Tetracenomycin X Exerts Antitumour Activity in Lung Cancer Cells through the Downregulation of Cyclin D1


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
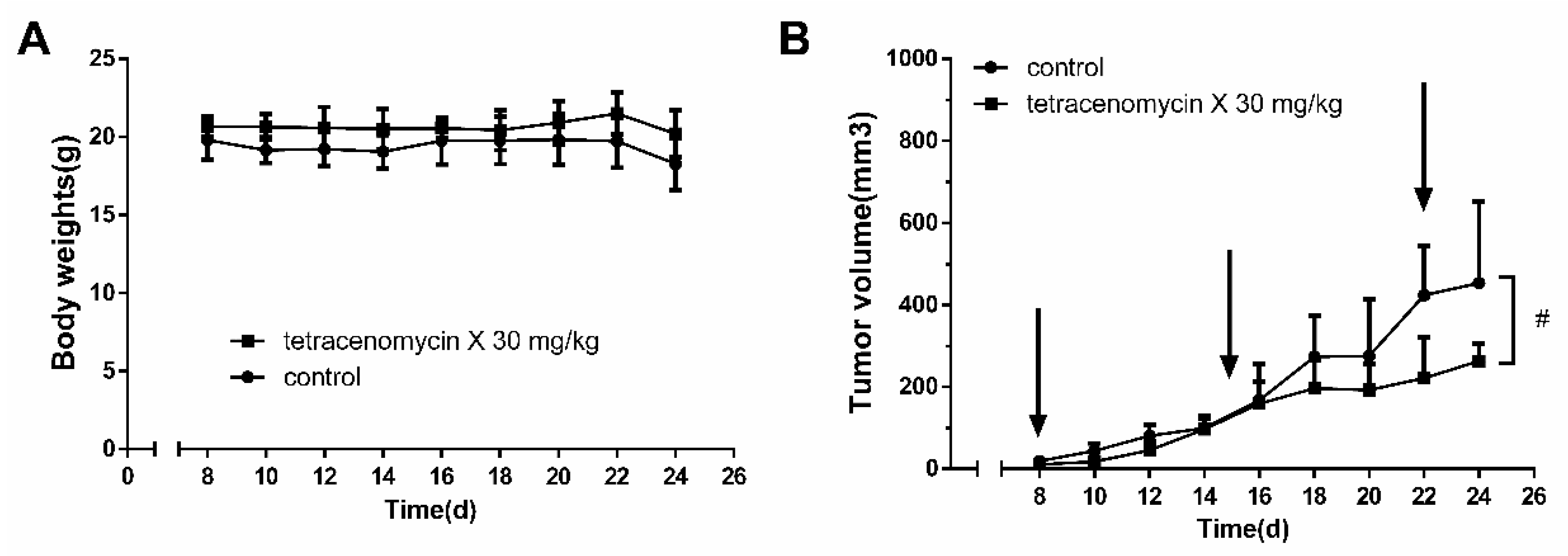
2.1. Tetracenomycin X Exerts Antitumour Activity in H460 Xenografts in BALB/c Nude Mice
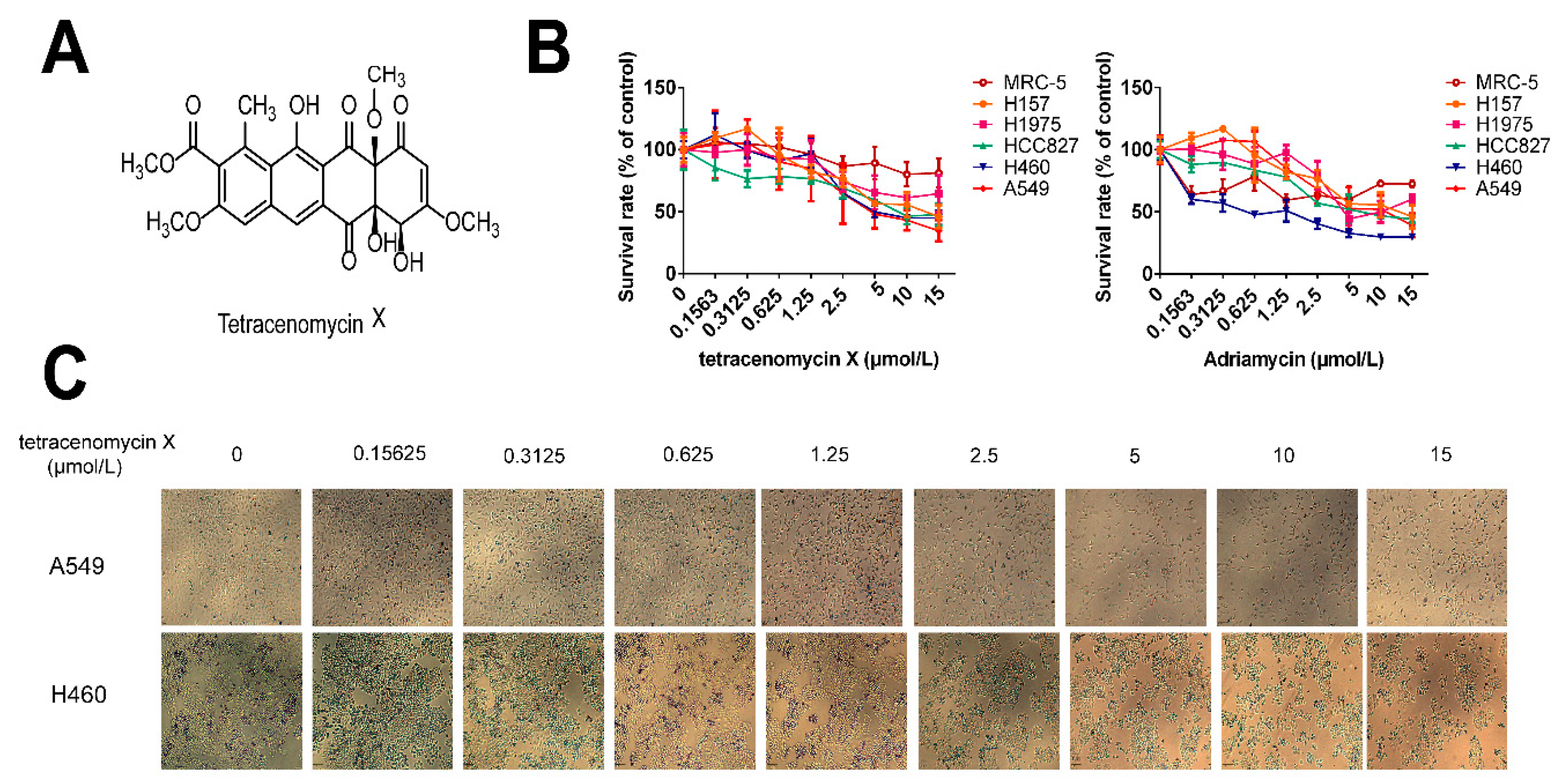
2.2. Tetracenomycin X Selectively Inhibits Human Lung Cancer Cell Proliferation
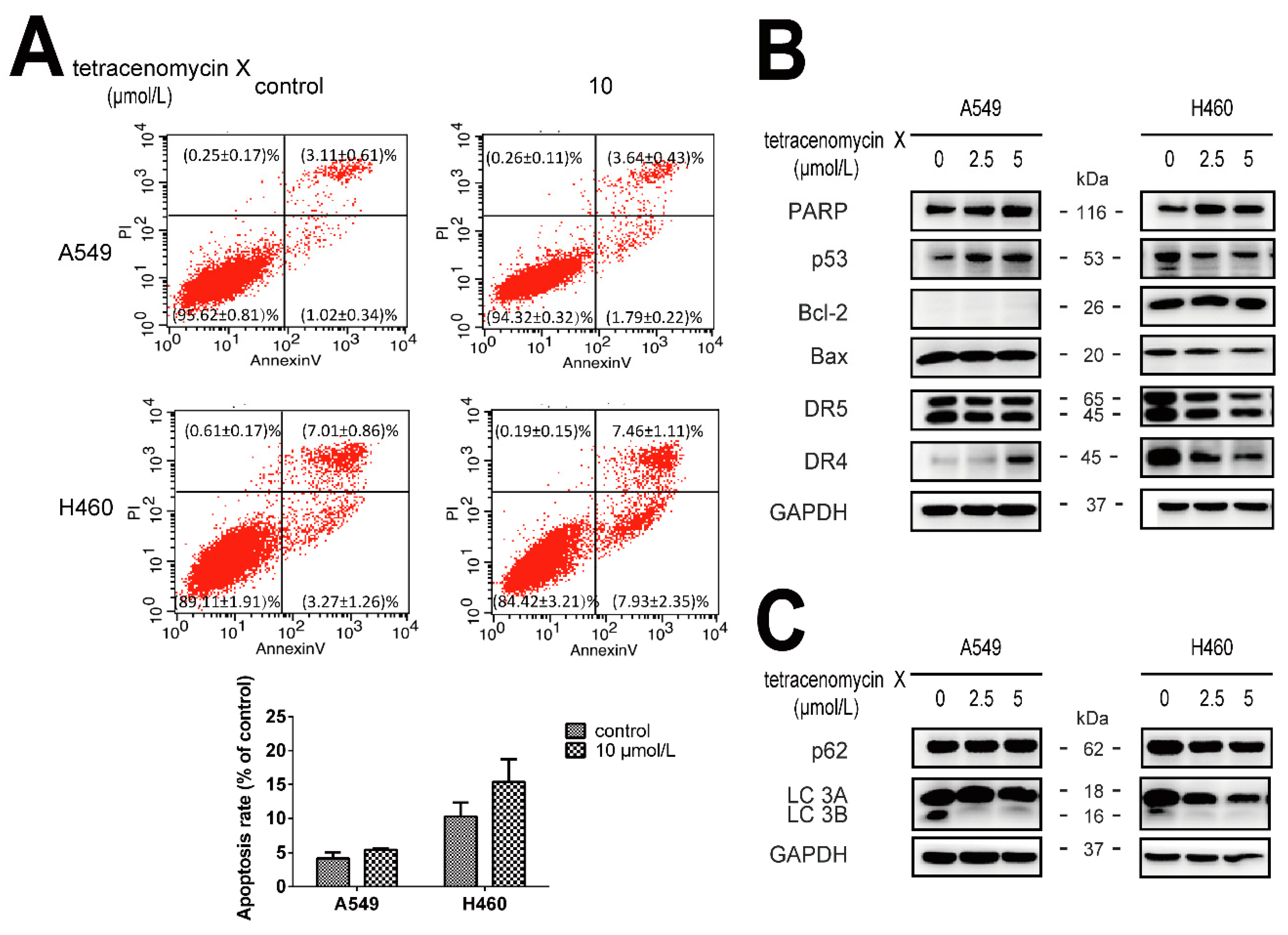
2.3. The Antitumour Activity of Tetracenomycin X Is Independent of Apoptosis and Autophagy
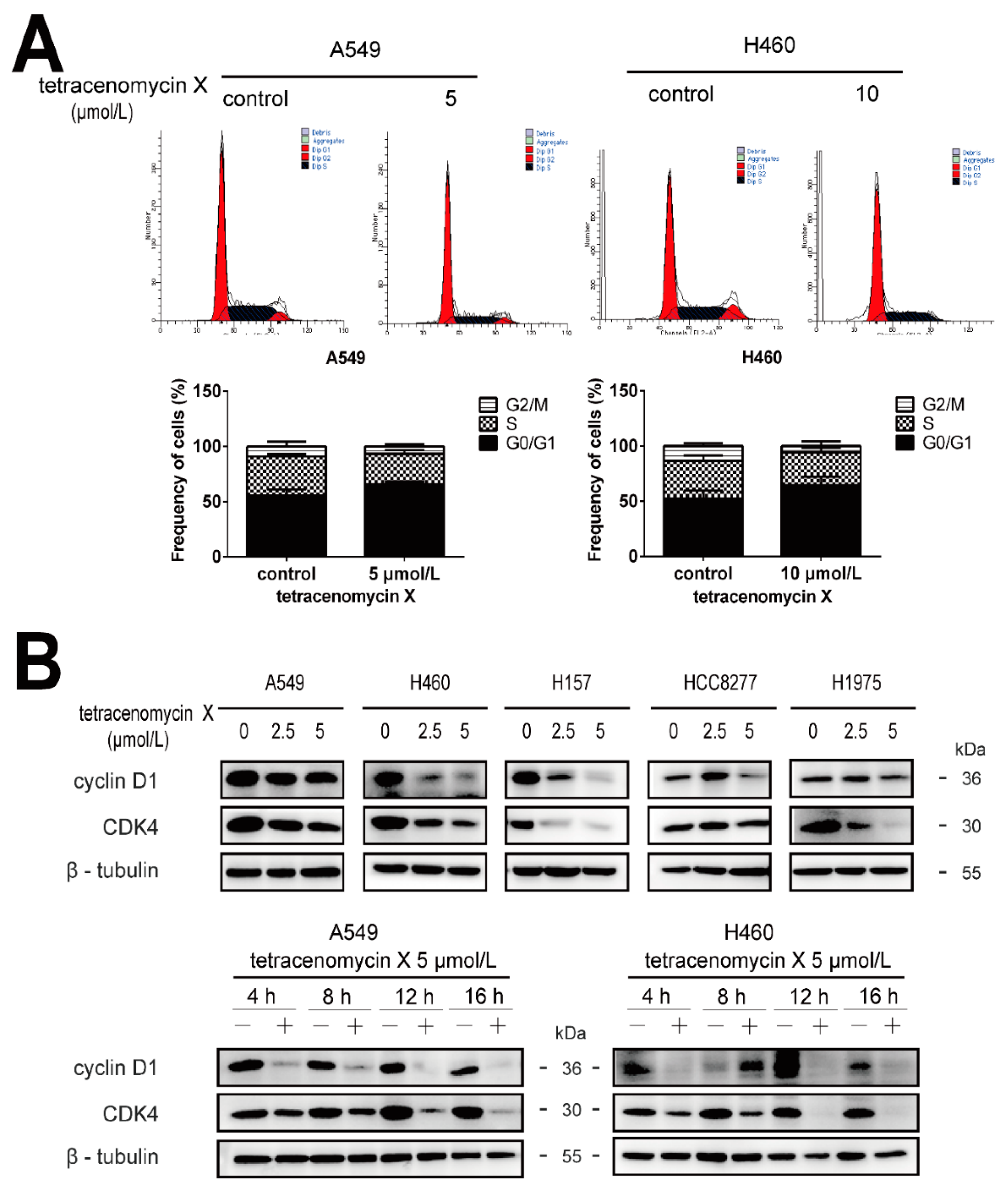
2.4. Tetracenomycin X Induces Cell Cycle Arrest in the G0/G1 Phase and Decreases the Expression Levels of Cell Cycle-Related Proteins in Lung Cancer Cells
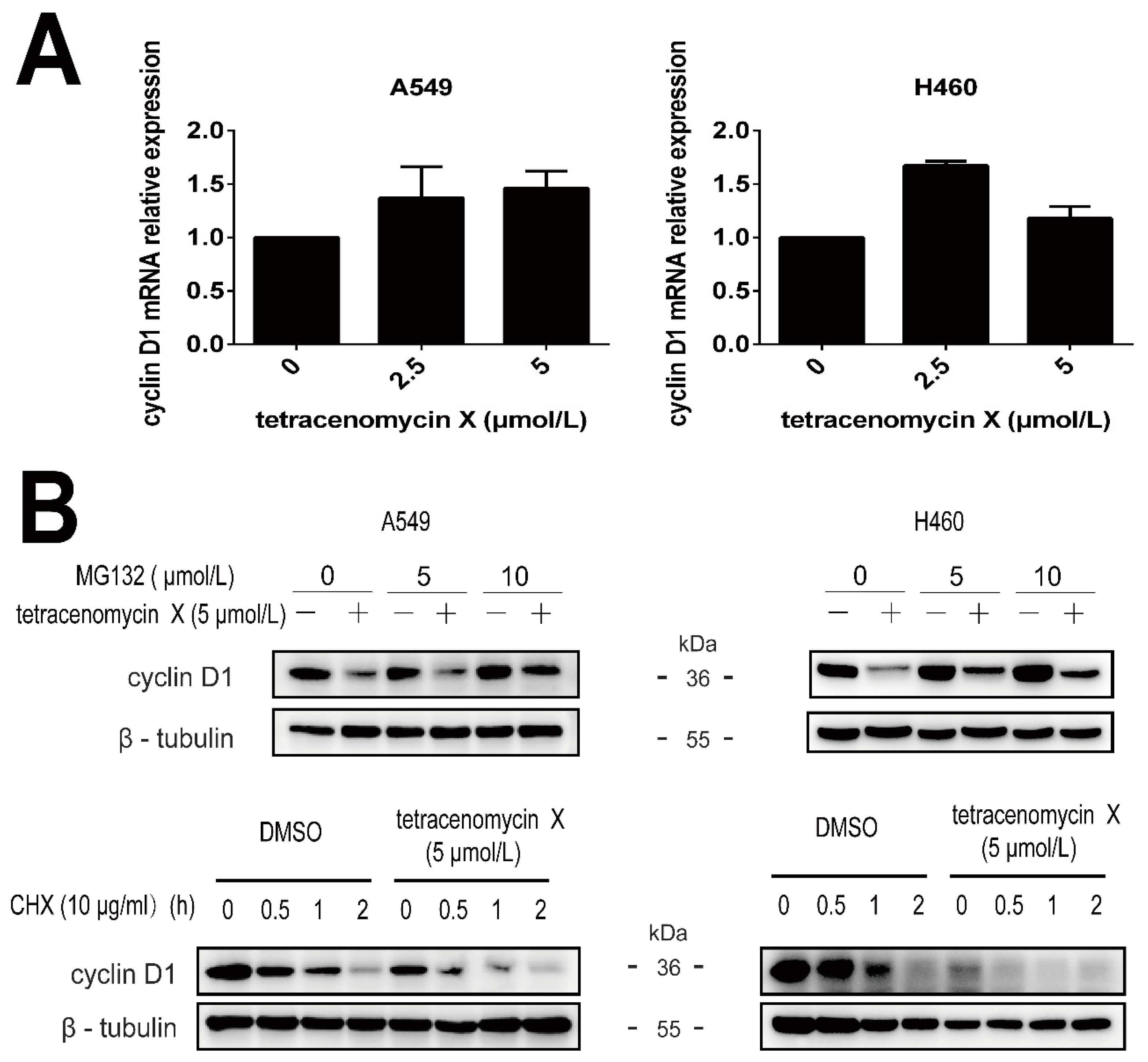
2.5. Tetracenomycin X Induces the Proteasomal Degradation of Cyclin D1
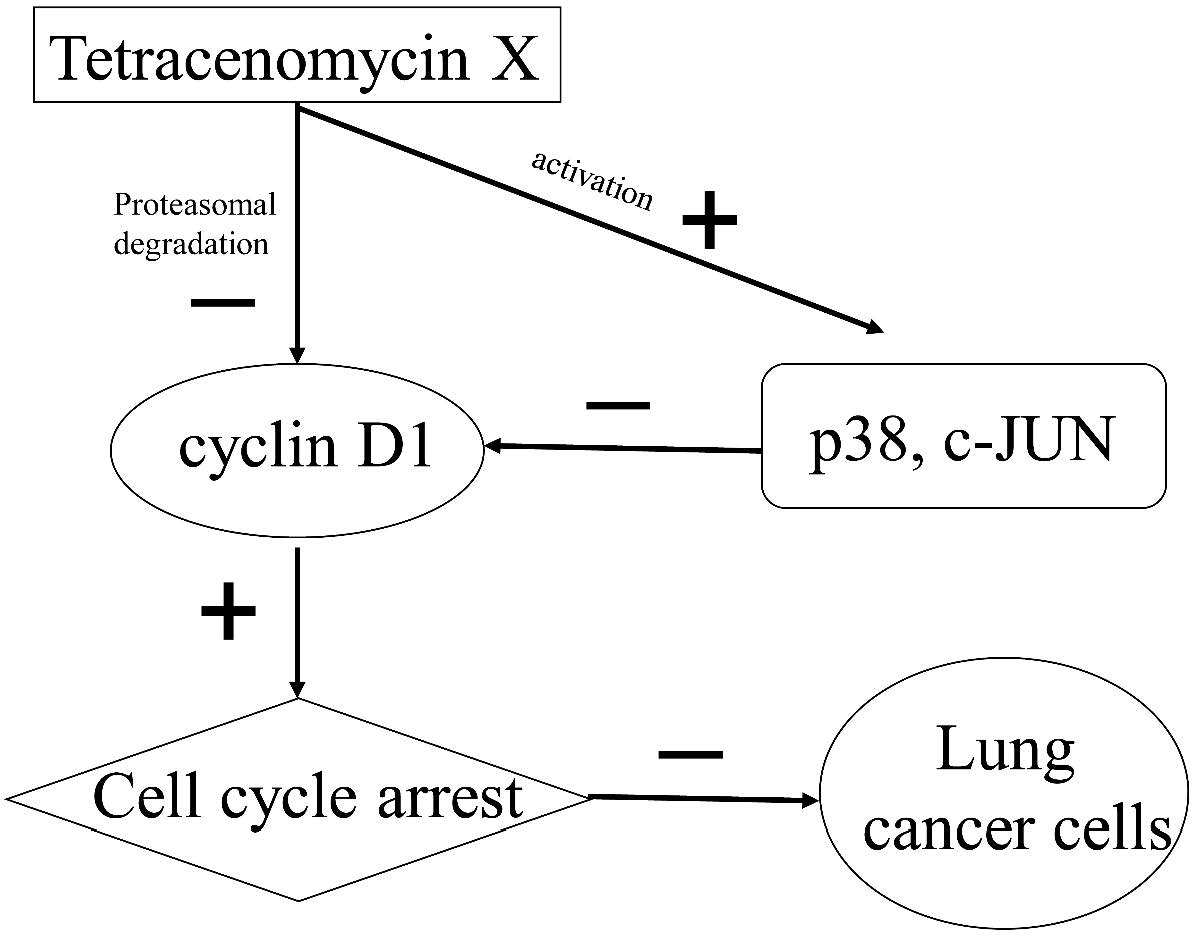
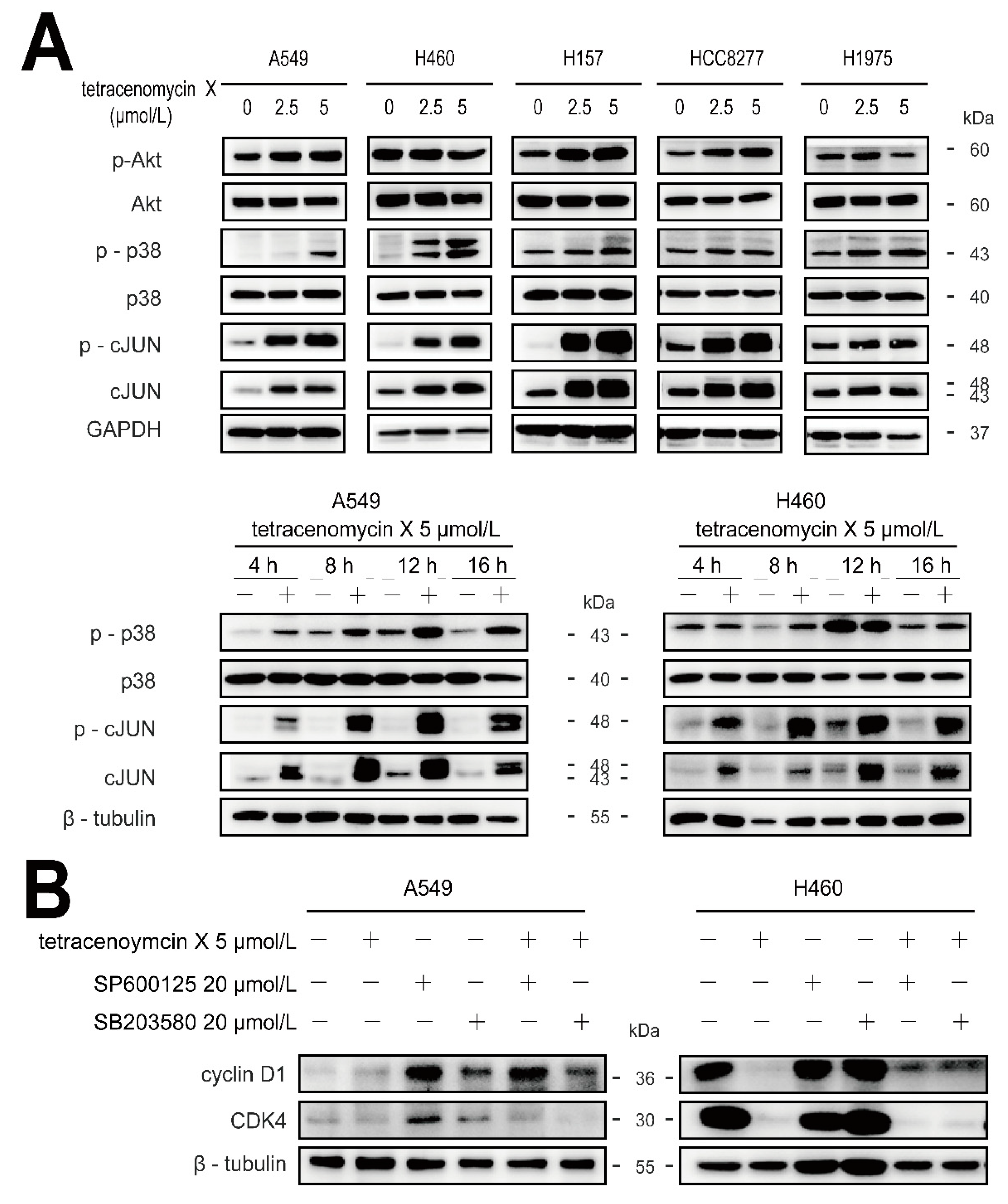
2.6. Tetracenomycin X Decreases the Expression of Cyclin D1 by the Activation of p38 and c-JUN
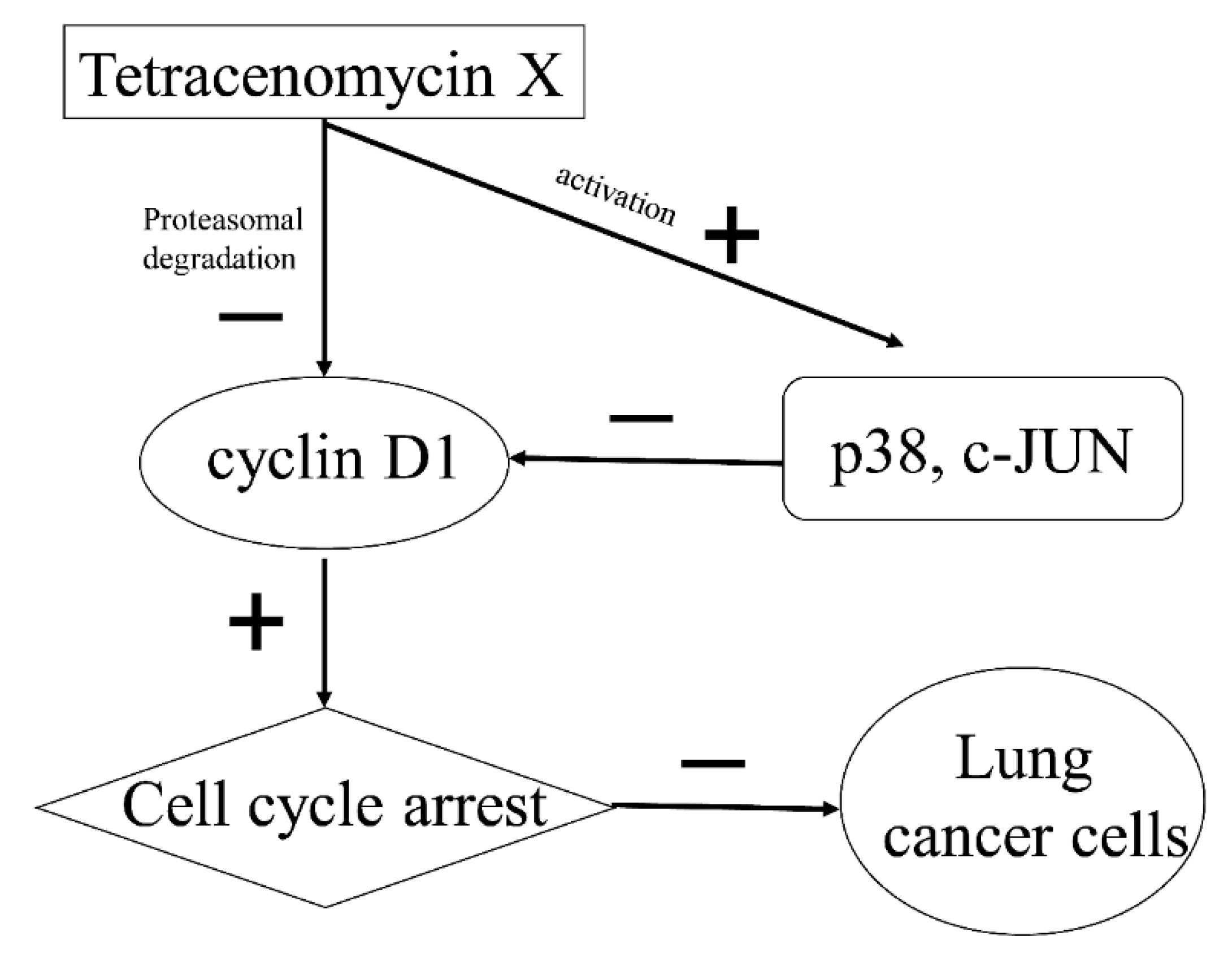
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Reagents and Antibodies
4.3. Cell Survival Assay
4.4. Western Blot Analysis
4.5. Annexin V-FITC/PI Staining
4.6. Cell Cycle Analysis
4.7. Reverse Transcriptase-Polymerase Chain Reaction
4.8. Lung Cancer Xenograft
4.9. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. CA A Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, A.; Ganti, A.K. Lung cancer-A global perspective. J. Surg. Oncol. 2017, 115, 550–554. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Newman, T.G.; Aronow, W.S. Lung cancer screening: History, current perspectives, and future directions. Arch. Med. Sci. AMS 2015, 11, 1033–1043. [Google Scholar] [PubMed]
- Jiang, J.; Huang, J.; Wang, X.R.; Quan, Y.H. MicroRNA-202 induces cell cycle arrest and apoptosis in lung cancer cells through targeting cyclin D1. Eur. Rev. Med. Pharm. Sci. 2016, 20, 2278–2284. [Google Scholar]
- Lazar, G.; Zähner, H.; Breiding, S.; Damberg, M.; Zeeck, A. 3-Demethoxy-3-ethoxy-tetracenomycin C. J. Antibiot. 1981, 34, 1067–1068. [Google Scholar] [CrossRef] [PubMed]
- Egert, E.; Noltemeyer, M.; Siebers, J.; Rohr, J.; Zeeck, A. The structure of tetracenomycin C. J. Antibiot. 1992, 45, 1190–1192. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Li, J.; Chen, M.; Hao, X.; Cao, F.; Tan, Y.; Ping, Y.; Wang, Y.; Xiao, C.; Gan, M. Seco-Tetracenomycins from the Marine-Derived Actinomycete Saccharothrix sp. 10-10. Mar. Drugs 2018, 16, 245. [Google Scholar] [CrossRef] [PubMed]
- Gan, M.; Liu, B.; Tan, Y.; Wang, Q.; Zhou, H.; He, H.; Ping, Y.; Yang, Z.; Wang, Y.; Xiao, C. Saccharothrixones A-D, Tetracenomycin-Type Polyketides from the Marine-Derived Actinomycete Saccharothrix sp. 10-10. J. Nat. Prod. 2015, 78, 2260–2265. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Tan, Y.; Gan, M.L.; Zhou, H.X.; Wang, Y.G.; Ping, Y.H.; Li, B.; Yang, Z.Y.; Xiao, C.L. [Identification of tetracenomycin X from a marine-derived Saccharothrix sp. guided by genes sequence analysis]. Yao Xue Xue Bao 2014, 49, 230–236. [Google Scholar] [PubMed]
- Weber, W.; Zähner, H.; Damberg, M.; Siebers, J.; Schroder, K.; Zeeck, A. Tetracenomycins—New antibiotics from Streptomyces glaucescens. In Proceedings of the International Symposium on Actinomycete Biology, Cologne, Germany, 3–7 September 1979. [Google Scholar]
- Wang, Y.; Gao, W.; Svitkin, Y.V.; Chen, A.P.; Cheng, Y.C. DCB-3503, a Tylophorine Analog, Inhibits Protein Synthesis through a Novel Mechanism. PLoS ONE 2010, 5, e11607. [Google Scholar] [CrossRef] [PubMed]
- Park, S.B.; Park, G.H.; Song, H.M.; Son, H.J.; Um, Y.; Kim, H.S.; Jeong, J.B. Anticancer activity of calyx of Diospyros kaki Thunb. through downregulation of cyclin D1 via inducing proteasomal degradation and transcriptional inhibition in human colorectal cancer cells. BMC Complement. Altern. Med. 2017, 17, 445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, M.K.; Park, G.H.; Eo, H.J.; Song, H.M.; Lee, J.W.; Kwon, M.J.; Koo, J.S.; Jeong, J.B. Tanshinone I induces cyclin D1 proteasomal degradation in an ERK1/2 dependent way in human colorectal cancer cells. Fitoterapia 2015, 101, 162–168. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Zhao, X.; Mo, Z.; Huang, W.; Yan, H.; Ling, Z.; Ye, Y. Formononetin promotes cell cycle arrest via downregulation of Akt/Cyclin D1/CDK4 in human prostate cancer cells. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2014, 34, 1351–1358. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.G.; Khoo, C.L.; Rickards, R.W. Oxidation processes in the biosynthesis of the tetracenomycin and elloramycin antibiotics. J. Antibiot. 1989, 42, 640–643. [Google Scholar] [CrossRef] [PubMed]
- Hutchinson, C.R. Biosynthetic Studies of Daunorubicin and Tetracenomycin C. Chem. Rev. 1997, 97, 2525–2536. [Google Scholar] [CrossRef] [PubMed]
- Vermeulen, K.; Bockstaele, D.R.V.; Berneman, Z.N. The cell cycle: A review of regulation, deregulation and therapeutic targets in cancer. Cell Prolif. 2010, 36, 131–149. [Google Scholar] [CrossRef]
- Mukhopadhyay, A.; Banerjee, S.; Stafford, L.J.; Xia, C.; Liu, M.; Aggarwal, B.B. Curcumin-induced suppression of cell proliferation correlates with down-regulation of cyclin D1 expression and CDK4-mediated retinoblastoma protein phosphorylation. Oncogene 2002, 21, 8852–8861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thoms, H.C.; Dunlop, M.G.; Stark, L.A. p38-mediated inactivation of cyclin D1/cyclin-dependent kinase 4 stimulates nucleolar translocation of RelA and apoptosis in colorectal cancer cells. Cancer Res. 2007, 67, 1660–1669. [Google Scholar] [CrossRef] [PubMed]
- Qiao, X.; Wang, X.; Shang, Y.; Li, Y.; Chen, S.Z. Azithromycin enhances anticancer activity of TRAIL by inhibiting autophagy and up-regulating the protein levels of DR4/5 in colon cancer cells in vitro and in vivo. Cancer Commun. 2018, 38, 43. [Google Scholar] [CrossRef] [PubMed]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qiao, X.; Gan, M.; Wang, C.; Liu, B.; Shang, Y.; Li, Y.; Chen, S. Tetracenomycin X Exerts Antitumour Activity in Lung Cancer Cells through the Downregulation of Cyclin D1. Mar. Drugs 2019, 17, 63. https://doi.org/10.3390/md17010063
Qiao X, Gan M, Wang C, Liu B, Shang Y, Li Y, Chen S. Tetracenomycin X Exerts Antitumour Activity in Lung Cancer Cells through the Downregulation of Cyclin D1. Marine Drugs. 2019; 17(1):63. https://doi.org/10.3390/md17010063
Chicago/Turabian StyleQiao, Xinran, Maoluo Gan, Chen Wang, Bin Liu, Yue Shang, Yi Li, and Shuzhen Chen. 2019. "Tetracenomycin X Exerts Antitumour Activity in Lung Cancer Cells through the Downregulation of Cyclin D1" Marine Drugs 17, no. 1: 63. https://doi.org/10.3390/md17010063
APA StyleQiao, X., Gan, M., Wang, C., Liu, B., Shang, Y., Li, Y., & Chen, S. (2019). Tetracenomycin X Exerts Antitumour Activity in Lung Cancer Cells through the Downregulation of Cyclin D1. Marine Drugs, 17(1), 63. https://doi.org/10.3390/md17010063