leeuwen, S.S. Biochemical Characterization of the Functional Roles of Residues in the Active Site of the beta-Galactosidase from Bacillus circulans ATCC 31382. Biochemistry 2017, 56, 3109–3118. [Google Scholar] [CrossRef] [PubMed]
- Carneiro, L.; Yu, L.; Dupree, P.; Ward, R.J. Characterization of a beta-galactosidase from Bacillus subtilis with transgalactosylation activity. Int. J. Biol. Macromol. 2018, 120, 279–287. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Zeng, Q.; Zhou, L.; Yu, Y.; Chen, B. Biochemical and Structural Insights into a Novel Thermostable beta-1,3-Galactosidase from Marinomonas sp. BSi20414. Mar. Drugs 2017, 15, 13. [Google Scholar] [CrossRef] [PubMed]
- Jayaprakash, N.G.; Surolia, A. Role of glycosylation in nucleating protein folding and stability. Biochem. J. 2017, 474, 2333–2347. [Google Scholar] [CrossRef] [PubMed]
- Nishi, H.; Shaytan, A.; Panchenko, A.R. Physicochemical mechanisms of protein regulation by phosphorylation. Front. Genet. 2014, 5, 270. [Google Scholar] [CrossRef] [PubMed]
- Murn, J.; Shi, Y. The winding path of protein methylation research: Milestones and new frontiers. Nat. Rev. Mol. Cell Biol. 2017, 18, 517–527. [Google Scholar] [CrossRef] [PubMed]
- Shental-Bechor, D.; Levy, Y. Effect of glycosylation on protein folding: A close look at thermodynamic stabilization. Proc. Natl. Acad Sci. USA 2008, 105, 8256–8261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-Colinas, B.; Fernandez-Arrojo, L.; Ballesteros, A.O.; Plou, F.J. Galactooligosaccharides formation during enzymatic hydrolysis of lactose: Towards a prebiotic-enriched milk. Food Chem. 2014, 145, 388–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Otieno, D.O. Synthesis of β-Galactooligosaccharides from Lactose Using Microbial β-Galactosidases. Compr. Rev. Food Sci. Food Saf. 2010, 9, 471–482. [Google Scholar] [CrossRef] [Green Version]
- Depeint, F.; Tzortzis, G.; Vulevic, J.; I’Anson, K.; Gibson, G.R. Prebiotic evaluation of a novel galactooligosaccharide mixture produced by the enzymatic activity of Bifidobacterium bifidum NCIMB 41171, in healthy humans: A randomized, double-blind, crossover, placebo-controlled intervention study. Am. J. Clin. Nutr. 2008, 87, 785–791. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Wang, Y.; Sun, K.; Yu, Y.; Chen, B. Preliminary studies on the screening, identification and optimum fermentative conditions of a strain Marinomonas sp. BSi20414 isolated from arctic sea ice producing β-galactosidase. J. Polar Res. 2011, 108–114. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Gao, F.; Liu, D.; Li, Z.; Xu, X.; Wu, M.; Zhao, Y. Significant improvement of thermal stability of glucose 1-dehydrogenase by introducing disulfide bonds at the tetramer interface. Enzym. Microb. Technol. 2013, 53, 365–372. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | Temperature (°C) | kd (h−1) | t1/2 (h) | ΔH (KJ mol−1) | ΔG (KJ mol−1) | ΔS (J mol−1 K−1) |
|---|---|---|---|---|---|---|
| wtMaBGA | 50 | 0.0433 | 16.00 | 114.03 | 87.75 | 81.35 |
| 60 | 0.1597 | 4.34 | 113.94 | 86.94 | 81.10 | |
| rMaBGA | 50 | 0.0721 | 9.61 | 147.75 | 86.38 | 189.99 |
| 60 | 0.3879 | 1.79 | 147.66 | 84.48 | 189.74 |
| Enzyme | Km (mM) | Vmax (μM min−1) |
|---|---|---|
| rMaBGA | 6.85 | 64.13 |
| wtMaBGA | 14.19 | 1.05 |
| Unit | α-glucosyl | β-glucosyl | β-galactosyl-A | β-galactosyl-B |
|---|---|---|---|---|
| 1 | 5.14 | 4.59 | 4.37 | 4.50 |
| 2 | 3.52 | 3.21 | 3.62 | 3.62 |
| 3 | 3.79 | 3.42 | 3.84 | 3.70 |
| 4 | 3.50 | 3.46 | 4.01 | 3.86 |
| 5 | 3.85 | 3.60 | 3.70 | 3.67 |
| 6 | 3.75–3.88 | 3.75–3.88 | 3.70–3.85 | 3.70–3.85 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, H.; Zhou, L.; Zeng, Q.; Yu, Y.; Chen, B. Heterologous Expression of a Thermostable β-1,3-Galactosidase and Its Potential in Synthesis of Galactooligosaccharides. Mar. Drugs 2018, 16, 415. https://doi.org/10.3390/md16110415
Ding H, Zhou L, Zeng Q, Yu Y, Chen B. Heterologous Expression of a Thermostable β-1,3-Galactosidase and Its Potential in Synthesis of Galactooligosaccharides. Marine Drugs. 2018; 16(11):415. https://doi.org/10.3390/md16110415
Chicago/Turabian StyleDing, Haitao, Lili Zhou, Qian Zeng, Yong Yu, and Bo Chen. 2018. "Heterologous Expression of a Thermostable β-1,3-Galactosidase and Its Potential in Synthesis of Galactooligosaccharides" Marine Drugs 16, no. 11: 415. https://doi.org/10.3390/md16110415
APA StyleDing, H., Zhou, L., Zeng, Q., Yu, Y., & Chen, B. (2018). Heterologous Expression of a Thermostable β-1,3-Galactosidase and Its Potential in Synthesis of Galactooligosaccharides. Marine Drugs, 16(11), 415. https://doi.org/10.3390/md16110415