Blueprints for the Next Generation of Bioinspired and Biomimetic Mineralised Composites for Bone Regeneration

 , and
, and 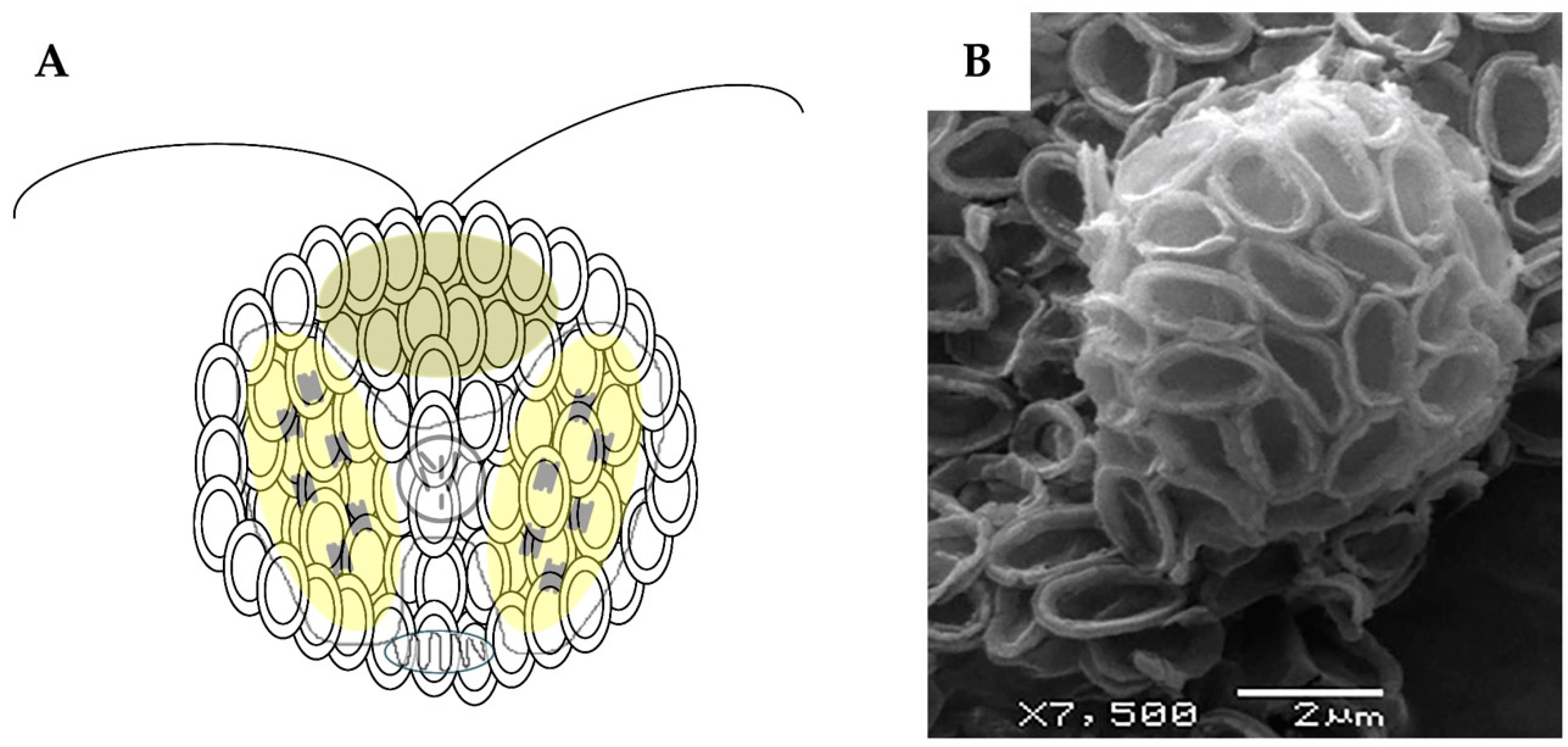{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Coccolithophores
3. Coccolithophore Organics
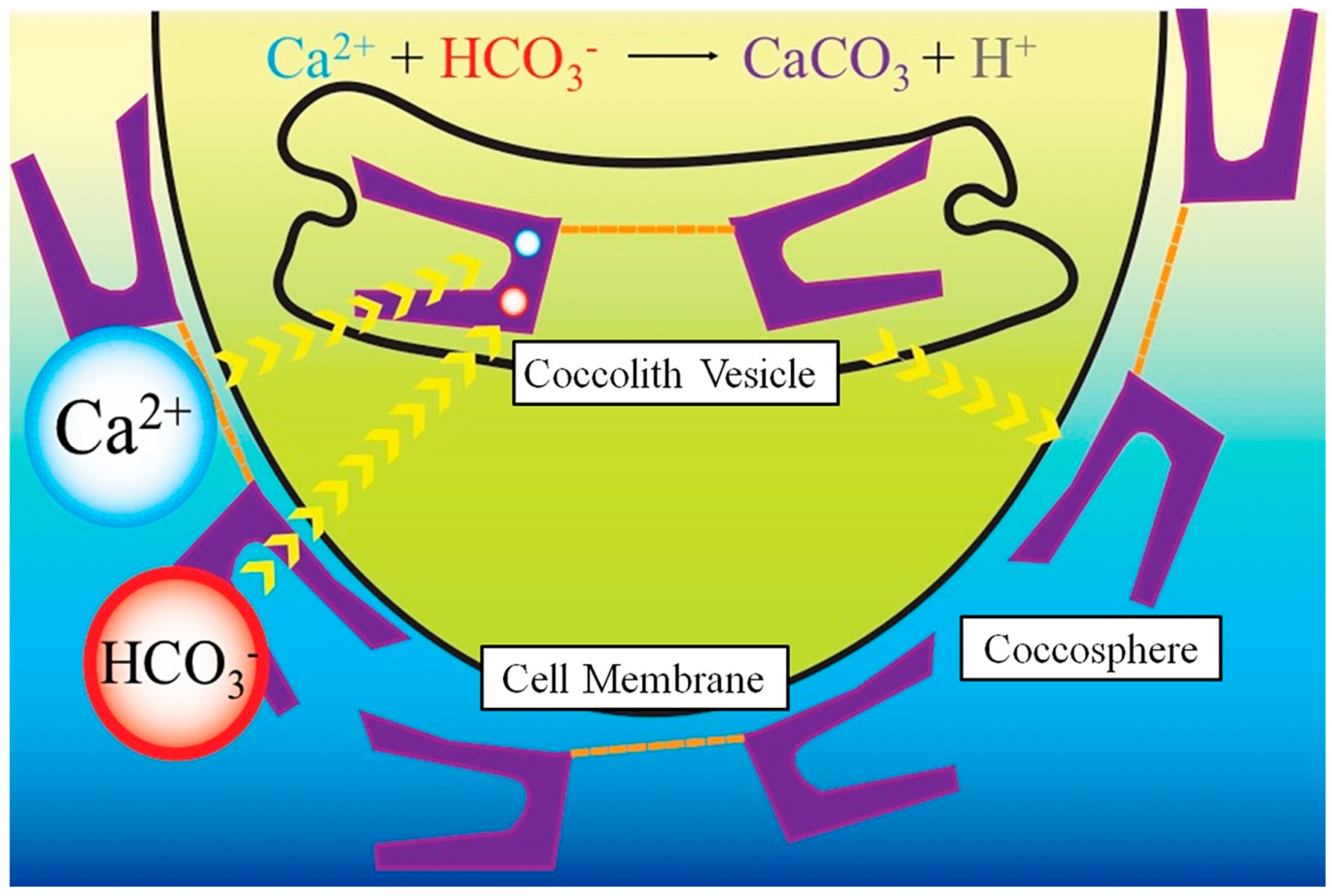
4. Coccolithophore Biomineralisation
5. Coccolithophore Source
6. Translations into Strategies for Bone Repair
7. Conclusions and Future Directions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bansal, V.; Bharde, A.; Ramanathan, R.; Bhargava, S.K. Inorganic materials using ‘unusual’ microorganisms. Adv. Colloid Interface Sci. 2012, 179, 150–168. [Google Scholar] [CrossRef] [PubMed]
- Gower, L.B. Biomimetic model systems for investigating the amorphous precursor pathway and its role in biomineralization. Chem. Rev. 2008, 108, 4551–4627. [Google Scholar] [CrossRef] [PubMed]
- Gal, A.; Weiner, S.; Addadi, L. A perspective on underlying crystal growth mechanisms in biomineralization: Solution mediated growth versus nanosphere particle accretion. CrystEngComm 2015, 17, 2606–2615. [Google Scholar] [CrossRef]
- Chen, P.-Y.; Lin, A.Y.M.; Lin, Y.-S.; Seki, Y.; Stokes, A.G.; Peyras, J.; Olevsky, E.A.; Meyers, M.A.; McKittrick, J. Structure and mechanical properties of selected biological materials. J. Mech. Behav. Biomed. Mater. 2008, 1, 208–226. [Google Scholar] [CrossRef] [PubMed]
- Studart, A.R. Towards high-performance bioinspired composites. Adv. Mater. 2012, 24, 5024–5044. [Google Scholar] [CrossRef] [PubMed]
- Kamat, S.; Su, X.; Ballarini, R.; Heuer, A.H. Structural basis for the fracture toughness of the shell of the conch Strombus gigas. Nature 2000, 405, 1036–1040. [Google Scholar] [CrossRef] [PubMed]
- Clarke, S.A.; Walsh, P.; Maggs, C.A.; Buchanan, F. Designs from the deep: Marine organisms for bone tissue engineering. Biotechnol. Adv. 2011, 29, 610–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, L.; Kolle, S.; Weaver, J.C.; Ortiz, C.; Aizenberg, J.; Kolle, M. A highly conspicuous mineralized composite photonic architecture in the translucent shell of the blue-rayed limpet. Nat. Commun. 2015, 6, 6322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.A.; Halfar, J. First evidence of chitin in calcified coralline algae: New insights into the calcification process of Clathromorphum compactum. Sci. Rep. 2014, 4, 6162. [Google Scholar] [CrossRef] [PubMed]
- Marsh, M.E. Regulation of CaCO3 formation in coccolithophores. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2003, 136, 743–754. [Google Scholar] [CrossRef]
- Drescher, B.; Dillaman, R.M.; Taylor, A.R. Coccolithogenesis in Scyphosphaera apsteinii (Prymnesiophyceae). J. Phycol. 2012, 48, 1343–1361. [Google Scholar] [CrossRef] [PubMed]
- Durak, G.M.; Taylor, A.R.; Probert, I.; de Vargas, C.; Audic, S.; Schroeder, D.C.; Brownlee, C.; Wheeler, G.L. A role for diatom-like silicon transporters in calcifying coccolithophores. Nat. Commun. 2016, 7, 10543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagasawa, H. The Molecular Mechanism of Calcification in Aquatic Organisms. Biosci. Biotechnol. Biochem. 2013, 77, 1991–1996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knight, M.J.; Senior, L.; Nancolas, B.; Ratcliffe, S.; Curnow, P. Direct evidence of the molecular basis for biological silicon transport. Nat. Commun. 2016, 7, 11926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Round, F.E.; Crawford, R.M.; Mann, D.G. Diatoms: Biology and Morphology of the Genera; Cambridge University Press: Cambridge, UK, 2007; pp. 334–339. ISBN 13 9780521714693. [Google Scholar]
- Gordon, R.; Losic, D.; Tiffany, M.A.; Nagy, S.S.; Sterrenburg, F.A.S. The glass menagerie: Diatoms for novel applications in nanotechnology. Trends Biotechnol. 2009, 27, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Young, J.R.; Geisen, M.; Cros, L.; Kleijne, A.; Sprengel, C.; Probert, I.; Østergaard, J. A guide to extant coccolithophore taxonomy. J. Nannoplankt. Res. 2003, 125. Available online: http://ina.tmsoc.org/JNR/online/SpecIssue/Young_et_al_2003_JNR%20Special_Issue_TaxonomyGuide.pdf (accessed on 20 August 2018).
- Lapidot, S.; Meirovitch, S.; Sharon, S.; Heyman, A.; Kaplan, D.L.; Shoseyov, O. Clues for biomimetics from natural composite materials. Nanomedicine 2012, 7, 1409–1423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finnemore, A.; Cunha, P.; Shean, T.; Vignolini, S.; Guldin, S.; Oyen, M.; Steiner, U. Biomimetic layer-by-layer assembly of artificial nacre. Nat. Commun. 2012, 3, 966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gehrke, N.; Nassif, N.; Pinna, N.; Antonietti, M.; Gupta, H.S.; Cölfen, H. Retrosynthesis of nacre via amorphous precursor particles. Chem. Mater. 2005, 17, 6514–6516. [Google Scholar] [CrossRef]
- Withers, N. Biominerals: Tomography reveals all. Nat. Chem. 2010, 2, 78. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.R.; Chrachri, A.; Wheeler, G.; Goddard, H.; Brownlee, C. A voltage-gated H+ channel underlying pH homeostasis in calcifying coccolithophores. PLoS Biol. 2011, 9. [Google Scholar] [CrossRef] [PubMed]
- Quinn, P.; Bowers, R.M.; Zhang, X.; Wahlund, T.M.; Fanelli, M.A.; Olszova, D.; Read, B.A. cDNA microarrays as a tool for identification of biomineralization proteins in the coccolithophorid Emiliania huxleyi (Haptophyta). Appl. Environ. Microbiol. 2006, 72, 5512–5526. [Google Scholar] [CrossRef] [PubMed]
- Brownlee, C.; Wheeler, G.; Taylor, A.R. Coccolithophore biomineralization: New questions, new answers. Semin. Cell Dev. Biol. 2015, 46, 11–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holtz, L.M.; Langer, G.; Rokitta, S.D.; Thoms, S. Synthesis of nanostructured calcite particles in coccolithophores, unicellular algae. In Green Biosynthesis of Nanoparticles: Mechanisms and Applications; Rai, M., Posten, C., Eds.; CABI: Oxfordshire, UK, 2013; p. 132. [Google Scholar]
- Gal, A.; Wirth, R.; Kopka, J.; Fratzl, P.; Faivre, D.; Scheffel, A. Macromolecular recognition directs calcium ions to coccolith mineralization sites. Science 2016, 353, 590–593. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Yao, Y.; Zou, X.; Fan, S.; Zhou, Q.; Dai, Q.; Dong, F.; Liu, M.; Nie, X.; Tan, D.; et al. Nano-scale spatial assessment of calcium distribution in coccolithophores using synchrotron-based nano-CT and STXM-NEXAFS. Int. J. Mol. Sci. 2014, 15, 23604–23615. [Google Scholar] [CrossRef] [PubMed]
- Young, J.R.; Henriksen, K. Biomineralization within Vesicles: The Calcite of Coccoliths. Rev. Mineral. Geochem. 2003, 189–215. [Google Scholar] [CrossRef]
- Taylor, A.R.; Russell, M.A.; Harper, G.M.; Collins, T.F.T.; Brownlee, C. Dynamics of formation and secretion of heterococcoliths by Coccolithus pelagicus ssp. braarudii. Eur. J. Phycol. 2007, 42, 125–136. [Google Scholar] [CrossRef]
- Henriksen, K.; Young, J.R.; Bown, P.R.; Stipp, S.L.S. Coccolith biomineralisation studied with atomic force microscopy. Palaeontology 2004, 47, 725–743. [Google Scholar] [CrossRef] [Green Version]
- Young, J.R.; Didymus, J.M.; Brown, P.R.; Prins, B.; Mann, S. Crystal assembly and phylogenetic evolution in heterococcoliths. Nature 1992, 356, 516–518. [Google Scholar] [CrossRef]
- Young, J.; Davis, S.; Bown, P.; Mann, S. Coccolith ultrastructure and biomineralisation. J. Struct. Biol. 1999, 126, 195–215. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, B.; Bowers, R.M.; Wahlund, T.M.; Read, B.A. Suppressive subtractive hybridization of and differences in gene expression content of calcifying and noncalcifying cultures of Emiliania huxleyi strain 1516. Appl. Environ. Microbiol. 2005, 71, 2564–2575. [Google Scholar] [CrossRef] [PubMed]
- Aloisi, G. Covariation of metabolic rates and cell size in coccolithophores. Biogeosciences 2015, 4665–4692. [Google Scholar] [CrossRef] [Green Version]
- Von Dassow, P.; Ogata, H.; Probert, I.; Wincker, P.; Da Silva, C.; Audic, S.; Claverie, J.-M.; de Vargas, C. Transcriptome analysis of functional differentiation between haploid and diploid cells of Emiliania huxleyi, a globally significant photosynthetic calcifying cell. Genome Biol. 2009, 10, R114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richier, S.; Fiorini, S.; Kerros, M.-E.; von Dassow, P.; Gattuso, J.-P. Response of the calcifying coccolithophore Emiliania huxleyi to low pH/high pCO(2): From physiology to molecular level. Mar. Biol. 2011, 158, 551–560. [Google Scholar] [CrossRef] [PubMed]
- Langer, G.; Probert, I.; Nehrke, G.; Ziveri, P. The morphological response of Emiliania huxleyi to seawater carbonate chemistry changes: An inter-strain comparison. J. Nannoplankt. Res. 2011, 32, 29–34. [Google Scholar]
- Langer, G.; De Nooijer, L.J.; Oetjen, K. On the role of the cytoskeleton in coccolith morphogenesis: The effect of cytoskeleton inhibitors. J. Phycol. 2010, 46, 1252–1256. [Google Scholar] [CrossRef] [Green Version]
- Gibbs, S.J.; Poulton, A.J.; Bown, P.R.; Daniels, C.J.; Hopkins, J.; Young, J.R.; Jones, H.L.; Thiemann, G.J.; O’Dea, S.A.; Newsam, C. Species-specific growth response of coccolithophores to Palaeocene–Eocene environmental change. Nat. Geosci. 2013, 6, 218–222. [Google Scholar] [CrossRef]
- Aubry, M. A major Pliocene coccolithophore turnover: Change in morphological strategy in the photic zone. Geol. Soc. Am. Spec. Pap. 2007, 424, 25–51. [Google Scholar]
- Van De Locht, R.; Slater, T.J.A.; Verch, A.; Young, J.R.; Haigh, S.J.; Kröger, R. Ultrastructure and crystallography of nanoscale calcite building blocks in Rhabdosphaera clavigera coccolith spines. Cryst. Growth Des. 2014, 14, 1710–1718. [Google Scholar] [CrossRef]
- Fritz, J.J. Carbon fixation and coccolith detachment in the coccolithophore Emiliania huxleyi in nitrate-limited cyclostats. Mar. Biol. 1999, 133, 509–518. [Google Scholar] [CrossRef]
- Luby-Phelps, K. The physical chemistry of cytoplasm and its influence on cell function: An update. Mol. Biol. Cell 2013, 24, 2593–2596. [Google Scholar] [CrossRef] [PubMed]
- Godoi, R.H.M.; Aerts, K.; Harlay, J.; Kaegi, R.; Ro, C.; Chou, L.; Van Grieken, R. Organic surface coating on Coccolithophores-Emiliania huxleyi: Its determination and implication in the marine carbon cycle. Microchem. J. 2009, 91, 266–271. [Google Scholar] [CrossRef]
- Yang, M.; Stipp, S.L.S.; Harding, J. Biological control on calcite crystallization by polysaccharides. Cryst. Growth Des. 2008, 8, 4066–4074. [Google Scholar] [CrossRef]
- Hamm, L.M.; Giuffre, A.J.; Han, N.; Tao, J.; Wang, D.; De Yoreo, J.J.; Dove, P.M. Reconciling disparate views of template-directed nucleation through measurement of calcite nucleation kinetics and binding energies. Proc. Natl. Acad. Sci. USA 2014, 111, 1304–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giuffre, A.J.; Hamm, L.M.; Han, N.; De Yoreo, J.J.; Dove, P.M. Polysaccharide chemistry regulates kinetics of calcite nucleation through competition of interfacial energies. Proc. Natl. Acad. Sci. USA 2013, 110, 9261–9266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raue, L.; Klein, H.; Raabe, D. The Exoskeleton of the American Lobster—From Texture to Anisotropic Properties. Solid State Phenom. 2010, 160, 287–294. [Google Scholar] [CrossRef]
- Marsh, M.E. Biomineralization: Progress in Biology, Molecular Biology and Application, 2nd ed.; Completely Revised and Extended Edition; Edmund, B., Ed.; Wiley-VCH: Weinheim, Germany, 2016; p. 361. ISBN 978-3-527-60461-6. [Google Scholar]
- Marsh, M.E.; Chang, D.K.; King, G.C. Isolation and characterization of a novel acidic polysaccharide containing tartrate and glyoxylate residues from the mineralized scales of a unicellular coccolithophorid alga Pleurochrysis carterae. J. Biol. Chem. 1992, 267, 20507–20512. [Google Scholar] [PubMed]
- Hirokawa, Y. Localization and Associative Strength of Acid Polysaccharides in Coccoliths of Pleurochrysis haptonemofera (Haptophyta) Predicted from Their Extractability from Partially Decalcified Coccoliths. Open J. Mar. Sci. 2013, 3, 48–54. [Google Scholar] [CrossRef]
- Verret, F.; Wheeler, G.; Taylor, A.R.; Farnham, G.; Brownlee, C. Calcium channels in photosynthetic eukaryotes: Implications for evolution of calcium-based signalling. New Phytol. 2010, 187, 23–43. [Google Scholar] [CrossRef] [PubMed]
- Lohbeck, K.T.; Riebesell, U.; Reusch, T.B.H. Gene expression changes in the coccolithophore Emiliania huxleyi after 500 generations of selection to ocean acidification. Proc. R. Soc. B 2014, 281, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakurada, S.; Fujiwara, S.; Suzuki, M.; Kogure, T.; Uchida, T.; Umemura, T.; Tsuzuki, M. Involvement of acidic polysaccharide Ph-PS-2 and protein in initiation of Coccolith mineralization, as demonstrated by in vitro calcification on the base plate. Mar. Biotechnol. 2018, 20, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Kegel, J.U.; John, U.; Valentin, K.; Frickenhaus, S. Genome Variations Associated with Viral Susceptibility and Calcification in Emiliania huxleyi. PLoS ONE 2013, 8, e80684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, A.R.; Brownlee, C.; Wheeler, G. Coccolithophore cell biology: Chalking up progress. Ann. Rev. Mar. Sci. 2016, 9, 18.1–18.28. [Google Scholar] [CrossRef] [PubMed]
- Kocot, K.M.; Aguilera, F.; McDougall, C.; Jackson, D.J.; Degnan, B.M. Sea shell diversity and rapidly evolving secretomes: Insights into the evolution of biomineralization. Front. Zool. 2016, 13, 23. [Google Scholar] [CrossRef] [PubMed]
- Kayano, K.; Saruwatari, K.; Kogure, T.; Shiraiwa, Y. Effect of coccolith polysaccharides isolated from the coccolithophorid, Emiliania huxleyi, on calcite crystal formation in in vitro CaCO3 crystallization. Mar. Biotechnol. 2011, 13, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Borman, A.H.; Jong, E.W.; Huizinga, M.; Kok, D.J.; Westbroek, P.; Bosch, L. The Role in CaCO3 Crystallization of an Acid Ca2+-Binding Polysaccharide Associated with Coccoliths of Emiliania huxleyi. Eur. J. Biochem. 1982, 129, 179–183. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, K.; Stipp, S.L.S.; Young, J.R.; Marsh, M.E. Biological control on calcite crystallization: AFM investigation of coccolith polysaccharide function. Am. Mineral. 2004, 89, 1709–1716. [Google Scholar] [CrossRef]
- Rodriguez, I.; Phytologist, N.; Rubisco, T.; Hemisphere, S.; Co, T. Commentary Calcification and ocean acidification: New insights from the coccolithophore Emiliania huxleyi. New Phytol. 2013, 2011–2013. [Google Scholar] [CrossRef]
- Read, B.A.; Kegel, J.; Klute, M.J.; Kuo, A.; Lefebvre, S.C.; Maumus, F.; Mayer, C.; Miller, J.; Monier, A.; Salamov, A.; et al. Pan genome of the phytoplankton Emiliania underpins its global distribution. Nature 2013, 499, 209–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sviben, S.; Gal, A.; Hood, M.A.; Bertinetti, L.; Politi, Y.; Bennet, M.; Krishnamoorthy, P.; Schertel, A.; Wirth, R.; Sorrentino, A.; et al. A vacuole-like compartment concentrates a disordered calcium phase in a key coccolithophorid alga. Nat. Commun. 2016, 7, 11228. [Google Scholar] [CrossRef] [PubMed]
- Gal, A.; Sviben, S.; Wirth, R.; Schreiber, A.; Lassalle-Kaiser, B.; Faivre, D.; Scheffel, A. Trace-element incorporation into intracellular pools uncovers calcium-pathways in a coccolithophore. Adv. Sci. 2017, 4. [Google Scholar] [CrossRef] [PubMed]
- Rosario Lorenzo, M.; Iñiguez, C.; Egge, J.K.; Larsen, A.; Berger, S.A.; García-Gómez, C.; Segovia, M. Increased CO2 and iron availability effects on carbon assimilation and calcification on the formation of Emiliania huxleyi blooms in a coastal phytoplankton community. Environ. Exp. Bot. 2018, 148, 47–58. [Google Scholar] [CrossRef]
- Andersson, M.; Chen, G.; Otikovs, M.; Landreh, M.; Nordling, K.; Kronqvist, N.; Westermark, P.; Jörnvall, H.; Knight, S.; Ridderstråle, Y.; et al. Carbonic anhydrase generates CO2 and H+ that drive spider silk formation via opposite effects on the terminal domains. PLoS Biol. 2014, 12, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Durak, G.M.; Brownlee, C.; Wheeler, G.L. The role of the cytoskeleton in biomineralisation in haptophyte algae. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Beaufort, L.; Probert, I.; de Garidel-Thoron, T.; Bendif, E.M.; Ruiz-Pino, D.; Metzl, N.; Goyet, C.; Buchet, N.; Coupel, P.; Grelaud, M.; et al. Sensitivity of coccolithophores to carbonate chemistry and ocean acidification. Nature 2011, 476, 80–83. [Google Scholar] [CrossRef] [PubMed]
- Bach, L.T.; MacKinder, L.C.M.; Schulz, K.G.; Wheeler, G.; Schroeder, D.C.; Brownlee, C.; Riebesell, U. Dissecting the impact of CO2 and pH on the mechanisms of photosynthesis and calcification in the coccolithophore Emiliania huxleyi. New Phytol. 2013, 199, 121–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Young, J.R. Morphology of Emilliania huxleyi coccoliths on the North West European shelf—Is there an influence of carbonate chemistry? Biogeosciences 2014, 11, 4531–4561. [Google Scholar] [CrossRef]
- Borchard, C.; Borges, A.V.; Händel, N.; Engel, A. Biogeochemical response of Emiliania huxleyi (PML B92/11) to elevated CO2 and temperature under phosphorous limitation: A chemostat study. J. Exp. Mar. Biol. Ecol. 2011, 410, 61–71. [Google Scholar] [CrossRef]
- Thierstein, H.R.; Young, J.R. Coccolithophores: From Molecular Processes to Global Impact; Springer: New York, NY, USA, 2004; pp. 95–125. ISBN 978-3-662-06278-4. [Google Scholar]
- Chow, J.S.; Lee, C.; Engel, A. The influence of extracellular polysaccharides, growth rate, and free coccoliths on the coagulation efficiency of Emiliania huxleyi. Mar. Chem. 2015, 175, 5–17. [Google Scholar] [CrossRef]
- Engel, A.; Szlosek, J.; Abramson, L.; Liu, Z.; Lee, C. Investigating the effect of ballasting by CaCO3 in Emiliania huxleyi: I. Formation, settling velocities and physical properties of aggregates. Deep. Res. Part II Top. Stud. Oceanogr. 2009, 56, 1396–1407. [Google Scholar] [CrossRef]
- Mizukawa, Y.; Miyashita, Y.; Satoh, M.; Shiraiwa, Y.; Iwasaka, M. Light intensity modulation by coccoliths of Emiliania huxleyi as a micro-photo-regulator. Sci. Rep. 2015, 5, 13577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.D.; Amirthalingam, S.; Kim, S.L.; Lee, S.S.; Rangasamy, J.; Hwang, N.S. Biomimetic Materials and Fabrication Approaches for Bone Tissue Engineering. Adv. Healthc. Mater. 2017. [Google Scholar] [CrossRef] [PubMed]
- Sailaja, G.S.; Ramesh, P.; Vellappally, S.; Anil, S.; Varma, H.K. Biomimetic approaches with smart interfaces for bone regeneration. J. Biomed. Sci. 2016, 23, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hasan, A.; Byambaa, B.; Morshed, M.; Cheikh, M.I.; Shakoor, R.A.; Mustafy, T.; Marei, H.E. Advances in osteobiologic materials for bone substitutes. J. Tissue Eng. Regen. Med. 2018, 12, 1448–1468. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, N.; Moratti, S.C.; Dias, G.J. Hydroxyapatite–polymer biocomposites for bone regeneration: A review of current trends. J. Biomed. Mater. Res. Part B Appl. Biomater. 2018, 106, 2046–2057. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Yeung, K.W.K. Bone grafts and biomaterials substitutes for bone defect repair: A review. Bioact. Mater. 2017. [Google Scholar] [CrossRef] [PubMed]
- Kolk, A.; Handschel, J.; Drescher, W.; Rothamel, D.; Kloss, F.; Blessmann, M.; Heiland, M.; Wolff, K.D.; Smeets, R. Current trends and future perspectives of bone substitute materials—From space holders to innovative biomaterials. J. Cranio-Maxillofac. Surg. 2012, 40, 706–718. [Google Scholar] [CrossRef] [PubMed]
- Ricciardi, B.F.; Bostrom, M.P. Bone graft substitutes: Claims and credibility. Semin. Arthroplasty 2013, 24, 119–123. [Google Scholar] [CrossRef]
- Khoury, F.; Keller, P.; Keeve, P. Stability of Grafted Implant Placement Sites After Sinus Floor Elevation Using a Layering Technique: 10-Year Clinical and Radiographic Results. Int. J. Oral Maxillofac. Implants 2017, 32, 1086–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fee, K.; Walker, G.M.; Dunne, N.J.; Buchanan, F.J. Hydrothermal synthesis of coccolith rich chalk to hydroxyapatite. Int. J. Nano Biomater. 2012, 4, 81. [Google Scholar] [CrossRef]
- Lord, M.S.; Foss, M.; Besenbacher, F. Influence of nanoscale surface topography on protein adsorption and cellular response. Nano Today 2010, 5, 66–78. [Google Scholar] [CrossRef]
- Yang, L.; Liu, H.; Lin, Y. Biomaterial nanotopography-mediated cell responses: Experiment and modeling. Int. J. Smart Nano Mater. 2015, 5, 227–256. [Google Scholar] [CrossRef]
- Jain, G.; Pendola, M.; Huang, Y.C.; Gebauer, D.; Evans, J.S. A Model Sea Urchin Spicule Matrix Protein, rSpSM50, Is a Hydrogelator That Modifies and Organizes the Mineralization Process. Biochemistry 2017, 56, 2663–2675. [Google Scholar] [CrossRef] [PubMed]
- So, C.R.; Liu, J.; Fears, K.P.; Leary, D.H.; Golden, J.P.; Wahl, K.J. Self-Assembly of Protein Nanofibrils Orchestrates Calcite Step Movement through Selective Nonchiral Interactions. ACS Nano 2015, 9, 5782–5791. [Google Scholar] [CrossRef] [PubMed]
- Jackson, E.; Ferrari, M.; Cuestas-Ayllon, C.; Fernández-Pacheco, R.; Perez-Carvajal, J.; De La Fuente, J.M.; Grazú, V.; Betancor, L. Protein-templated biomimetic silica nanoparticles. Langmuir 2015, 31, 3687–3695. [Google Scholar] [CrossRef] [PubMed]
- Foo, C.W.P.; Huang, J.; Kaplan, D.L. Lessons from seashells: Silica mineralization via protein templating. Trends Biotechnol. 2004, 22, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Yang, S.; Li, C.; Miao, Y.; Zhu, L.; Mao, C.; Yang, M. Nucleation and Assembly of Silica into Protein-Based Nanocomposites as Effective Anticancer Drug Carriers Using Self-Assembled Silk Protein Nanostructures as Biotemplates. ACS Appl. Mater. Interfaces 2017, 9, 22259–22267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnell, O.; Kanis, J.A. An estimate of the worldwide prevalence and disability associated with osteoporotic fractures. Osteoporos. Int. 2006, 17, 1726–1733. [Google Scholar] [CrossRef] [PubMed]



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walsh, P.J.; Fee, K.; Clarke, S.A.; Julius, M.L.; Buchanan, F.J. Blueprints for the Next Generation of Bioinspired and Biomimetic Mineralised Composites for Bone Regeneration. Mar. Drugs 2018, 16, 288. https://doi.org/10.3390/md16080288
Walsh PJ, Fee K, Clarke SA, Julius ML, Buchanan FJ. Blueprints for the Next Generation of Bioinspired and Biomimetic Mineralised Composites for Bone Regeneration. Marine Drugs. 2018; 16(8):288. https://doi.org/10.3390/md16080288
Chicago/Turabian StyleWalsh, Pamela J., Kathryn Fee, Susan A. Clarke, Matthew L. Julius, and Fraser J. Buchanan. 2018. "Blueprints for the Next Generation of Bioinspired and Biomimetic Mineralised Composites for Bone Regeneration" Marine Drugs 16, no. 8: 288. https://doi.org/10.3390/md16080288
APA StyleWalsh, P. J., Fee, K., Clarke, S. A., Julius, M. L., & Buchanan, F. J. (2018). Blueprints for the Next Generation of Bioinspired and Biomimetic Mineralised Composites for Bone Regeneration. Marine Drugs, 16(8), 288. https://doi.org/10.3390/md16080288