Chemically-Induced Production of Anti-Inflammatory Molecules in Microalgae

 ,
,  and
and
Abstract
:1. Microalgae as Source of Anti-Inflammatory Compounds
2. Chemically-Induced Oxidative Stress to Improve Production of Anti-Inflammatory Compounds
2.1. Chemicals Triggering Accumulation of Anti-Inflammatory Compounds
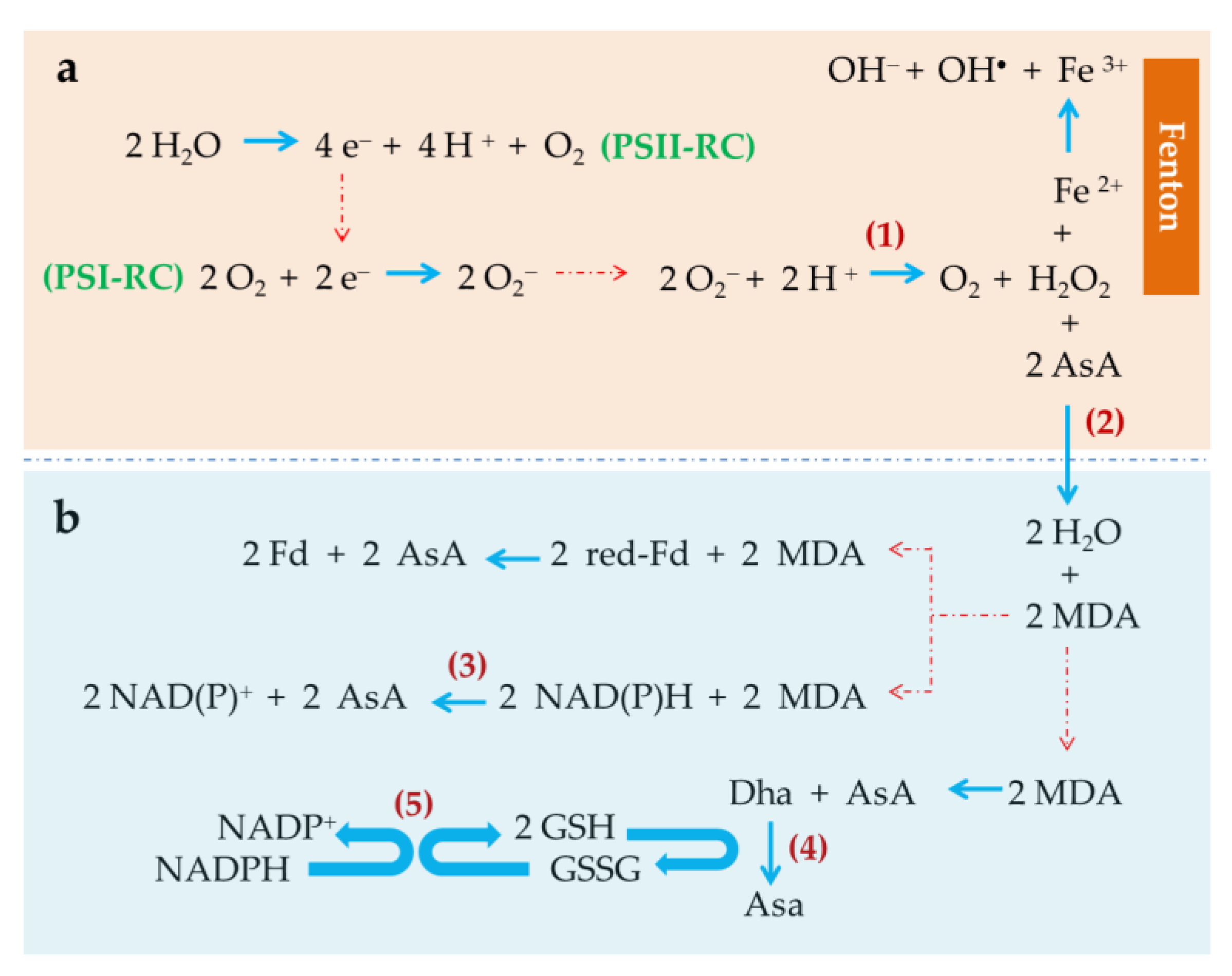
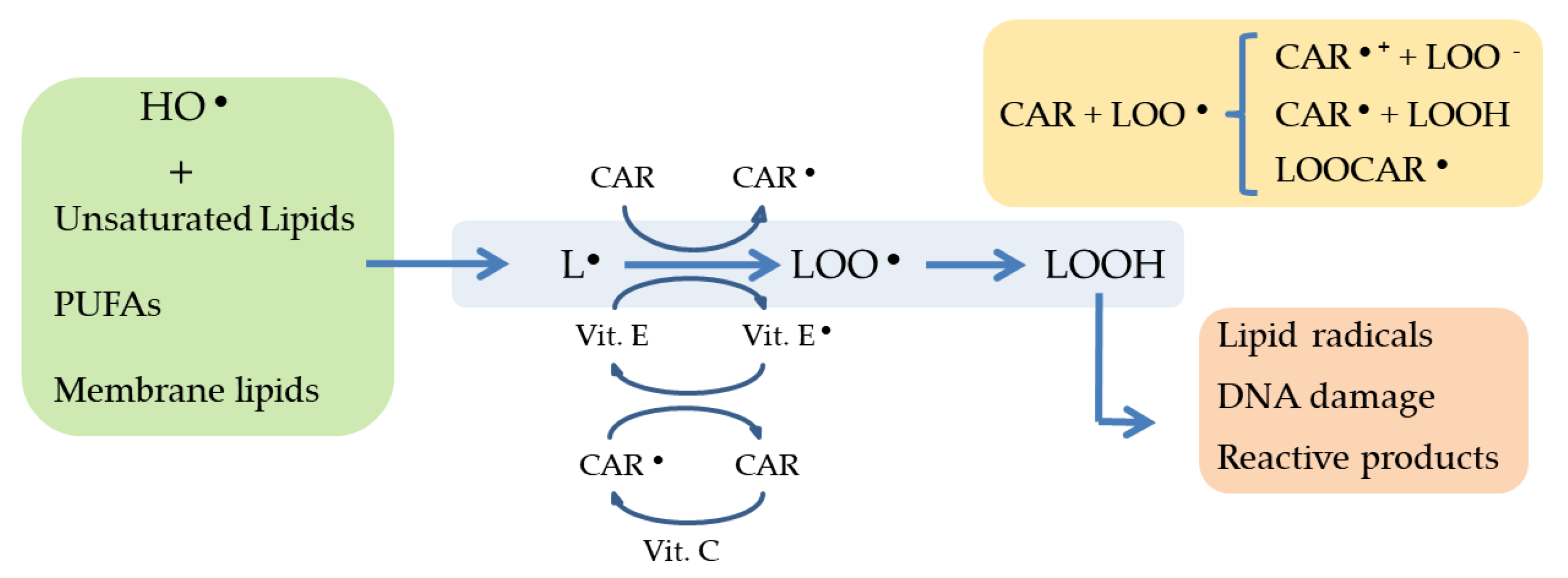
2.2. Induced-Oxidative Stress: A Key Strategy to Trigger Accumulation of Anti-Inflammatory Compounds
2.3. Production of Pigments with Anti-Inflammatory Activity
2.4. Production of Polyunsaturated Fatty Acids (PUFA) with Anti-Inflammatory Activity
2.5. Production of Sulphated Polysaccharides (sPS) with Anti-Inflammatory Activity
2.6. Production of Phenolic Compounds with Anti-Inflammatory Activity
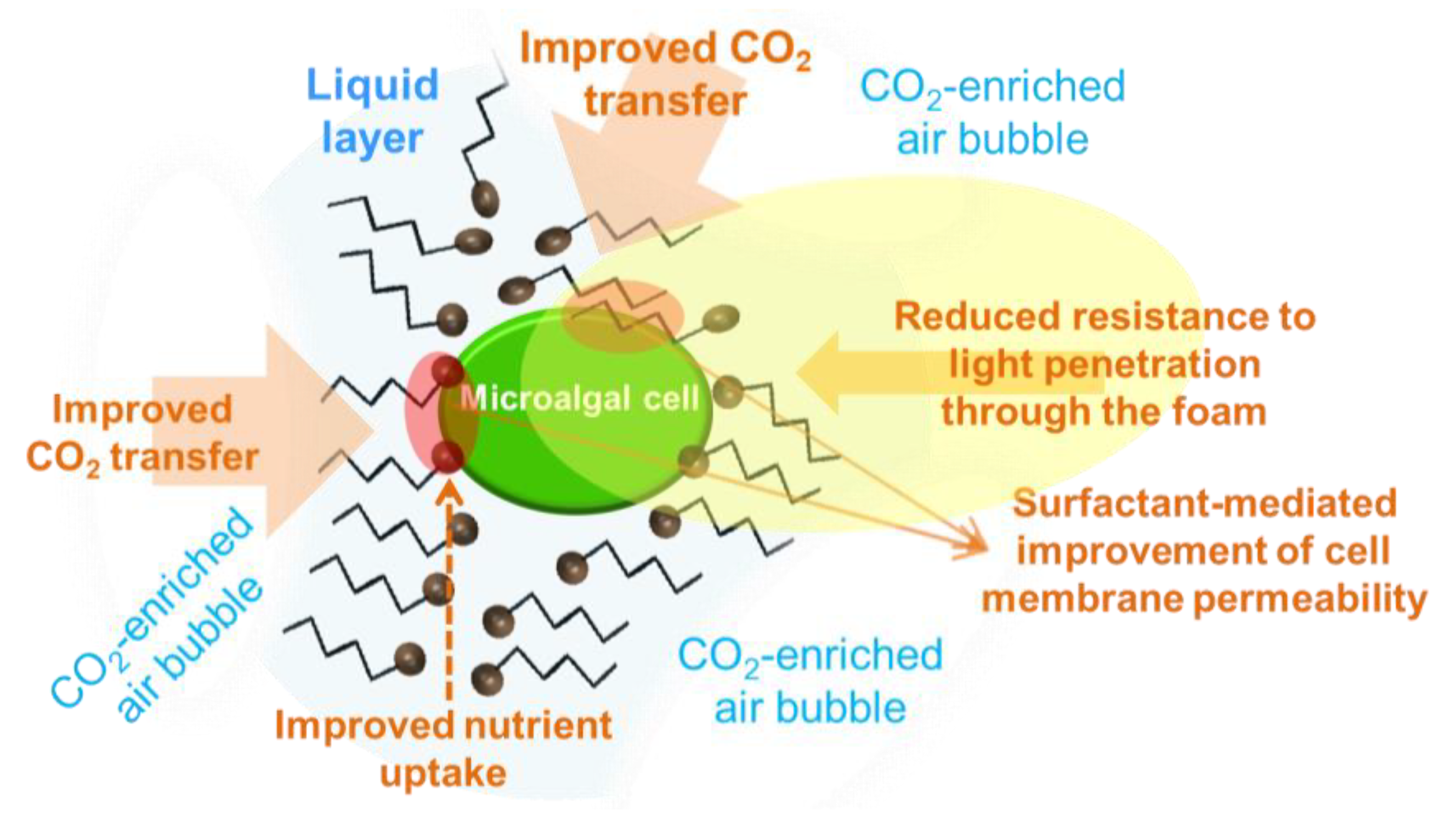
3. Surfactant-Facilitated Accumulation of Anti-Inflammatory Molecules
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Singh, P.; Kumari, S.; Guldhe, A.; Misra, R.; Ismail, R.; Rawat, I.; Bux, F. Trends and novel strategies for enhancing lipid accumulation and quality in microalgae. Renew. Sustain. Energy Rev. 2016, 55, 1–16. [Google Scholar] [CrossRef]
- Thomas, N.V.; Kim, S.K. Beneficial effects of marine algal compounds in cosmeceuticals. Mar. Drugs 2013, 11, 146–164. [Google Scholar] [CrossRef] [PubMed]
- Forján, E.; Navarro, F.; Cuaresma, M.; Vaquero, I.; Ruíz-Domínguez, M.C.; Gojkovic, Ž.; Vázquez, M.; Márquez, M.; Mogedas, B.; Bermejo, E.; et al. Microalgae: Fast-growth sustainable green factories. Crit. Rev. Environ. Sci. Technol. 2015, 45, 1705–1755. [Google Scholar] [CrossRef]
- Gong, M.; Bassi, A. Carotenoids from microalgae: A review of recent developments. Biotechnol. Adv. 2016, 34, 1396–1412. [Google Scholar] [CrossRef] [PubMed]
- Khozin-Goldberg, I.; Iskandarov, U.; Cohen, Z. LC-PUFA from photosynthetic microalgae: occurrence, biosynthesis, and prospects in biotechnology. Appl. Microbiol. Biotechnol. 2011, 91, 905–915. [Google Scholar] [CrossRef] [PubMed]
- Uttaro, A.D. Biosynthesis of polyunsaturated fatty acids in lower eukaryotes. Life 2006, 58, 563–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Talero, E.; García-Mauriño, S.; Ávila-Román, J.; Rodríguez-Luna, A.; Alcaide, A.; Motilva, V. Bioactive compounds isolated from microalgae in chronic inflammation and cancer. Mar. Drugs 2015, 13, 6152–6209. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Chen, L.; Zhang, W. Chemicals to enhance microalgal growth and accumulation of high-value bioproducts. Front. Microbiol. 2015, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.H.; Jiang, J.G.; Chen, Q. Influence of daily collection and culture medium recycling on the growth and β-carotene yield of Dunaliellasalina. J. Agric. Food Chem. 2008, 56, 4027–4031. [Google Scholar] [CrossRef] [PubMed]
- Panis, G.; Carreon, J.R. Commercial astaxanthin production derived by green alga Haematococcuspluvialis: A microalgae process model and a techno-economic assessment all through production line. Algal Res. 2016, 18, 175–190. [Google Scholar] [CrossRef]
- Pérez, G.; Doldán, S.; Scavone, P.; Borsani, O.; Irisarri, P. Osmotic stress alters UV-based oxidative damage tolerance in a heterocyst forming cyanobacterium. Plant Physiol. Biochem. 2016, 108, 231–240. [Google Scholar] [CrossRef] [PubMed]
- de Farias Silva, C.E.; Sforza, E. Carbohydrate productivity in continuous reactor under nitrogen limitation: Effect of light and residence time on nutrient uptake in Chlorella vulgaris. Process Biochem. 2016, 51, 2112–2118. [Google Scholar] [CrossRef]
- Steinrücken, P.; Prestegard, S.K.; de Vree, J.H.; Storesund, J.E.; Pree, B.; Mjøs, S.A.; Erga, S.R. Comparing EPA production and fatty acid profiles of three Phaeodactylum tricornutum strains under western Norwegian climate conditions. Algal Res. 2018, 30, 11–12. [Google Scholar] [CrossRef] [PubMed]
- Cesário, M.T.; da Fonseca, M.M.R.; Marques, M.M.; de Almeida, M.C.M.D. Marine algal carbohydrates as carbon sources for the production of biochemicals and biomaterials. Biotechnol. Adv. 2018, 36, 798–817. [Google Scholar] [CrossRef] [PubMed]
- Vílchez, C.; Forján, E.; Cuaresma, M.; Bédmar, F.; Garbayo, I.; Vega, J.M. Marine carotenoids: biological functions and commercial applications. Mar. Drugs 2011, 9, 319–333. [Google Scholar] [CrossRef] [PubMed]
- Enzing, C.; Ploeg, M.; Barbosa, M.; Sijtsma, L. Microalgae-based products for the food and feed sector: an outlook for Europe. In JRC Scientific and Policy Reports, 1st ed.; Vigani, M., Parisi, C., Rodriguez-Cerezo, E., Eds.; Publications Office of the European Union: Luxembourg, 2014; pp. 1–82. ISBN 978-92-79-34037-6. [Google Scholar]
- Robertson, R.C.; Guihéneuf, F.; Bahar, B.; Schmid, M.; Stengel, D.B.; Fitzgerald, G.F.; Ross, R.P.; Stanton, C. The anti-inflammatory effect of algae-derived lipid extracts on lipopolysaccharide (LPS)-stimulated human THP-1 macrophages. Mar. Drugs 2015, 13, 5402–5424. [Google Scholar] [CrossRef] [PubMed]
- Safafar, H.; van Wagenen, J.; Møller, P.; Jacobsen, C. Carotenoids, phenolic compounds and tocopherols contribute to the antioxidative properties of some microalgae species grown on industrial wastewater. Mar. Drugs 2015, 13, 7339–7356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takaichi, S. Carotenoids in algae: distributions, biosyntheses and functions. Mar. Drugs 2011, 9, 1101–1118. [Google Scholar] [CrossRef] [PubMed]
- Goiris, K.; Muylaert, K.; Fraeye, I.; Foubert, I.; de Brabanter, J.; de Cooman, L. Antioxidant potential of microalgae in relation to their phenolic and carotenoid content. J. Appl. Phycol. 2012, 24, 1477–1486. [Google Scholar] [CrossRef]
- Blagojević, D.; Babić, O.; Rašeta, M.; Šibul, F.; Janjušević, L.; Simeunović, J. Antioxidant activity and phenolic profile in filamentous cyanobacteria: The impact of nitrogen. J. Appl. Phycol. 2018, 30, 2337. [Google Scholar] [CrossRef]
- Norsker, N.; Barbosa, M.J.; Vermuë, M.H.; Wijffels, R.H. Microalgal production—A close look at the economics. Biotechnol. Adv. 2011, 29, 24–27. [Google Scholar] [CrossRef] [PubMed]
- Richmond, A. Handbook of Microalgae Culture: Biotechnology and Phycology; Blackwell Science: Cambridge, UK, 2004. [Google Scholar]
- Varshney, P.; Mikulic, P.; Vonshak, A.; Beardall, J.; Wangikar, P.P. Extremophilic micro-algae and their potential contribution in biotechnology. Bioresour. Technol. 2015, 184, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Ramos, A.A.; Polle, J.J.; Tran, D.; Cushman, J.C.; Jin, E.; Varela, J.C. The unicellular green alga Dunaliella salina Teod. as a model for abiotic stress tolerance: Genetic advances and future perspectives. Algae 2011, 26, 3–20. [Google Scholar] [CrossRef]
- Yuan, J.P.; Peng, J.; Yin, K.; Wang, J.H. Potential health-promoting effects of astaxanthin: A high-value carotenoid mostly from microalgae. Mol. Nutr. Food Res. 2011, 55, 150–165. [Google Scholar] [CrossRef] [PubMed]
- Shi, X.M.; Chen, F. High-yield production of lutein by the green microalga Chlorella protothecoides in heterotrophic fed-batch culture. Biotechnol. Progr. 2002, 18, 723–727. [Google Scholar] [CrossRef] [PubMed]
- Lagarde, D.; Beuf, L.; Vermaas, W. Increased production of zeaxanthin and other pigments by application of genetic engineering techniques to Synechocystis sp. strain PCC 6803. Appl. Environ. Microbiol. 2000, 66, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Romay, Ch.; González, R.; Ledón, N.; Remirez, D.; Rimbau, V. C-phycocyanin: A biliprotein with antioxidant, anti-inflammatory and neuroprotective effects. Curr. Protein Pept. Sci. 2003, 4, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Fiedor, J.; Burda, K. Potential role of carotenoids as antioxidants in human health and disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef] [PubMed]
- Richard, D.; Kefi, K.; Barbe, U.; Bausero, P.; Visioli, F. Polyunsaturated fatty acids as antioxidants. Pharmacol. Res. 2008, 57, 451–455. [Google Scholar] [CrossRef] [PubMed]
- De Jesus Raposo, M.F.; de Morais, R.M.S.C.; de Morais, A.M.M.B. Bioactivity and applications of sulphated polysaccharides from marine microalgae. Mar. Drugs 2015, 11, 233–252. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wang, C.; Shi, Q.; Ma, C. Preparation of different molecular weight polysaccharides from Porphyridium cruentum and their antioxidant activities. Int. J. Biol. Macromol. 2009, 45, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; You, B.; Huang, J.; Yu, Y.; Chen, W. Isolation and antioxidant property of the extracellular polysaccharide from Rhodella reticulata. World J. Microbiol. Biotechnol. 2010, 26, 833–840. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, X. Separation, antitumor activities, and encapsulation of polypeptide from Chlorella pyrenoidosa. Biotechnol. Progr. 2013, 29, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Asada, K. Production and scavenging of reactive oxygen species in chloroplasts and their functions. Plant Physiol. 2006, 141, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Janoska, A.; Vázquez, M.; Janssen, M.; Wijffels, R.H.; Cuaresma, M.; Vílchez, C. Surfactant selection for a liquid foam-bed photobioreactor. Biotechnol. Progr. 2018. [Google Scholar] [CrossRef] [PubMed]
- Franz, A.; Danielewicz, M.; Wong, D.; Anderson, L.; Boothe, J. Phenotypic screening with oleaginous microalgae reveals modulators of lipid productivity. ACS Chem. Biol. 2013, 8, 1053–1062. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, M.; Kakizono, T.; Nagai, S. Enhanced carotenoid biosynthesis by oxidative stress in acetate-induced cyst cells of a green unicellular alga, Haematococcus pluvialis. Appl. Environ. Microbiol. 1993, 59, 867–873. [Google Scholar] [PubMed]
- Zheng, Y.; Li, Z.; Tao, M.; Li, J.; Hu, Z. Effects of selenite on green microalga Haematococcus pluvialis: Bioaccumulation of selenium and enhancement of astaxanthin production. Aquat. Toxicol. 2017, 183, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Chen, F. Induction of astaxanthin formation by reactive oxygen species in mixotrophic culture of Chlorococcum sp. Biotechnol. Lett. 2001, 23, 519–523. [Google Scholar] [CrossRef]
- Wang, Y.; Liu, Z.; Qin, S. Effects of iron on fatty acid and astaxanthin accumulation in mixotrophic Chromochloris zofingiensis. Biotechnol. Lett. 2013, 35, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Raman, V.; Ravi, S. Effect of salicylic acid and methyl jasmonate on antioxidant systems of Haematococcus pluvialis. Acta Physiol. Plant 2010, 33, 1043–1049. [Google Scholar] [CrossRef]
- Kobayashi, M. In vivo antioxidant role of astaxanthin under oxidative stress in the green alga Haematococcus pluvialis. Appl. Microbiol. Biotechnol. 2000, 54, 550–555. [Google Scholar] [CrossRef] [PubMed]
- Pirastru, L.; Darwish, M.; Chu, F.L.; Perreault, F.; Sirois, L.; Sleno, L.; Popovic, R. Carotenoid production and change of photosynthetic functions in Scenedesmus sp. exposed to nitrogen limitation and acetate treatment. J. Appl. Phycol. 2012, 24, 117–124. [Google Scholar] [CrossRef]
- Boussiba, S.; Fan, L.; Vonshak, A. Enhancement and determination of astaxanthin accumulation in green alga Haematococcus pluvialis. Methods Enzymol. 1992, 213, 386–391. [Google Scholar] [CrossRef]
- Ip, P.F.; Chen, F. Employment of reactive oxygen species to enhance astaxanthin formation in Chlorella zofingiensis in heterotrophic culture. Process Biochem. 2005, 40, 3491–3496. [Google Scholar] [CrossRef]
- Wei, D.; Chen, F.; Chen, G.; Zhang, X.; Liu, L.; Zhang, H. Enhanced production of lutein in heterotrophic Chlorella protothecoides by oxidative stress. Sci. China C Life Sci. 2008, 51, 1088–1093. [Google Scholar] [CrossRef] [PubMed]
- Vaquero, I.; Ruiz-Domínguez, M.C.; Márquez, M.; Vílchez, C. Cu-mediated biomass productivity enhancement and lutein enrichment of the novel microalga Coccomyxa onubensis. Process Biochem. 2012, 47, 694–700. [Google Scholar] [CrossRef]
- Ruiz-Domínguez, M.C.; Vaquero, I.; Obregón, V.; de la Morena, B.; Vílchez, C.; Vega, J.M. Lipid accumulation and antioxidant activity in the eukaryotic acidophilic microalga Coccomyxa sp. (strain onubensis) under nutrient starvation. J. Appl. Phycol. 2015, 27, 1099–1108. [Google Scholar] [CrossRef]
- Ambati, R.R.; Ravi, S.; Aswathanarayana, R.G. Enhancement of carotenoids in green alga Botryococcus braunii in various autotrophic media under stress conditions. Int. J. Biomed. Pharm. Sci. 2010, 4, 87–92. [Google Scholar]
- Carvalho, A.P.; Malcata, F.X. Effect of culture media on production of polyunsaturated fatty acids by Pavlova lutheri. Cryptogam. Algol. 2000, 21, 59–71. [Google Scholar] [CrossRef]
- Meng, Y.; Jiang, J.; Wang, H.; Cao, X.; Xue, S.; Yang, Q.; Wang, W. The characteristics of TAG and EPA accumulation in Nannochloropsis oceanica IMET1 under different nitrogen supply regimes. Bioresour. Technol. 2015, 179, 483–489. [Google Scholar] [CrossRef] [PubMed]
- Qu, L.; Ji, X.J.; Ren, L.J.; Nie, Z.K.; Feng, Y.; Wu, W.J.; Ouyang, P.K.; Huang, H. Enhancement of docosahexaenoic acid production by Schizochytrium sp. using a two stage oxygen supply control strategy based on oxygen transfer coefficient. Lett. Appl. Microbiol. 2011, 52, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Tonon, T.; Harvey, D.; Larson, T.R.; Graham, I.A. Long chain polyunsaturated fatty acid production and partitioning to triacylglycerols in four microalgae. Phytochemistry 2002, 61, 15–24. [Google Scholar] [CrossRef]
- Villay, A.; Laroche, C.; Roriz, D.; El Alaoui, H.; Delbac, F.; Michaud, P. Optimisation of culture parameters for exopolysaccharides production by the microalga Rhodella violacea. Bioresour. Technol. 2013, 146, 732–735. [Google Scholar] [CrossRef] [PubMed]
- Abdullahi, A.S.; Underwood, G.J.C.; Gretz, M.R. Extracellular matrix assembly in diatoms (bacillariophyceae). V. Environmental effects on polysaccharide synthesis in the model diatom, Phaeodactylum tricornutum. J. Phycol. 2006, 42, 363–378. [Google Scholar] [CrossRef]
- Mishra, A.; Jha, B. Isolation and characterization of extracellular polymeric substances from micro-algae Dunaliella salina under salt stress. Bioresour. Technol. 2009, 100, 3382–3386. [Google Scholar] [CrossRef] [PubMed]
- López, A.; Rico, M.; Santana-Casiano, J.M.; González, A.G.; González-Dávila, M. Phenolic profile of Dunaliella tertiolecta growing under high levels of copper and iron. Environ. Sci. Pollut. Res. 2015, 22, 14820. [Google Scholar] [CrossRef] [PubMed]
- El-Baky, H.H.A.; El Baz, F.K.; El-Baroty, G.S. Phenolics from Spirulina maxima: Over-production and in vitro protective effect of its phenolics on CCl4 induced hepatotoxicity. J. Med. Plants Res. 2009, 3, 24–30. [Google Scholar] [CrossRef]
- Cirulis, J.T.; Scott, J.A.; Ross, G.M. Management of oxidative stress by microalgae. Can. J. Physiol. Pharmacol. 2013, 91, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Tripathy, B.C.; Oelmüller, R. Reactive oxygen species generation and signaling in plants. Plant Signal Behav. 2012, 7, 1621–1633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehler, A.H. Studies on reactivities of illuminated chloroplasts. I. Mechanism of the reduction of oxygen and other Hill reagents. Arch. Biochem. Biophys. 1951, 33, 65–77. [Google Scholar] [CrossRef]
- Asada, K.; Kiso, K.; Yoshikawa, K. Univalent reduction of molecular oxygen by spinach chloroplasts on illumination. J. Biol. Chem. 1974, 249, 2175–2181. [Google Scholar] [PubMed]
- Arulselvan, P.; Fard, M.T.; Tan, W.S.; Gothai, S.; Fakurazi, S.; Norhaizan, M.E.; Kumar, S.S. Role of antioxidants and natural products in inflammation. Oxid. Med. Cell Longev. 2016, 5276130. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Meng, C.; Zhang, X.; Xu, D.; Miao, X.; Wang, Y.; Yang, L.; Lv, H.; Chen, L.; Ye, N. Induction of salicylic acid (SA) on transcriptional expression of eight carotenoid genes and astaxanthin accumulation in Haematococcus pluvialis. Enzyme Microb. Technol. 2012, 51, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Lamers, P.P.; Janssen, M.; De Vos, R.C.H.; Bino, R.J.; Wijffels, R.H. Carotenoid and fatty acid metabolism in nitrogen-starved Dunaliella salina, a unicellular green microalga. J. Biotechnol. 2012, 162, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Le Marchand, L.; Hankin, J.H.; Kolonel, L.N.; Beecher, G.R.; Wilkens, L.R.; Zhao, L.P. Intake of specific carotenoids and lung cancer risk. Cancer Epidemiol. Biomarkers Prev. 1993, 2, 183–187. [Google Scholar] [PubMed]
- Biesalski, H. Evidence from Intervention Studies. In Functions of Vitamins beyond Recommended Dietary Allowances; Walter, P., Hornig, D., Moser, U., Eds.; Woodhead Publishing Limited: Cambridge, UK, 2001; pp. 92–134. [Google Scholar]
- Guerin, M.; Huntley, M.E.; Olaizola, M. Haematococcus astaxanthin: Applications for human health and nutrition. Trends Biotechnol. 2003, 21, 210–216. [Google Scholar] [CrossRef]
- Landrum, J.T.; Bohne, R. Luteín, zeaxanthin and the macular pigment. Arch. Biochem. Biophys. 2001, 385, 28–40. [Google Scholar] [CrossRef] [PubMed]
- Friedman, D.S.; O’Colmain, B.J.; Muñoz, B.; Tomany, S.C.; McCarty, C.; De Jong, P.T.; Nemesure, B.; Mitchell, P.; Kempen, J.; Congdon, N. Prevalence of age-related macular degeneration in the United States. Arch. Ophthalmol. 2004, 122, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Hammond, B.R.; Wooten, B.; Snodderly, D.M. Preservation of visual sensitivity of older individuals: Association with macular pigment density. Invest. Ophthalmol. Vis. Sci. 1998, 39, 397–406. [Google Scholar] [PubMed]
- Lee, S.J.; Bai, S.K.; Lee, K.S.; Namkoong, S.; Na, H.J.; Ha, K.S.; Han, J.A.; Yim, S.V.; Chang, K.; Kwon, Y.G.; et al. Astaxanthin inhibits nitric oxide production and inflammatory gene expression by suppressing IκB kinase-dependent NF-κB activation. Mol. Cells 2003, 16, 97–105. [Google Scholar] [PubMed]
- Akyön, Y. Effect of antioxidants on the immune response of Helicobacter pylori. Clin. Microbiol. Infect. 2002, 8, 438–441. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Jean, D.; Lim, Y.P.; Lim, C.; An, G. Weight gain limitation and liver protection by long-term feeding of astaxanthin in murines. J. K. Soc. Appl. Biol. Chem. 2009, 52, 180–185. [Google Scholar] [CrossRef]
- Bolin, A.P.; Macedo, R.C.; Marin, D.P.; Barros, M.P.; Otton, R. Astaxanthin prevents in vitro auto-oxidative injury in human lymphocytes. Cell Biol. Toxicol. 2010, 26, 457–467. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M. Method of Inhibiting 5α-Reductase with Astaxanthin to Prevent and Treat Benign Prostate Hyperplasia (BPH) and Prostate Cancer in Human Males. U.S. Patent No. 6277417, 21 August 2001. [Google Scholar]
- Zuluaga, M.; Gueguen, V.; Pavon-Djavid, G.; Letourneur, D. Carotenoids from microalgae to block oxidative stress. BioImpacts 2017, 7, 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olofsson, P.; Hultqvist, M.; Hellgren, L.I.; Holmdahl, R. Phytol: A chlorophyll component with anti-inflammatory and metabolic properties. In Recent Advances in Redox Active Plant and Microbial Products; Jacob, C., Kirsch, G., Slusarenko, A., Winyard, P., Burkholz, T., Eds.; Springer: Dordrecht, The Netherlands, 2014. [Google Scholar]
- Cuaresma, M.; Janssen, M.; Vílchez, C.; Wijffels, R.H. Productivity of Chlorella sorokiniana in a short light-path (SLP) panel photobioreactor under high irradiance. Biotechnol. Bioeng. 2009, 104, 352–359. [Google Scholar] [CrossRef] [PubMed]
- Beyer, P.; Kleinig, H. On the desaturation and cyclization reactions of carotenes. In Chromoplast Membranes, Carotenoids; Krinsky, N., Mathews-Roth, M., Taylor, R., Eds.; Springer: New York, NY, USA, 1989; pp. 195–206. [Google Scholar]
- Harker, M.; Tsavalos, A.J.; Young, A.J. Factors responsible for astaxanthin formation in the chlorophyte Haematococcus pluvialis. Bioresour. Technol. 1996, 55, 207–214. [Google Scholar] [CrossRef]
- Nahidian, B.; Ghanati, F.; Shahbazi, M.; Soltani, N. Effect of nutrients on the growth and physiological features of newly isolated Haematococcus pluvialis TMU. Bioresour. Technol. 2018, 255, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Minhas, A.K.; Hodgson, P.; Barrow, C.J.; Adholeya, A. A review on the assessment of stress conditions for simultaneous production of microalgal lipids and carotenoids. Front. Microbiol. 2016, 7, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Serini, S.; Bizzarro, A.; Piccioni, E.; Fasano, E.; Rossi, C.; Lauria, A.; Cittadini, A.; Masullo, C.; Calviello, G. EPA and DHA differentially affect in vitro inflammatory cytokine release by peripheral blood mononuclear cells from Alzheimer’s patients. Curr. Alzheimer Res. 2012, 9, 913–923. [Google Scholar] [CrossRef] [PubMed]
- Das, U.N. Can vagus nerve stimulation halt or ameliorate rheumatoid arthritis and lupus? Lipids Health Dis. 2011, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Van Beelen, V.A.; Spenkelink, B.; Mooibroek, H.; Sijtsma, L.; Bosch, D.; Rietjens, I.M.C.M.; Alink, G.M. An n-3 PUFA-rich microalgal oil diet protects to a similar extent as a fish oil-rich diet against AOM-induced colonic aberrant crypt foci in F344 rats. Food Chem. Toxicol. 2009, 47, 316–320. [Google Scholar] [CrossRef] [PubMed]
- Halliwell, B. Cellular stress and protection mechanisms. Biochem. Soc. Trans. 1996, 24, 1023–1027. [Google Scholar] [CrossRef] [PubMed]
- Kiritsakis, A.K.; Dugan, L.R. Studies in photooxidation of olive oil. J. Am. Oil Chem. Soc. 1985, 62, 892–896. [Google Scholar] [CrossRef]
- Li, M.; Hu, C.; Zhu, Q.; Chen, L.; Kong, Z.; Liu, Z. Copper and zinc induction of lipid peroxidation and effects on antioxidant enzyme activities in the microalga Pavlova viridis (Prymnesiophyceae). Chemosphere 2006, 62, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Li, H.B.; Cheng, K.W.; Wong, C.C.; Fan, K.W.; Chen, F.; Jiang, Y. Evaluation of antioxidant capacity and total phenolic content of different fractions of selected microalgae. Food Chem. 2007, 102, 771–776. [Google Scholar] [CrossRef]
- Yilancioglu, K.; Cokol, M.; Pastirmaci, I.; Erman, B.; Cetiner, S. Oxidative stress is a mediator for increased lipid accumulation in a newly isolated Dunaliella salina strain. PLoS ONE 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Li, T.; Wu, N.; Ren, L.; Jiang, L.; Ji, X.; Huang, H. Mechanism of arachidonic acid accumulation during aging in Mortierella alpina: A large-scale label-free comparative proteomics study. J. Agric. Food Chem. 2016, 64, 9124–9134. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.Y.; Lu, W.C.; Chu, I.M. A fermentation strategy for producing docosahexaenoic acid in Aurantiochytrium limacinum SR21 and increasing C22:6 proportions in total fatty acid. Bioresour. Technol. 2012, 123, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Ruenwai, R.; Neiss, A.; Laoteng, K.; Vongsangnak, W.; Dalfard, A.B.; Cheevadhanarak, S.; Petranovic, D.; Nielsen, J. Heterologous production of polyunsaturated fatty acids in Saccharomyces cerevisiae causes a global transcriptional response resulting in reduced proteasomal activity and increased oxidative stress. Biotechnol. J. 2011, 6, 343–356. [Google Scholar] [CrossRef] [PubMed]
- Chokshi, K.; Pancha, I.; Ghosh, A.; Mishra, S. Salinity induced oxidative stress alters the physiological responses and improves the biofuel potential of green microalgae Acutodesmus dimorphus. Bioresour. Technol. 2017, 244, 1376–1383. [Google Scholar] [CrossRef] [PubMed]
- Qiao, H.; Cong, C.; Sun, C.; Li, B.; Wang, J.; Zhang, L. Effect of culture conditions on growth, fatty acid composition and DHA/EPA ratio of Phaeodactylum tricornutum. Aquaculture 2016, 452, 311–317. [Google Scholar] [CrossRef]
- Dahmen-Ben Moussa, I.; Chtourou, H.; Karray, F.; Sayadi, S.; Dhouib, A. Nitrogen or phosphorus repletion strategies for enhancing lipid or carotenoid production from Tetraselmis marina. Bioresour. Technol. 2017, 238, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Tsai, H.P.; Chuang, L.T.; Chen, C.N.N. Production of long chain omega-3 fatty acids and carotenoids in tropical areas by a new heat-tolerant microalga Tetraselmis sp. DS3. Food Chem. 2016, 192, 682–690. [Google Scholar] [CrossRef] [PubMed]
- Burg, A.; Oshrat, L.O. Salt effect on the antioxidant activity of red microalgal sulfated polysaccharides in soy-bean formula. Mar. Drugs 2015, 13, 6425–6439. [Google Scholar] [CrossRef] [PubMed]
- Fisher, T.; Berner, T.; Iluz, D.; Dubinsky, Z. The kinetics of the photoacclimation response of Nannochloropsis sp. (Eustigmatophyceae): A study of changes in ultrastructure and PSU density. J. Phycol. 1998, 34, 818–824. [Google Scholar] [CrossRef]
- Guzmán, S.; Gato, A.; Lamela, M.; Freire-Garabal, M.; Calleja, J.M. Anti-inflammatory and immunomodulatory activities of polysaccharide from Chlorella stigmatophora and Phaeodactylum tricornutum. Phytother. Res. 2003, 17, 665–670. [Google Scholar] [CrossRef] [PubMed]
- Matsui, M.S.; Muizzuddin, N.; Arad, S.; Marenus, K. Sulfated polysaccharides from red microalgae have antiinflammatory properties in vitro and in vivo. Appl. Biochem. Biotechnol. 2003, 104, 13–22. [Google Scholar] [CrossRef]
- Delattre, C.; Pierre, G.; Laroche, C.; Michaud, P. Production, extraction and characterization of microalgal and cyanobacterial exopolysaccharides. Biotechnol. Adv. 2016, 34, 1159–1179. [Google Scholar] [CrossRef] [PubMed]
- Passow, U. Transparent exopolymer particles (TEP) in aquatic environments. Prog. Oceanogr. 2002, 55, 287–333. [Google Scholar] [CrossRef] [Green Version]
- Tannin-Spitz, T.; Bergman, M.; Van-Moppes, D.; Grossman, S.; Arad, S. Antioxidant activity of the polysaccharide of the red microalga Porphyridium sp. J. Appl. Phycol. 2005, 17, 215–222. [Google Scholar] [CrossRef]
- Markou, G.; Nerantzis, E. Microalgae for high-value compounds and biofuels production: A new focus on cultivation under stress conditions. Biotechnol. Adv. 2013, 31, 1532–1542. [Google Scholar] [CrossRef] [PubMed]
- Chi, Z.; Su, C.D.; Lu, W.D. A new exopolysaccharide produced by marine Cyanothece sp. 113. Bioresour. Technol. 2007, 98, 1329–1332. [Google Scholar] [CrossRef] [PubMed]
- Underwood, G.J.C.; Boulcot, M.; Raines, C.A.; Waldron, K. Environmental effects on exopolymer production by marine benthic diatoms–dynamics, changes in composition and pathways of production. J. Phycol. 2004, 40, 293–304. [Google Scholar] [CrossRef]
- Guerrini, F.; Cangini, M.; Boni, L.; Trost, P.; Pistocchi, R. Metabolic responses of the diatom Achnanthes brevipes (Bacillariophyceae) to nutrient limitation. J. Phycol. 2000, 36, 882–890. [Google Scholar] [CrossRef]
- Alcoverro, T.; Conte, E.; Mazzella, L. The production of mucilage by the epipelic diatom Cylindrotheca closterium from the Adriatic Sea under nutrient limitation. J. Phycol. 2000, 36, 1087–1095. [Google Scholar] [CrossRef]
- Kumar, A.S.; Mody, K.; Jha, B. Bacterial exopolysaccharides–a perception. J. Basic Microbiol. 2007, 47, 103–117. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Zhenming, C.; Weidong, L. Optimization of medium and cultivation conditions for enhanced exopolysaccharide yield by marine Cyanothece sp. 113. Chin. J. Oceanol. Limnol. 2007, 25, 411–417. [Google Scholar] [CrossRef]
- Ozturk, S.; Aslim, B. Modification of exopolysaccharide composition and production by three cyanobacterial isolates under salt stress. Environ. Sci. Pollut. Res. 2010, 17, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Sanjeewa, K.K.A.; Fernando, I.P.S.; Samarakoon, K.W.; Lakmal, H.H.C.; Kim, E.A.; Kwon, O.N.; Dilshara, M.G.; Lee, J.B.; Jeon, Y.J. Anti-inflammatory and anti-cancer activities of sterol rich fraction of cultured marine microalga Nannochloropsis oculata. Algae 2016, 31, 277–287. [Google Scholar] [CrossRef]
- Hämäläinen, M.; Nieminen, R.; Vuorela, P.; Heinonen, M.; Moilanen, E. Anti-inflammatory effects of flavonoids: genistein, kaempferol, quercetin, and daidzein inhibit STAT-1 and NF-κB activations, whereas flavone, isorhamnetin, naringenin, and pelargonidin inhibit only NF-κB activation along with their inhibitory effect on iNOS expression and NO production in activated macrophages. Mediat. Inflamm. 2007. [Google Scholar] [CrossRef]
- Janoska, A.; Lamers, P.P.; Hamhuis, A.; van Eimeren, Y.; Wijffels, R.H.; Janssen, M. A liquid foam-bed photobioreactor for microalgae production. Chem. Eng. J. 2017, 313, 1206–1214. [Google Scholar] [CrossRef]
- Janssen, M.; Lamers, P.P.; de Haan, M.; Wijffels, R.H. Growing Microalgae or Cyanobacteria in Liquid-Based Foam. International Patent WO 2014072294 A1, 15 May 2014. [Google Scholar]
- Vázquez, M.; Fuentes, J.L.; Hincapié, A.; Garbayo, I.; Vílchez, C.; Cuaresma, M. Selection of microalgae with potential for cultivation in surfactant-stabilized foam. Algal Res. 2018, 31, 216–224. [Google Scholar] [CrossRef]
- Taoka, Y.; Nagano, N.; Okita, Y.; Izumida, H.; Sugimoto, S.; Hayashi, M. Effect of Tween 80 on the growth, lipid accumulation and fatty acid composition of Thraustochytrium aureum ATCC 34304. J. Biosci. Bioeng. 2011, 111, 420–424. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.M.; Geng, L.J.; Ren, L.J.; Ji, X.J.; Hao, N.; Chen, K.Q.; Huang, H. Influence of oxygen on the biosynthesis of polyunsaturated fatty acids in microalgae. Bioresour. Technol. 2018, 250, 868–876. [Google Scholar] [CrossRef] [PubMed]
- Bailey, R.B.; Dimasi, D.; Hansen, J.M.; Mirrasoul, P.J.; Ruecker, C.M.; Iii, G.T.; Kaneko, T.; Barclay, W.R. Enhanced Production of Lipids Containing Polyenoic Fatty Acid by Very High Density Cultures of Eukaryotic Microbes in Fermentors. US Patent 8206956, 29 May 2012. [Google Scholar]
- Ren, L.J.; Ji, X.J.; Huang, H.; Qu, L.; Feng, Y.; Tong, Q.Q.; Ouyang, P.K. Development of a stepwise aeration control strategy for efficient docosahexaenoic acid production by Schizochytrium sp. Appl. Microbiol. Biotechnol. 2010, 87, 1649–1656. [Google Scholar] [CrossRef] [PubMed]
- Shelest, E.; Heimerl, N.; Fichtner, M.; Sasso, S. Multimodular type I polyketide synthases in algae evolve by module duplications and displacement of AT domains in trans. BMC Genomics 2015, 16, 1015. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Li, Y.; Wen, H.; Ren, X.; Liu, J.; Yang, L. Investigation on the role of surfactants in bubble-algae interaction in flotation harvesting of Chlorella vulgaris. Sci. Rep. 2018, 8, 3303. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Biochemical Group | Microalgae | References |
|---|---|---|
| Carotenoids | Haematococcus pluvialis Dunaliella salina Chlorella sorokiniana Synechocystis sp. | [9,10] |
| PUFA | Phaeodactylum tricornutum Nannochloropsis gaditana | [11] |
| Carbohydrates | Chlorella vulgaris Phaeodactylum tricornutum Porphyridium sp. Tetraselmis suecica | [12,13,14] |
| Anti-Inflammatory Molecule | Chemical Trigger | Induction Mechanism | Microalgae | References |
|---|---|---|---|---|
| Astaxanthin | H2O2, SeO32, Fe (II) | Oxidative stress | Haematococcus pluvialis | [39,40] |
| MV | Oxidative stress | Chlorococcum sp. | [41] | |
| Fe (II) | Oxidative stress | Cromochloris zofingiensis | [42] | |
| Jasm., salic. acid | Oxidative stress | Haematococcus pluvialis | [43] | |
| N starvation, NaCl | Oxidative stress | Haematococcus pluvialis | [44,45,46] | |
| Lutein | H2O2, NaClO | Oxidative stress | Chlorella zoofingiensis | [47,48] |
| Fe (II), Cu (II) | Oxidative stress | Coccomyxa onubensis | [3,24,49] | |
| N starvation | Oxidative stress | Coccomyxa onubensis | [50] | |
| NaCl | Oxidative stress | Botryococcus braunii | [51] | |
| EPA | Low oxygen | PUFA stimulation | Pavlova lutheri | [52] |
| N, P repletion | PUFA stimulation | Nannochloropsis oceanica | [53] | |
| DHA | Low oxygen | PUFA stimulation | Schizochytrium sp. | [54] |
| N, P repletion | PUFA stimulation | Pavlova lutheri | [55] | |
| Sulphated polysaccharides | N starvation | Oxidative stress | Rhodella violacea | [56] |
| P starvation | Oxidative stress | Phaeodactylum tricornutum | [57] | |
| NaCl | Oxidative stress | Dunaliella salina | [58] | |
| Phenolic compounds | Cu (II), Fe (II) | Oxidative stress | Dunaliella tertiolecta | [59] |
| N repletion + Phe | Phenylpr. synth. | Spirulina platensis | [60] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montero-Lobato, Z.; Vázquez, M.; Navarro, F.; Fuentes, J.L.; Bermejo, E.; Garbayo, I.; Vílchez, C.; Cuaresma, M. Chemically-Induced Production of Anti-Inflammatory Molecules in Microalgae. Mar. Drugs 2018, 16, 478. https://doi.org/10.3390/md16120478
Montero-Lobato Z, Vázquez M, Navarro F, Fuentes JL, Bermejo E, Garbayo I, Vílchez C, Cuaresma M. Chemically-Induced Production of Anti-Inflammatory Molecules in Microalgae. Marine Drugs. 2018; 16(12):478. https://doi.org/10.3390/md16120478
Chicago/Turabian StyleMontero-Lobato, Zaida, María Vázquez, Francisco Navarro, Juan Luis Fuentes, Elisabeth Bermejo, Inés Garbayo, Carlos Vílchez, and María Cuaresma. 2018. "Chemically-Induced Production of Anti-Inflammatory Molecules in Microalgae" Marine Drugs 16, no. 12: 478. https://doi.org/10.3390/md16120478
APA StyleMontero-Lobato, Z., Vázquez, M., Navarro, F., Fuentes, J. L., Bermejo, E., Garbayo, I., Vílchez, C., & Cuaresma, M. (2018). Chemically-Induced Production of Anti-Inflammatory Molecules in Microalgae. Marine Drugs, 16(12), 478. https://doi.org/10.3390/md16120478