Griffithsin, a Highly Potent Broad-Spectrum Antiviral Lectin from Red Algae: From Discovery to Clinical Application


Abstract
:1. Introduction
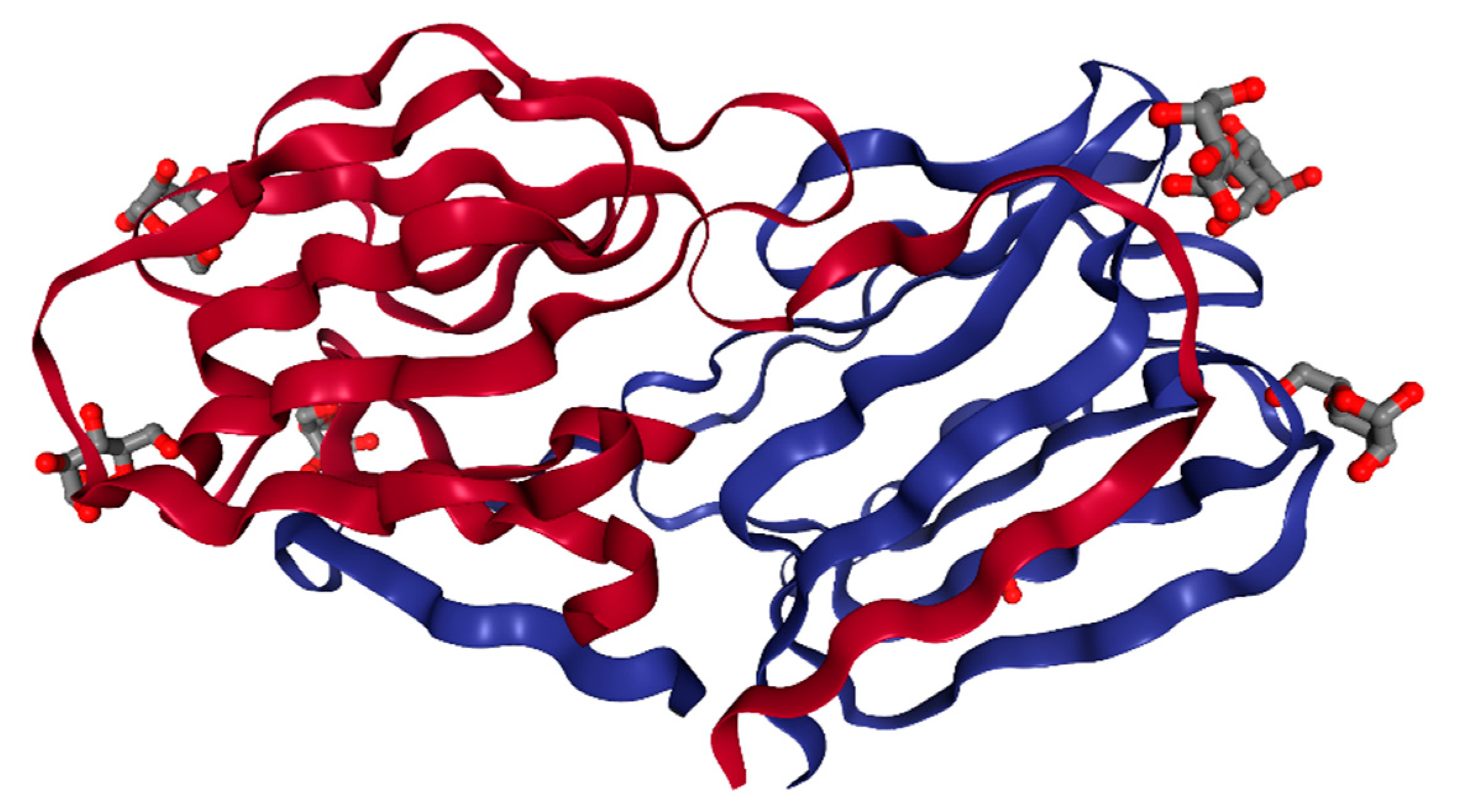
2. Structure
3. Anti-HIV-1 Activity and Cytotoxicity in Vitro
4. Drug Combination
5. Anti-HIV-1 Mechanism of Action
6. Resistance
7. Toxicity
8. Pharmacokinetic and in Vivo Efficacy Studies
9. Large-Scale Production
10. A Formulation for Efficient Delivery
11. Clinical Study
12. The Antimicrobial Activity of GRFT on Other Viruses
13. Conclusions
Funding
Conflicts of Interest
References
- Swanson, M.D.; Winter, H.C.; Goldstein, I.J.; Markovitz, D.M. A lectin isolated from bananas is a potent inhibitor of HIV replication. J. Biol. Chem. 2010, 285, 8646–8655. [Google Scholar] [CrossRef] [PubMed]
- O’Keefe, B.R.; Smee, D.F.; Turpin, J.A.; Saucedo, C.J.; Gustafson, K.R.; Mori, T.; Blakeslee, D.; Buckheit, R.; Boyd, M.R. Potent anti-influenza activity of cyanovirin-n and interactions with viral hemagglutinin. Antimicrobial. Agents Chemother. 2003, 47, 2518–2525. [Google Scholar] [CrossRef] [PubMed]
- Huskens, D.; Ferir, G.; Vermeire, K.; Kehr, J.C.; Balzarini, J.; Dittmann, E.; Schols, D. Microvirin, a novel alpha(1,2)-mannose-specific lectin isolated from microcystis aeruginosa, has anti-HIV-1 activity comparable with that of cyanovirin-n but a much higher safety profile. J. Biol. Chem. 2010, 285, 24845–24854. [Google Scholar] [CrossRef] [PubMed]
- Janahi, E.M.A.; Haque, S.; Akhter, N.; Wahid, M.; Jawed, A.; Mandal, R.K.; Lohani, M.; Areeshi, M.Y.; Almalki, S.; Das, S.; et al. Bioengineered intravaginal isolate of lactobacillus plantarum expresses algal lectin scytovirin demonstrating anti-HIV-1 activity. Microb. Pathog. 2018, 122, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Mori, T.; O’Keefe, B.R.; Sowder, R.C., 2nd; Bringans, S.; Gardella, R.; Berg, S.; Cochran, P.; Turpin, J.A.; Buckheit, R.W., Jr.; McMahon, J.B.; et al. Isolation and characterization of griffithsin, a novel HIV-inactivating protein, from the red alga Griffithsia sp. J. Biol. Chem. 2005, 280, 9345–9353. [Google Scholar] [CrossRef] [PubMed]
- Carr, A. Toxicity of antiretroviral therapy and implications for drug development. Nat. Rev. Drug Discov. 2003, 2, 624–634. [Google Scholar] [CrossRef]
- Abdool Karim, Q.; Abdool Karim, S.S.; Frohlich, J.A.; Grobler, A.C.; Baxter, C.; Mansoor, L.E.; Kharsany, A.B.; Sibeko, S.; Mlisana, K.P.; Omar, Z.; et al. Effectiveness and safety of tenofovir gel, an antiretroviral microbicide, for the prevention of HIV infection in women. Science 2010, 329, 1168–1174. [Google Scholar] [CrossRef]
- Beyrer, C.; Baral, S.D.; van Griensven, F.; Goodreau, S.M.; Chariyalertsak, S.; Wirtz, A.L.; Brookmeyer, R. Global epidemiology of HIV infection in men who have sex with men. Lancet 2012, 380, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Lusvarghi, S.; Bewley, C.A. Griffithsin: An antiviral lectin with outstanding therapeutic potential. Viruses 2016, 8, 296. [Google Scholar] [CrossRef]
- O’Keefe, B.R.; Giomarelli, B.; Barnard, D.L.; Shenoy, S.R.; Chan, P.K.; McMahon, J.B.; Palmer, K.E.; Barnett, B.W.; Meyerholz, D.K.; Wohlford-Lenane, C.L.; et al. Broad-spectrum in vitro activity and in vivo efficacy of the antiviral protein griffithsin against emerging viruses of the family coronaviridae. J. Virol. 2010, 84, 2511–2521. [Google Scholar] [CrossRef]
- Millet, J.K.; Seron, K.; Labitt, R.N.; Danneels, A.; Palmer, K.E.; Whittaker, G.R.; Dubuisson, J.; Belouzard, S. Middle east respiratory syndrome coronavirus infection is inhibited by griffithsin. Antiviral Res. 2016, 133, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Takebe, Y.; Saucedo, C.J.; Lund, G.; Uenishi, R.; Hase, S.; Tsuchiura, T.; Kneteman, N.; Ramessar, K.; Tyrrell, D.L.; Shirakura, M.; et al. Antiviral lectins from red and blue-green algae show potent in vitro and in vivo activity against hepatitis c virus. PLoS ONE 2013, 8, e64449. [Google Scholar] [CrossRef] [PubMed]
- Meuleman, P.; Albecka, A.; Belouzard, S.; Vercauteren, K.; Verhoye, L.; Wychowski, C.; Leroux-Roels, G.; Palmer, K.E.; Dubuisson, J. Griffithsin has antiviral activity against hepatitis C virus. Antimicrob. Agents Chemother. 2011, 55, 5159–5167. [Google Scholar] [CrossRef] [PubMed]
- Nixon, B.; Stefanidou, M.; Mesquita, P.M.; Fakioglu, E.; Segarra, T.; Rohan, L.; Halford, W.; Palmer, K.E.; Herold, B.C. Griffithsin protects mice from genital herpes by preventing cell-to-cell spread. J. Virol. 2013, 87, 6257–6269. [Google Scholar] [CrossRef] [PubMed]
- Levendosky, K.; Mizenina, O.; Martinelli, E.; Jean-Pierre, N.; Kizima, L.; Rodriguez, A.; Kleinbeck, K.; Bonnaire, T.; Robbiani, M.; Zydowsky, T.M.; et al. Griffithsin and carrageenan combination to target herpes simplex virus 2 and human papillomavirus. Antimicrob. Agents Chemother. 2015, 59, 7290–7298. [Google Scholar] [CrossRef]
- Ishag, H.Z.; Li, C.; Wang, F.; Mao, X. Griffithsin binds to the glycosylated proteins (E and prM) of japanese encephalitis virus and inhibit its infection. Virus Res. 2016, 215, 50–54. [Google Scholar] [CrossRef]
- Ishag, H.Z.; Li, C.; Huang, L.; Sun, M.X.; Wang, F.; Ni, B.; Malik, T.; Chen, P.Y.; Mao, X. Griffithsin inhibits japanese encephalitis virus infection in vitro and in vivo. Arch. Virol. 2013, 158, 349–358. [Google Scholar] [CrossRef]
- Li, L.; Yu, X.; Zhang, H.; Cheng, H.; Hou, L.; Zheng, Q.; Hou, J. In vitro antiviral activity of griffithsin against porcine epidemic diarrhea virus. Virus Genes 2019, 55, 174–181. [Google Scholar] [CrossRef]
- Ziolkowska, N.E.; O’Keefe, B.R.; Mori, T.; Zhu, C.; Giomarelli, B.; Vojdani, F.; Palmer, K.E.; McMahon, J.B.; Wlodawer, A. Domain-swapped structure of the potent antiviral protein griffithsin and its mode of carbohydrate binding. Structure 2006, 14, 1127–1135. [Google Scholar] [CrossRef]
- Moulaei, T.; Shenoy, S.R.; Giomarelli, B.; Thomas, C.; McMahon, J.B.; Dauter, Z.; O’Keefe, B.R.; Wlodawer, A. Monomerization of viral entry inhibitor griffithsin elucidates the relationship between multivalent binding to carbohydrates and anti-HIV activity. Structure 2010, 18, 1104–1115. [Google Scholar] [CrossRef]
- Xue, J.; Gao, Y.; Hoorelbeke, B.; Kagiampakis, I.; Zhao, B.; Demeler, B.; Balzarini, J.; Liwang, P.J. The role of individual carbohydrate-binding sites in the function of the potent anti-HIV lectin griffithsin. Mol. Pharm. 2012, 9, 2613–2625. [Google Scholar] [CrossRef] [PubMed]
- Ziolkowska, N.E.; Shenoy, S.R.; O’Keefe, B.R.; McMahon, J.B.; Palmer, K.E.; Dwek, R.A.; Wormald, M.R.; Wlodawer, A. Crystallographic, thermodynamic, and molecular modeling studies of the mode of binding of oligosaccharides to the potent antiviral protein griffithsin. Proteins 2007, 67, 661–670. [Google Scholar] [CrossRef] [PubMed]
- Ziolkowska, N.E.; Shenoy, S.R.; O’Keefe, B.R.; Wlodawer, A. Crystallographic studies of the complexes of antiviral protein griffithsin with glucose and n-acetylglucosamine. Protein Sci. 2007, 16, 1485–1489. [Google Scholar] [CrossRef] [PubMed]
- Micewicz, E.D.; Cole, A.L.; Jung, C.L.; Luong, H.; Phillips, M.L.; Pratikhya, P.; Sharma, S.; Waring, A.J.; Cole, A.M.; Ruchala, P. Grifonin-1: A small HIV-1 entry inhibitor derived from the algal lectin, griffithsin. PLoS ONE 2010, 5, e14360. [Google Scholar] [CrossRef] [PubMed]
- Alexandre, K.B.; Gray, E.S.; Lambson, B.E.; Moore, P.L.; Choge, I.A.; Mlisana, K.; Karim, S.S.; McMahon, J.; O’Keefe, B.; Chikwamba, R.; et al. Mannose-rich glycosylation patterns on HIV-1 subtype c gp120 and sensitivity to the lectins, griffithsin, cyanovirin-n and scytovirin. Virology 2010, 402, 187–196. [Google Scholar] [CrossRef]
- Alexandre, K.B.; Gray, E.S.; Pantophlet, R.; Moore, P.L.; McMahon, J.B.; Chakauya, E.; O’Keefe, B.R.; Chikwamba, R.; Morris, L. Binding of the mannose-specific lectin, griffithsin, to HIV-1 gp120 exposes the CD4-binding site. J. Virol. 2011, 85, 9039–9050. [Google Scholar] [CrossRef]
- Emau, P.; Tian, B.; O’Keefe B, R.; Mori, T.; McMahon, J.B.; Palmer, K.E.; Jiang, Y.; Bekele, G.; Tsai, C.C. Griffithsin, a potent HIV entry inhibitor, is an excellent candidate for anti-HIV microbicide. J. Med. Primatol. 2007, 36, 244–253. [Google Scholar] [CrossRef]
- Ferir, G.; Huskens, D.; Palmer, K.E.; Boudreaux, D.M.; Swanson, M.D.; Markovitz, D.M.; Balzarini, J.; Schols, D. Combinations of griffithsin with other carbohydrate-binding agents demonstrate superior activity against HIV type 1, HIV type 2, and selected carbohydrate-binding agent-resistant HIV type 1 strains. AIDS Res. Hum. Retroviruses 2012, 28, 1513–1523. [Google Scholar] [CrossRef]
- Ferir, G.; Palmer, K.E.; Schols, D. Synergistic activity profile of griffithsin in combination with tenofovir, maraviroc and enfuvirtide against HIV-1 clade c. Virology 2011, 417, 253–258. [Google Scholar] [CrossRef]
- Hoorelbeke, B.; Xue, J.; LiWang, P.J.; Balzarini, J. Role of the carbohydrate-binding sites of griffithsin in the prevention of DC-SIGN-mediated capture and transmission of HIV-1. PLoS ONE 2013, 8, e64132. [Google Scholar] [CrossRef]
- Hu, B.; Du, T.; Li, C.; Luo, S.; Liu, Y.; Huang, X.; Hu, Q. Sensitivity of transmitted and founder human immunodeficiency virus type 1 envelopes to carbohydrate-binding agents griffithsin, cyanovirin-n and galanthus nivalis agglutinin. J. Gen. Virol. 2015, 96, 3660–3666. [Google Scholar] [CrossRef] [PubMed]
- Kagiampakis, I.; Gharibi, A.; Mankowski, M.K.; Snyder, B.A.; Ptak, R.G.; Alatas, K.; LiWang, P.J. Potent strategy to inhibit HIV-1 by binding both gp120 and gp41. Antimicrob. Agents Chemother. 2011, 55, 264–275. [Google Scholar] [CrossRef] [PubMed]
- Ferir, G.; Palmer, K.E.; Schols, D. Griffithsin, alone and combined with all classes of antiretroviral drugs, potently inhibits HIV cell-cell transmission and destruction of CD4+ T cells. Antivir. Antiretrovir. 2012, 4, 103. [Google Scholar] [CrossRef]
- Banerjee, K.; Michael, E.; Eggink, D.; van Montfort, T.; Lasnik, A.B.; Palmer, K.E.; Sanders, R.W.; Moore, J.P.; Klasse, P.J. Occluding the mannose moieties on human immunodeficiency virus type 1 gp120 with griffithsin improves the antibody responses to both proteins in mice. AIDS Res. Hum. Retroviruses 2012, 28, 206–214. [Google Scholar] [CrossRef] [PubMed]
- Lusvarghi, S.; Lohith, K.; Morin-Leisk, J.; Ghirlando, R.; Hinshaw, J.E.; Bewley, C.A. Binding site geometry and subdomain valency control effects of neutralizing lectins on HIV-1 viral particles. ACS Infect. Dis 2016, 2, 882–891. [Google Scholar] [CrossRef] [PubMed]
- Moulaei, T.; Alexandre, K.B.; Shenoy, S.R.; Meyerson, J.R.; Krumpe, L.R.; Constantine, B.; Wilson, J.; Buckheit, R.W., Jr.; McMahon, J.B.; Subramaniam, S.; et al. Griffithsin tandemers: Flexible and potent lectin inhibitors of the human immunodeficiency virus. Retrovirology 2015, 12, 6. [Google Scholar] [CrossRef]
- Xue, J.; Hoorelbeke, B.; Kagiampakis, I.; Demeler, B.; Balzarini, J.; Liwang, P.J. The griffithsin dimer is required for high-potency inhibition of HIV-1: Evidence for manipulation of the structure of gp120 as part of the griffithsin dimer mechanism. Antimicrob. Agents Chemother. 2013, 57, 3976–3989. [Google Scholar] [CrossRef]
- Alexandre, K.B.; Gray, E.S.; Mufhandu, H.; McMahon, J.B.; Chakauya, E.; O’Keefe, B.R.; Chikwamba, R.; Morris, L. The lectins griffithsin, cyanovirin-n and scytovirin inhibit HIV-1 binding to the DC-SIGN receptor and transfer to CD4(+) cells. Virology 2012, 423, 175–186. [Google Scholar] [CrossRef]
- Alexandre, K.B.; Moore, P.L.; Nonyane, M.; Gray, E.S.; Ranchobe, N.; Chakauya, E.; McMahon, J.B.; O’Keefe, B.R.; Chikwamba, R.; Morris, L. Mechanisms of HIV-1 subtype C resistance to GRFT, CV-N and SVN. Virology 2013, 446, 66–76. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Jin, W.; Griffin, G.E.; Shattock, R.J.; Hu, Q. Removal of two high-mannose N-linked glycans on gp120 renders human immunodeficiency virus 1 largely resistant to the carbohydrate-binding agent griffithsin. J. Gen. Virol. 2011, 92, 2367–2373. [Google Scholar] [CrossRef]
- Barton, C.; Kouokam, J.C.; Lasnik, A.B.; Foreman, O.; Cambon, A.; Brock, G.; Montefiori, D.C.; Vojdani, F.; McCormick, A.A.; O’Keefe, B.R.; et al. Activity of and effect of subcutaneous treatment with the broad-spectrum antiviral lectin griffithsin in two laboratory rodent models. Antimicrob. Agents Chemother. 2014, 58, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Girard, L.; Birse, K.; Holm, J.B.; Gajer, P.; Humphrys, M.S.; Garber, D.; Guenthner, P.; Noel-Romas, L.; Abou, M.; McCorrister, S.; et al. Impact of the griffithsin anti-HIV microbicide and placebo gels on the rectal mucosal proteome and microbiome in non-human primates. Sci. Rep. 2018, 8, 8059. [Google Scholar] [CrossRef] [PubMed]
- Kouokam, J.C.; Huskens, D.; Schols, D.; Johannemann, A.; Riedell, S.K.; Walter, W.; Walker, J.M.; Matoba, N.; O’Keefe, B.R.; Palmer, K.E. Investigation of griffithsin’s interactions with human cells confirms its outstanding safety and efficacy profile as a microbicide candidate. PLoS ONE 2011, 6, e22635. [Google Scholar] [CrossRef] [PubMed]
- Kouokam, J.C.; Lasnik, A.B.; Palmer, K.E. Studies in a murine model confirm the safety of griffithsin and advocate its further development as a microbicide targeting HIV-1 and other enveloped viruses. Viruses 2016, 8, 331. [Google Scholar] [CrossRef]
- O’Keefe, B.R.; Vojdani, F.; Buffa, V.; Shattock, R.J.; Montefiori, D.C.; Bakke, J.; Mirsalis, J.; d’Andrea, A.L.; Hume, S.D.; Bratcher, B.; et al. Scaleable manufacture of HIV-1 entry inhibitor griffithsin and validation of its safety and efficacy as a topical microbicide component. Proc. Natl. Acad. Sci. USA 2009, 106, 6099–6104. [Google Scholar] [CrossRef] [Green Version]
- Barton, C.; Kouokam, J.C.; Hurst, H.; Palmer, K.E. Pharmacokinetics of the antiviral lectin griffithsin administered by different routes indicates multiple potential uses. Viruses 2016, 8, 331. [Google Scholar] [CrossRef]
- Farr Zuend, C.; Nomellini, J.F.; Smit, J.; Horwitz, M.S. A caulobacter crescentus microbicide protects from vaginal infection with HIV-1JR-CSF in humanized bone marrow-liver-thymus mice. J. Virol. 2019, 3, e00614-19. [Google Scholar] [CrossRef]
- Fuqua, J.L.; Wanga, V.; Palmer, K.E. Improving the large scale purification of the HIV microbicide, griffithsin. BMC Biotechnol. 2015, 15, 12. [Google Scholar] [CrossRef]
- Giomarelli, B.; Schumacher, K.M.; Taylor, T.E.; Sowder, R.C., 2nd; Hartley, J.L.; McMahon, J.B.; Mori, T. Recombinant production of anti-HIV protein, griffithsin, by auto-induction in a fermentor culture. Protein Expr. Purif. 2006, 47, 194–202. [Google Scholar] [CrossRef]
- Hahn, S.; Giritch, A.; Bartels, D.; Bortesi, L.; Gleba, Y. A novel and fully scalable agrobacterium spray-based process for manufacturing cellulases and other cost-sensitive proteins in plants. Plant. Biotechnol. J. 2015, 13, 708–716. [Google Scholar] [CrossRef]
- Petrova, M.I.; van den Broek, M.F.L.; Spacova, I.; Verhoeven, T.L.A.; Balzarini, J.; Vanderleyden, J.; Schols, D.; Lebeer, S. Engineering lactobacillus rhamnosus GG and GR-1 to express HIV-inhibiting griffithsin. Int. J. Antimicrob. Agents 2018, 52, 599–607. [Google Scholar] [CrossRef] [PubMed]
- Vamvaka, E.; Arcalis, E.; Ramessar, K.; Evans, A.; O’Keefe, B.R.; Shattock, R.J.; Medina, V.; Stoger, E.; Christou, P.; Capell, T. Rice endosperm is cost-effective for the production of recombinant griffithsin with potent activity against HIV. Plant. Biotechnol. J. 2016, 14, 1427–1437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoelscher, M.; Tiller, N.; Teh, A.Y.; Wu, G.Z.; Ma, J.K.; Bock, R. High-level expression of the HIV entry inhibitor griffithsin from the plastid genome and retention of biological activity in dried tobacco leaves. Plant. Mol. Biol. 2018, 97, 357–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, B.M.; Lotter-Stark, H.C.T.; Rybicki, E.P.; Chikwamba, R.K.; Palmer, K.E. Characterization of the hypersensitive response-like cell death phenomenon induced by targeting antiviral lectin griffithsin to the secretory pathway. Plant. Biotechnol. J. 2018, 16, 1811–1821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vamvaka, E.; Farre, G.; Molinos-Albert, L.M.; Evans, A.; Canela-Xandri, A.; Twyman, R.M.; Carrillo, J.; Ordonez, R.A.; Shattock, R.J.; O’Keefe, B.R.; et al. Unexpected synergistic HIV neutralization by a triple microbicide produced in rice endosperm. Proc. Natl. Acad. Sci. USA 2018, 115, E7854–E7862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moncla, B.J.; Pryke, K.; Rohan, L.C.; Graebing, P.W. Degradation of naturally occurring and engineered antimicrobial peptides by proteases. Adv. Biosci. Biotechnol. 2011, 2, 404–408. [Google Scholar] [CrossRef] [Green Version]
- Grooms, T.N.; Vuong, H.R.; Tyo, K.M.; Malik, D.A.; Sims, L.B.; Whittington, C.P.; Palmer, K.E.; Matoba, N.; Steinbach-Rankins, J.M. Griffithsin-modified electrospun fibers as a delivery scaffold to prevent HIV infection. Antimicrob. Agents Chemother. 2016, 60, 6518–6531. [Google Scholar] [CrossRef]
- Lal, M.; Lai, M.; Ugaonkar, S.; Wesenberg, A.; Kizima, L.; Rodriguez, A.; Levendosky, K.; Mizenina, O.; Fernandez-Romero, J.; Zydowsky, T. Development of a vaginal fast-dissolving insert combining griffithsin and carrageenan for potential use against sexually transmitted infections. J. Pharm. Sci. 2018, 107, 2601–2610. [Google Scholar] [CrossRef]
- Tyo, K.M.; Duan, J.; Kollipara, P.; Dela Cerna, M.V.C.; Lee, D.; Palmer, K.E.; Steinbach-Rankins, J.M. pH-responsive delivery of griffithsin from electrospun fibers. Eur. J. Pharm. Biopharm. 2019, 138, 64–74. [Google Scholar] [CrossRef]
- Yang, H.; Li, J.; Patel, S.K.; Palmer, K.E.; Devlin, B.; Rohan, L.C. Design of poly(lactic-co-glycolic acid) (PLGA) nanoparticles for vaginal co-delivery of griffithsin and dapivirine and their synergistic effect for HIV prophylaxis. Pharmaceutics 2019, 11, 184. [Google Scholar] [CrossRef]
- Study to Evaluate the Safety of Griffithsin in a Carrageenan Gel in Healthy Women. Available online: https://clinicaltrials.gov/ct2/show/study/NCT02875119 (accessed on 5 October 2019).
- Palmer, K.E. Griffithsin-based Rectal Microbicides for Prevention of Viral Entry (PREVENT). Available online: https://clinicaltrials.gov/ct2/show/NCT04032717?term=griffithsin&rank=2 (accessed on 5 October 2019).
- Developing and Testing a Griffithsin (non-ARV) Microbicide. Available online: https://www.popcouncil.org/research/developing-and-testing-a-griffithsin-non-arv-microbicide (accessed on 5 October 2019).
- Farr Zuend, C.; Nomellini, J.F.; Smit, J.; Horwitz, M.S. Generation of a dual-target, safe, inexpensive microbicide that protects against HIV-1 and HSV-2 disease. Sci. Rep. 2018, 8, 2786. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo, L.; Oliveira, C.; Fournier, C.; Descamps, V.; Morel, V.; Dubuisson, J.; Brochot, E.; Francois, C.; Castelain, S.; Duverlie, G.; et al. Hepatitis C virus resistance to carbohydrate-binding agents. PLoS ONE 2016, 11, e0149064. [Google Scholar] [CrossRef] [PubMed]
- Derby, N.; Lal, M.; Aravantinou, M.; Kizima, L.; Barnable, P.; Rodriguez, A.; Lai, M.; Wesenberg, A.; Ugaonkar, S.; Levendosky, K.; et al. Griffithsin carrageenan fast dissolving inserts prevent SHIV HSV-2 and HPV infections in vivo. Nat. Commun. 2018, 9, 3881. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Structure Studied | Crystal Resolution (A°) | Structural and Functional Characteristics | Ref. |
|---|---|---|---|
| GRFT alone or with mannose | 1.3 and 0.94 | A domain-swapped dimer, a Jacalin-related lectin with a β-prism motif, and three identical mannose-binding sites on each monomer | [19] |
| GRFT with glucose or N-acetylglucosamine | 30–1.50 and 30–1.56 | All six monosaccharide binding sites of GRFT are occupied by both glucose and N-acetylglucosamine with a mode of binding similar to that of mannose | [23] |
| GRFT with 1-6α-mannobiose or maltose | 2.0 and 1.5 | The binding of 1-6α-mannobiose is similar to that of mannose and the binding of maltose is weaker than that of mannose | [22] |
| Monomeric GRFT | 30–0.97 | Reduced activity against HIV-1 due to a loss of multivalent interaction and the binding of a monomeric GRFT to two different nona-mannosides | [20] |
| GRFT with a disrupted carbohydrate-binding site | NMR study | Reduced binding to mannose and a weaker correlation between anti-HIV-1 activity and gp120 binding | [21] |
| HIV-1 Type | Cell Line | Anti-HIV-1 Assay Readout | EC50 (nM) | CC50 (nM) | Ref. |
|---|---|---|---|---|---|
| Laboratory strains and primary isolates of T- and M-tropic HIV-1 | CEM-SS cells, PBMC, and macrophage | Virus-induced cell killing | 0.043–0.63 | >783 | [5] |
| CXCR4-and CCR5-tropic HIV-1, SHIV strains, and SIVmac251 | CEM-174, MT-2, MT-4, and PBMC | p24 | <1 | >500 | [27] |
| HIV-1 subtype C primary virus isolates | TZM-bl and PBMC | TZM-bl and p24 | 0.4 | N/A | [25] |
| HIV-1 CCR5-tropic strain ADA and CXCR4-tropic strain HXB2 | HeLa-ADA effector cells and HeLa-P5L CCR5-bearing target cells | CCR5-tropic cell-cell fusion | 1.31 | N/A | [32] |
| HIV-1 clade B and clade C isolates | PBMCs and CD4+ MT-4 cells | p24 HIV-1 core Ag ELISA | 0.18 | N/A | [29] |
| HIV-1 subtype B QH0515 and C SW7 and Du179 | Raji/DC-SIGN cells and TZM-bl cells | HIV-1 binding to the DC-SIGN receptor | 345 | N/A | [26] |
| HIV-1 subtype B QH0515 and C SW7 and Du179 | Raji/DC-SIGN cells and TZM-bl cells | HIV-1 bound Raji cells transferred to CD4 cells | 4.8–35 | N/A | [26] |
| HIV-1 X4 strain IIIB and dual-tropic R5/X4 HIV-1 strain HE | Persistently HIV-infected T cells and noninfected CD4+ target T cells | Giant cell formation | 0.087 | N/A | [28] |
| HIV-1 X4 strain IIIB and dual-tropic R5/X4 HIV-1 strain HE | Raji.DC-SIGN cells | DC-SIGN mediated HIV transmission | 0.025 | N/A | [28] |
| HIV-1(IIIB) and HIV NL4.3 | CEM, C8166, HuT-78, and Sup-T1 cells, Raji/DC-SIGN cells | DC-SIGN-mediated virus capture | 1.5 | N/A | [30] |
| HIV-1(IIIB) and HIV NL4.3 | CEM, C8166, HuT-78, and Sup-T1 cells, Raji/DC-SIGN cells | HIV-1 transmission to CD4+ T-lymphocytes | 0.012 | N/A | [30] |
| Single transmitted/founder HIV-1 | TZM-bl cells | Neutralization assay using pseudotyped viruses | 0.006–10 | N/A | [31] |
| Cell Line | Assay Readout | Combination Drug | Result | Ref. |
|---|---|---|---|---|
| PBMCs and CD4+ MT-4 cells | p24 HIV-1 core Ag ELISA | Tenofovir, maraviroc, and enfuvirtide | Synergy | [29] |
| HeLa-ADA effector cells and HeLa-P5L CCR5-bearing target cells | CCR5-tropic cell-cell fusion | Covalently linked gp41-binding peptide C37 | Synergy | [32] |
| TZM-bl cells, U87-CCR5 and U87-CXCR4. | Pseudovirus neutralization | 2G12 mAb | Synergy | [26] |
| Persistently HIV-infected T cells and noninfected CD4+ target T cells | Giant cell formation | Efavirenz, tenofovir, raltegravir, and elvitegravir | Synergy | [28] |
| Raji.DC-SIGN cells | DC-SIGN mediated HIV transmission | Tenofovir, saquinavir, and 2G12 mAb | Synergy | [28] |
| MT-4 cells, PBMC | Virus-induced cytopathic effect | Microvirin, 2G12 mAb, BanLec, and HHA | Synergy | [28] |
| The Effect of GRFT on HIV-1 gp120 | Ref. |
|---|---|
| Exposure of the CD4 binding site of gp120 through the glycan at position 386 and blockage of coreceptor binding step | [26] |
| Inhibition of mannose-binding to gp120 and improvement of the humoral immune response to gp120 | [34] |
| Inhibition of gp120 binding to DC-SIGN and expulsion of gp120 from the gp120/DC-SIGN complex | [30] |
| Alteration of gp120 structure through the exposure of the CD4 binding site | [37] |
| Intra-virion crosslinking of gp120 | [36] |
| Inter-virion aggregation or clustering of gp120 | [35] |
| HIV Type | Cell Line | Assay Readout | Resistance Mechanism | Ref. |
|---|---|---|---|---|
| HIV-1 subtype C primary virus isolates | TZM-bl and PBMC | HIV-1 neutralization assay | Loss of glycans at positions 234 and 295 | [25] |
| Primary and T-cell-line-adapted HIV-1 isolates | TZM-bl cells | Env-pseudotyped viruses to infect TZM-bl cells | Loss of glycan at position 295 and 448 | [40] |
| HIV-1 subtype C primary virus isolates | Raji/DC-SIGN cells | The DC-SIGN-mediated HIV-1 transfer to TZM-bl cells | Introduction of a glycan at position 234 | [38] |
| HIV-1 subtype C primary virus isolates | TheTZM-bl and PBMC | HIV-1 neutralization assay in PBMC | Loss of glycan at positions 230, 234, 339, 392, and 448 | [39] |
| Single transmitted/founder HIV-1 | TZM-bl cells | Neutralization assay using pseudotyped viruses | Loss of glycan at positions 295, 339, and 448 | [31] |
| Models Used | Dose Tested | Effects of GRFT | Ref. |
|---|---|---|---|
| Human cervical explants and rabbits | Up to 2 μM | No effect on the production of proinflammatory cytokines and chemokines. No vaginal irritation in rabbits. | [45] |
| Human cervical epithelial cells, cervicovaginal cells, and PBMCs | Up to 84 μM | Minimal changes in secretion of inflammatory cytokines and chemokines No measurable effect on cell viability and T-cell activation markers. | [43] |
| Guinea pig BALB/c mice | Single 50 mg/kg and 10 daily subcutaneous injections of 10 mg/kg | Minimal overall toxicity. Well tolerated. Increase in spleen and liver mass. | [41] |
| Mouse PBMC and BALB/c mice with parenteral administration | Up to 4 μM in vitro, subcutaneously with a single dose of 10 mg/kg, fourteen daily doses of 10 mg/kg, 2 mg/kg subcutaneous, intravaginal, and intraperitoneal administration | No mitogenic properties in vitro. No effect on cell surface activation markers or animal fitness. No major organ toxicity with reversible splenomegaly. | [44] |
| Rhesus macaques | Intravaginal 0.1% gels | No change in rental proteome or microbiome | [42] |
| Models Used | Dose Tested | Effects of GRFT | Ref. |
|---|---|---|---|
| Guinea pig BALB/c mice | Single 50 mg/kg and 10 daily subcutaneous injections of 10 mg/kg. | Retention of antiviral activity in serum | [41] |
| Sprague Dawley (SD) rats | A single dose of 10 mg/mL intravenously or subcutaneously. Ten 40 mg/kg doses for 10 days. | Neutralization activity found in fecal extracts | [46] |
| Humanized bone marrow-liver-thymus mice | 108 GRFT-expressing recombinant C. crescentus intravaginally. | Protection against HIV-1 infection | [47] |
| Production Method | Expression Organism | Effects and Yield | Ref. |
|---|---|---|---|
| Transformation and the use of an autoinduction fermentor | Escherichia coli | 45-fold increase | [49] |
| Transduction with tobacco mosaic virus | Nicotiana benthamiana | Multigram quantity | [45] |
| Agrobacterium vectors | Nicotiana benthamiana | 90% of the leaf cells and 50% of the total soluble protein | [50] |
| Transduction with tobacco mosaic virus in pH 4 buffer, heating the extract to 55 °C, a bentonite MgCl2 mixture, and chromatography. | Nicotiana benthamiana | 88% ± 5% of griffithsin from the initial extract | [48] |
| Particle bombardment | Rice endosperm | 223 μg/g dry seed weight | [52] |
| Use of probiotic microorganisms | Lactobacillus rhamnosus GG and L. rhamnosus GR-1 | N/A | [51] |
| Chloroplast transformation | Nicotiana tabacum | 360 μg of pure griffithsin per gram | [53] |
| Formulation Method | Delivery Route | Effects on Delivery | Ref. |
|---|---|---|---|
| PLGA nanoparticles | Vaginal | A biphasic release with an initial burst phase followed by a sustained release phase | [60] |
| Electrospun fibers | In vitro | Maintenance of antiviral efficacy | [57] |
| FDI comprised of | Vaginal | Good friability, hardness, and stability | [58] |
| mPEG-PLGA:PBA-co-PAA | Vaginal | High GRFT loading and pH-dependent release | [59] |
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, C. Griffithsin, a Highly Potent Broad-Spectrum Antiviral Lectin from Red Algae: From Discovery to Clinical Application. Mar. Drugs 2019, 17, 567. https://doi.org/10.3390/md17100567
Lee C. Griffithsin, a Highly Potent Broad-Spectrum Antiviral Lectin from Red Algae: From Discovery to Clinical Application. Marine Drugs. 2019; 17(10):567. https://doi.org/10.3390/md17100567
Chicago/Turabian StyleLee, Choongho. 2019. "Griffithsin, a Highly Potent Broad-Spectrum Antiviral Lectin from Red Algae: From Discovery to Clinical Application" Marine Drugs 17, no. 10: 567. https://doi.org/10.3390/md17100567
APA StyleLee, C. (2019). Griffithsin, a Highly Potent Broad-Spectrum Antiviral Lectin from Red Algae: From Discovery to Clinical Application. Marine Drugs, 17(10), 567. https://doi.org/10.3390/md17100567