Characteristics of a Novel Manganese Superoxide Dismutase of a Hadal Sea Cucumber (Paelopatides sp.) from the Mariana Trench

Abstract
:1. Introduction
2. Results
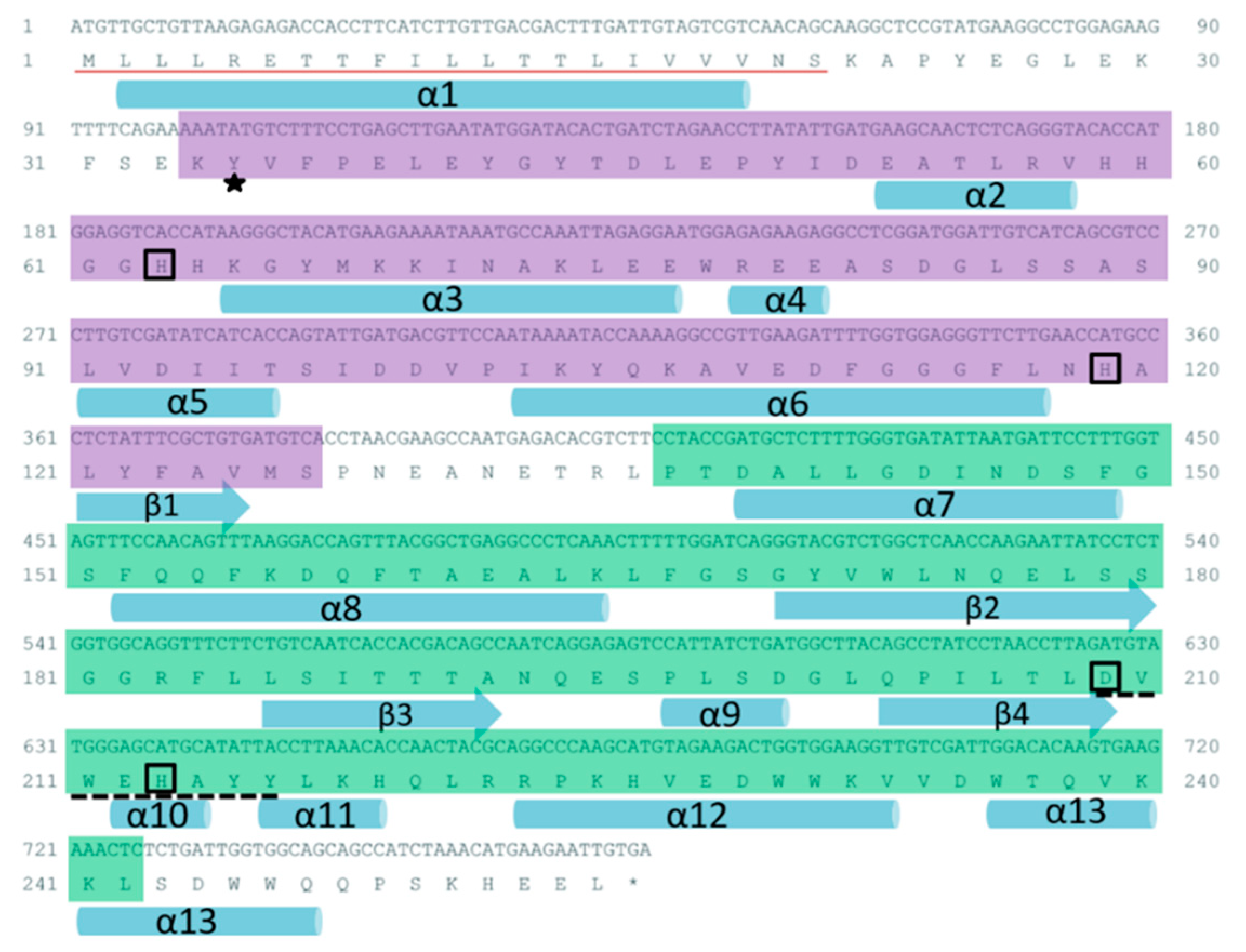
2.1. Sequence Characteristics
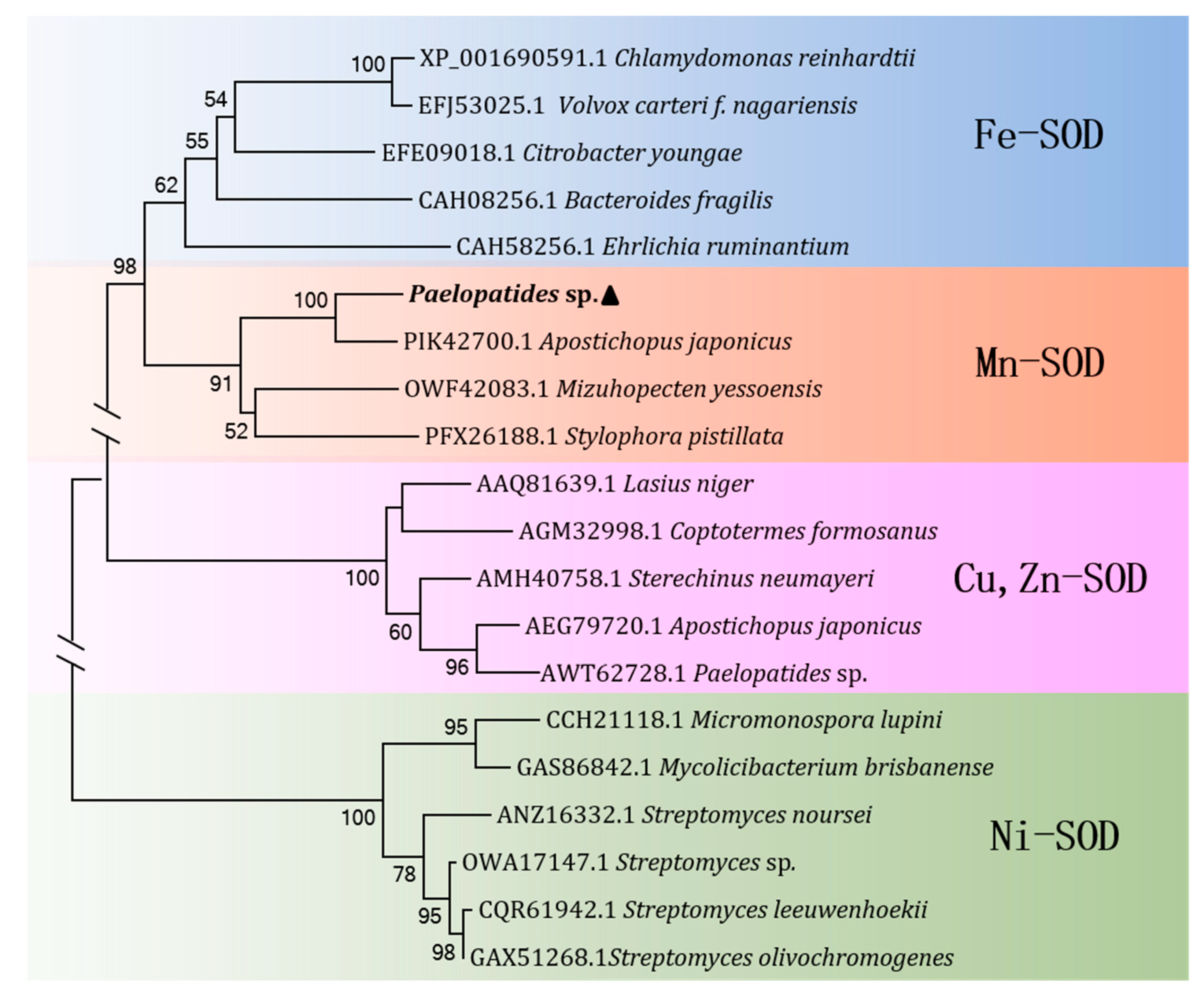
2.2. Homology and Phylogenetic Analysis
2.3. Expression, Purification, and Validation of Ps-Mn-SOD
2.4. Characterizations of Ps-Mn-SOD
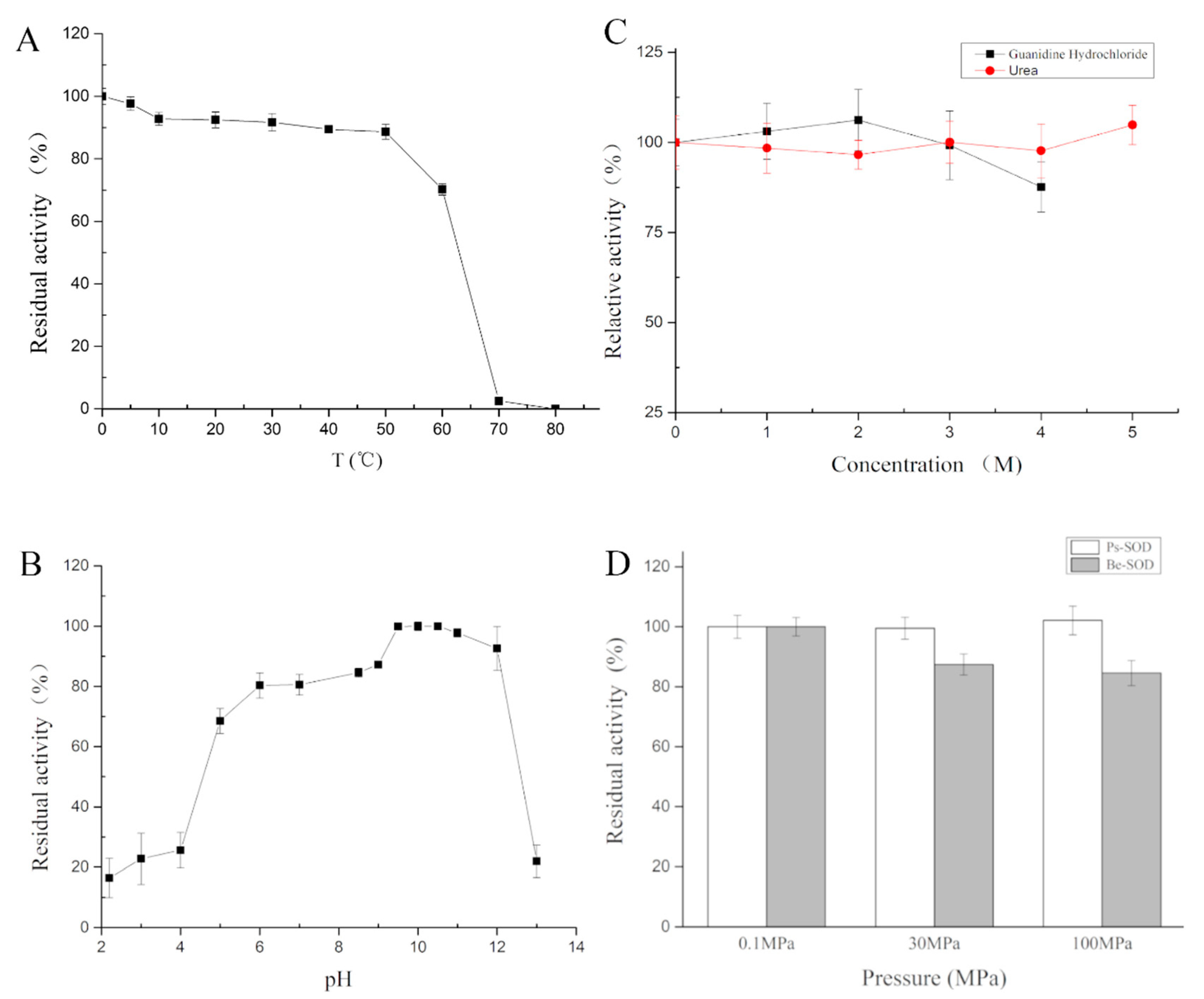
2.4.1. Effects of Temperature on Ps-Mn-SOD
2.4.2. Effects of pH on Ps-Mn-SOD
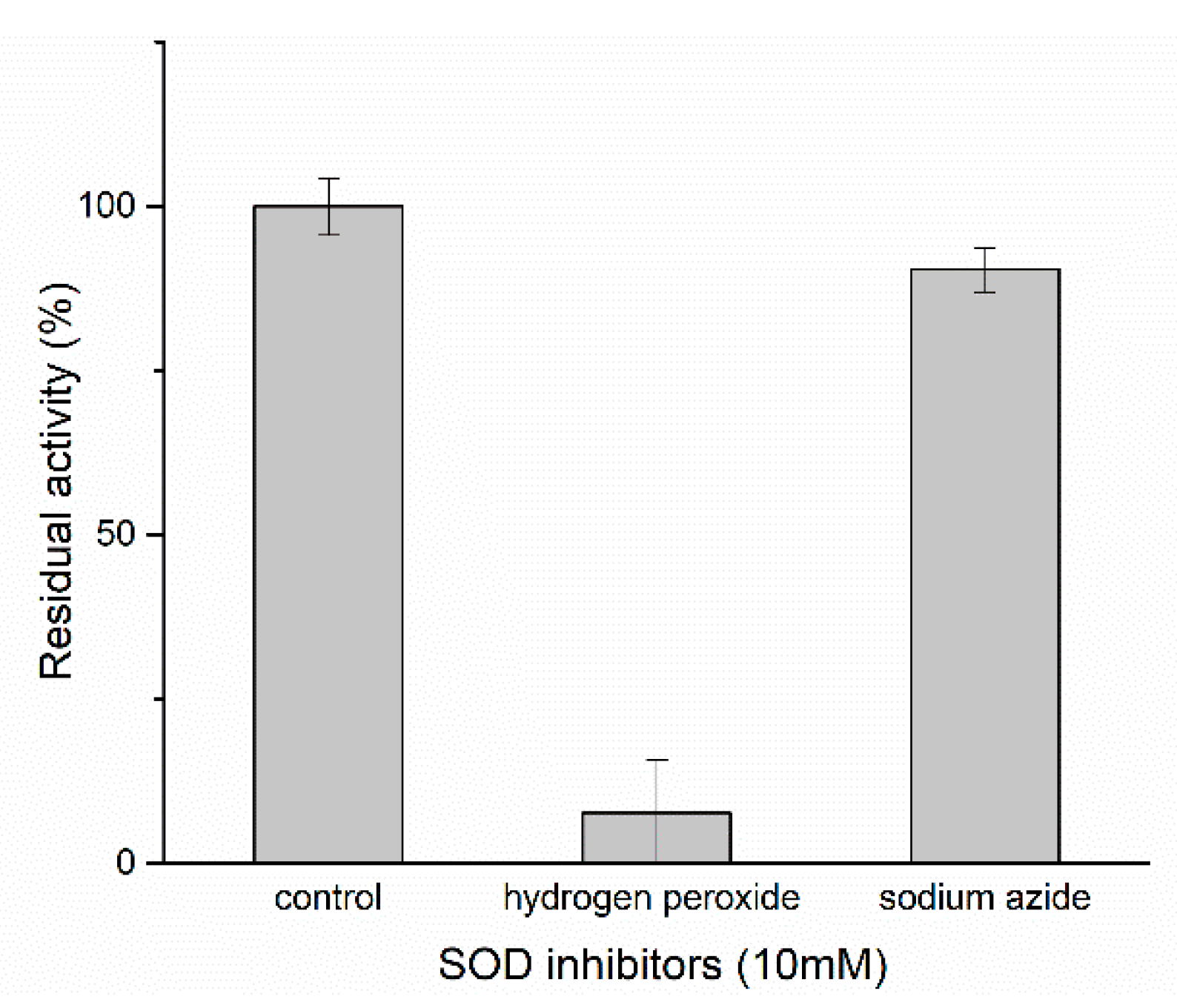
2.4.3. Effects of Chemicals on Ps-Mn-SOD
2.4.4. Effects of Digestive Enzymes on Ps-Mn-SOD
2.4.5. Effects of High Hydrostatic Pressure on Ps-Mn-SOD
2.4.6. Kinetic Parameters
3. Discussion
4. Materials and Methods
4.1. Material and Reagents
4.2. Cloning and Recombinant
4.3. Protein Overproduction, Purification, and Confirmation
4.4. Bioinformatics Analyses
4.5. Enzyme Assays
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Torres, M.A. ROS in biotic interactions. Physiol. Plant. 2010, 138, 414–429. [Google Scholar] [CrossRef] [PubMed]
- Kawanishi, S.; Inoue, S. Damage to DNA by reactive oxygen and nitrogen species. Seikagaku 1997, 69, 1014–1017. [Google Scholar] [PubMed]
- Kim, F.J.; Kim, H.P.; Hah, Y.C.; Roe, J.H. Differential expression of superoxide dismutases containing Ni and Fe/Zn in Streptomyces coelicolor. Eur. J. Biochem. 1996, 241, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Wu, B.; Liu, Z.; Tian, J.; Yu, T.; Zhou, L.; Sun, X.; Yang, A.G. A manganese superoxide dismutase (MnSOD) from ark shell, Scapharca broughtonii: Molecular characterization, expression and immune activity analysis. Fish Shellfish Immunol. 2015, 45, 656–665. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; Jiang, Y.H.; Miao, J.L.; Wang, Q.F.; Zhang, B.T.; Li, G.Y. Purification and characterization of a cold-active iron superoxide dismutase from a psychrophilic bacterium, Marinomonas sp. NJ522. Biotechnol. Lett. 2006, 28, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Krauss, I.R.; Merlino, A.; Pica, A.; Rullo, R.; Bertoni, A.; Capasso, A.; Amato, M.; Riccitiello, F.; De Vendittis, E.; Sica, F. Fine tuning of metal-specific activity in the Mn-like group of cambialistic superoxide dismutases. RSC Adv. 2015, 5, 87876–87887. [Google Scholar] [CrossRef]
- Sheng, Y.; Abreu, I.A.; Cabelli, D.E.; Maroney, M.J.; Miller, A.-F.; Teixeira, M.; Valentine, J.S. Superoxide Dismutases and Superoxide Reductases. Chem. Rev. 2014, 114, 3854–3918. [Google Scholar] [CrossRef] [PubMed]
- Ncn, P.; Godahewa, G.I.; Lee, S.; Kim, M.J.; Hwang, J.Y.; Kwon, M.G.; Hwang, S.D.; Lee, J. Manganese-superoxide dismutase (MnSOD), a role player in seahorse (Hippocampus abdominalis) antioxidant defense system and adaptive immune system. Fish Shellfish Immunol. 2017, 68, 435–442. [Google Scholar]
- Kim, B.M.; Rhee, J.S.; Park, G.S.; Lee, J.; Lee, Y.M.; Lee, J.S. Cu/Zn- and Mn-superoxide dismutase (SOD) from the copepod Tigriopus japonicus: Molecular cloning and expression in response to environmental pollutants. Chemosphere 2011, 84, 1467–1475. [Google Scholar] [CrossRef]
- Li, C.; He, J.; Su, X.; Li, T. A manganese superoxide dismutase in blood clam Tegillarca granosa: Molecular cloning, tissue distribution and expression analysis. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2011, 159, 64–70. [Google Scholar] [CrossRef]
- Wang, H.; Yang, H.; Liu, J.; Yanhong, L.I.; Liu, Z. Combined effects of temperature and copper ion concentration on the superoxide dismutase activity in Crassostrea ariakensis. Acta Oceanol. Sin. 2016, 35, 51–57. [Google Scholar] [CrossRef]
- Xie, Z.; Jian, H.; Jin, Z.; Xiao, X. Enhancing the adaptability of the deep-sea bacterium Shewanella piezotolerans WP3 to high pressure and low temperature by experimental evolution under H2O2 stress. Appl. Environ. Microbiol. 2017, 84, e02342-17. [Google Scholar] [CrossRef] [PubMed]
- Lebovitz, R.M.; Zhang, H.; Vogel, H.; Cartwright, J.; Dionne, L.; Lu, N.; Huang, S.; Matzuk, M.M. Neurodegeneration, myocardial injury, and perinatal death in mitochondrial superoxide dismutase-deficient mice. Proc. Natl. Acad. Sci. USA 1996, 93, 9782–9787. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Huang, T.T.; Carlson, E.J.; Melov, S.; Ursell, P.C.; Olson, J.L.; Noble, L.J.; Yoshimura, M.P.; Berger, C.; Chan, P.H. Dilated cardiomyopathy and neonatal lethality in mutant mice lacking manganese superoxide dismutase. Nat. Genet. 1995, 11, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Delacourte, A.; Defossez, A.; Ceballos, I.; Nicole, A.; Sinet, P.M. Preferential localization of copper zinc superoxide dismutase in the vulnerable cortical neurons in Alzheimer’s disease. Neurosci. Lett. 1988, 92, 247–253. [Google Scholar] [CrossRef]
- Kruman, I.I.; Pedersen, W.A.; Springer, J.E.; Mattson, M.P. ALS-linked Cu/Zn-SOD mutation increases vulnerability of motor neurons to excitotoxicity by a mechanism involving increased oxidative stress and perturbed calcium homeostasis. Exp. Neurol. 1999, 160, 28–39. [Google Scholar] [CrossRef] [PubMed]
- Cullen, J.J.; Weydert, C.; Hinkhouse, M.M.; Ritchie, J.; Domann, F.E.; Spitz, D.; Oberley, L.W. The role of manganese superoxide dismutase in the growth of pancreatic adenocarcinoma. Cancer Res. 2003, 63, 1297–1303. [Google Scholar]
- Zhang, Y.; Wang, J.Z.; Wu, Y.J.; Li, W.G. Anti-inflammatory effect of recombinant human superoxide dismutase in rats and mice and its mechanism. Acta Pharmacol. Sin. 2002, 23, 439–444. [Google Scholar]
- Luisa, C.M.; Jorge, J.C.; Van’T, H.R.; Cruz, M.E.; Crommelin, D.J.; Storm, G. Superoxide dismutase entrapped in long-circulating liposomes: Formulation design and therapeutic activity in rat adjuvant arthritis. BBA Biomembr. 2002, 1564, 227–236. [Google Scholar] [CrossRef]
- Cloarec, M.; Caillard, P.; Provost, J.C.; Dever, J.M.; Elbeze, Y.; Zamaria, N. GliSODin, a vegetal sod with gliadin, as preventative agent vs. atherosclerosis, as confirmed with carotid ultrasound-B imaging. Eur. Ann. Allergy Clin. Immunol. 2007, 39, 45–50. [Google Scholar]
- Muth, C.M.; Glenz, Y.; Klaus, M.; Radermacher, P.; Speit, G.; Leverve, X. Influence of an orally effective SOD on hyperbaric oxygen-related cell damage. Free Radic. Res. 2004, 38, 927–932. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-X.; Zhou, D.-Y.; Liu, Z.-Q.; Lu, T.; Song, L.; Li, D.-M.; Dong, X.-P.; Qi, H.; Zhu, B.-W.; Shahidi, F. Structural and biochemical changes in dermis of sea cucumber (Stichopus japonicus) during autolysis in response to cutting the body wall. Food Chem. 2018, 240, 1254–1261. [Google Scholar] [CrossRef] [PubMed]
- Jamieson, A.J.; Gebruk, A.; Fujii, T.; Solan, M. Functional effects of the hadal sea cucumber Elpidia atakama (Echinodermata: Holothuroidea, Elasipodida) reflect small-scale patterns of resource availability. Mar. Biol. 2011, 158, 2695–2703. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Lin, S.; Zeng, R. Cloning, expression, and characterization of a cold-adapted lipase gene from an antarctic deep-sea psychrotrophic bacterium, Psychrobacter sp. 7195. J. Microbiol. Biotechnol. 2007, 17, 604. [Google Scholar] [PubMed]
- Michels, P.C.; Clark, D.S. Pressure-enhanced activity and stability of a hyperthermophilic protease from a deep-sea methanogen. Appl. Environ. Microbiol. 1997, 63, 3985–3991. [Google Scholar] [PubMed]
- Borgstahl, G.E.O.; Parge, H.E.; Hickey, M.J.; Beyer, W.F., Jr.; Hallewell, R.A.; Tainer, J.A. The structure of human mitochondrial manganese superoxide dismutase reveals a novel tetrameric interface of two 4-helix bundles. Cell 1992, 71, 107–118. [Google Scholar] [CrossRef]
- Liu, P.; Ewis, H.E.; Huang, Y.J.; Lu, C.D.; Tai, P.C.; Weber, I.T. Crystal Structure of the Bacillus subtilis Superoxide Dismutase. Acta Crystallogr. 2008, 63 Pt 12, 1003–1007. [Google Scholar]
- Benkert, P.; Biasini, M.; Schwede, T. Toward the estimation of the absolute quality of individual protein structure models. Bioinformatics 2011, 27, 343–350. [Google Scholar] [CrossRef]
- Liu, J.; Yin, M.; Hu, Z.; Lu, J.; Cui, Z. Purification and characterization of a hyperthermostable Mn-superoxide dismutase from Thermus thermophilus HB27. Extremophiles 2011, 15, 221–226. [Google Scholar] [CrossRef]
- Zhu, Y.B.; Wang, G.H.; Ni, H.; Xiao, A.F.; Cai, H.N. Cloning and characterization of a new manganese superoxide dismutase from deep-sea thermophile Geobacillus sp. EPT3. World J. Microbiol. Biotechnol. 2014, 30, 1347–1357. [Google Scholar] [CrossRef]
- Song, N.N.; Zheng, Y.; Shi-Jin, E.; Li, D.C. Cloning, expression, and characterization of thermostable Manganese superoxide dismutase from Thermoascus aurantiacus var. levisporus. J. Microbiol. 2009, 47, 123–130. [Google Scholar] [CrossRef] [PubMed]
- Dolashki, A.; Abrashev, R.; Stevanovic, S.; Stefanova, L.; Ali, S.A.; Velkova, L.; Hristova, R.; Angelova, M.; Voelter, W.; Devreese, B. Biochemical properties of Cu/Zn-superoxide dismutase from fungal strain Aspergillus niger 26. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2009, 71, 975–983. [Google Scholar] [CrossRef] [PubMed]
- Bao, Y.; Li, L.; Xu, F.; Zhang, G. Intracellular copper/zinc superoxide dismutase from bay scallop Argopecten irradians: Its gene structure, mRNA expression and recombinant protein. Fish Shellfish Immunol. 2009, 27, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Kan, G.; Wen, H.; Wang, X.; Zhou, T.; Shi, C. Cloning and characterization of iron-superoxide dismutase in Antarctic yeast strain Rhodotorula mucilaginosa AN5. J. Basic Microbiol. 2017, 57, 680–690. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Kong, X.; Chen, J.; Liu, H.; Zhang, H. Characteristics of the Copper, Zinc Superoxide Dismutase of a Hadal Sea Cucumber (Paelopatides sp.) from the Mariana Trench. Mar. Drugs 2018, 16, 169. [Google Scholar] [CrossRef] [PubMed]
- Kawano, H.; Nakasone, K.; Matsumoto, M.; Yoshida, Y.; Usami, R.; Kato, C.; Abe, F. Differential pressure resistance in the activity of RNA polymerase isolated from Shewanella violacea and Escherichia coli. Extremophiles 2004, 8, 367–375. [Google Scholar] [CrossRef]
- Wang, S.L.; Li, Y.L.; Han, Z.; Chen, X.; Chen, Q.J.; Wang, Y.; He, L.S. Molecular Characterization of a NovelN-Acetylneuraminate Lyase from a Deep-Sea Symbiotic Mycoplasma. Mar. Drugs 2018, 16, 80. [Google Scholar] [CrossRef]
- Brindley, A.A.; Pickersgill, R.W.; Partridge, J.C.; Dunstan, D.J.; Hunt, D.M.; Warren, M.J. Enzyme sequence and its relationship to hyperbaric stability of artificial and natural fish lactate dehydrogenases. PLoS ONE 2008, 3, e2042. [Google Scholar] [CrossRef]
- Abe, F.; Minegishi, H.; Miura, T.; Nagahama, T.; Usami, R.; Horikoshi, K. Characterization of cold- and high-pressure-active polygalacturonases from a deep-sea yeast, Cryptococcus liquefaciens strain N6. J. Agric. Chem. Soc. 2006, 70, 296–299. [Google Scholar]
- Sujiwattanarat, P.; Pongsanarakul, P.; Temsiripong, Y.; Temsiripong, T.; Thawornkuno, C.; Uno, Y.; Unajak, S.; Matsuda, Y.; Choowongkomon, K.; Srikulnath, K. Molecular cloning and characterization of Siamese crocodile (Crocodylus siamensis) copper, zinc superoxide dismutase (CSI-Cu,Zn-SOD) gene. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2016, 191, 187–195. [Google Scholar] [CrossRef]
- Zhu, Y.; Li, H.; Ni, H.; Liu, J.; Xiao, A.; Cai, H. Purification and biochemical characterization of manganesecontaining superoxide dismutase from deep-sea thermophile Geobacillus sp. EPT3. Acta Oceanol. Sin. 2014, 33, 163–169. [Google Scholar] [CrossRef]
- Ken, C.F.; Hsiung, T.M.; Huang, Z.X.; Juang, R.H.; Lin, C.T. Characterization of Fe/Mn−Superoxide Dismutase from Diatom Thallassiosira weissflogii: Cloning, Expression, and Property. J. Agric. Food Chem. 2005, 53, 1470–1474. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Divalent Metal Ions | Concentration/mmol·L−1 | Relative Activity/% |
|---|---|---|
| Control | — | 100 ± 2.39 |
| Mn2+ | 0.1 | 92.89 ± 1.53 ** |
| 1 | 84.99 ± 2.77 ** | |
| Co2+ | 0.1 | 80.13 ± 1.23 ** |
| 1 | 61.49 ± 1.54 ** | |
| Ni2+ | 0.1 | 95.70 ± 2.38 ** |
| 1 | 94.42 ± 2.92 ** | |
| Zn2+ | 0.1 | 90.25 ± 1.76 ** |
| 1 | 90.99 ± 4.63 ** | |
| Cu2+ | 0.1 | 98.39 ± 3.97 |
| 1 | 88.99 ± 5.44 ** | |
| Ba2+ | 0.1 | 99.16 ± 2.18 |
| 1 | 95.94 ± 2.40 ** | |
| Mg2+ | 0.1 | 100.68 ± 3.27 |
| 1 | 100.61 ± 2.16 | |
| Ca2+ | 0.1 | 99.71 ± 1.13 |
| 1 | 100.39 ± 4.48 |
| Divalent Metal Ions | Concentration | Relative Activity/% |
|---|---|---|
| Control | — | 100 ± 2.84 |
| EDTA | 1 mmol·L−1 | 64.33 ± 3.08 ** |
| 10 mmol·L−1 | 58.03 ± 2.59 ** | |
| DTT | 1 mmol·L−1 | 96.36 ± 4.65 |
| 10 mmol·L−1 | 97.00 ± 5.46 | |
| β-ME | 1 mmol·L−1 | 96.53 ± 4.47 |
| 10 mmol·L−1 | 101.85 ± 3.72 | |
| Tween 20 | 0.1% | 109.96 ± 6.62 ** |
| 1% | 105.01 ± 3.28 ** | |
| Chaps | 0.1% | 103.13 ± 2.32 * |
| 1% | 99.32 ± 3.66 | |
| Triton X-100 | 0.1% | 105.02 ± 3.2 9** |
| 1% | 99.22 ± 3.79 | |
| SDS | 0.1% | 5.21 ± 3.45 ** |
| 1% | 6.11 ± 4.15 ** |
| Time (h) | Relative Activity (%) |
|---|---|
| 0 | 100 ± 2.21 |
| 1 | 108.66 ± 5.70 |
| 2 | 104.46 ± 4.54 |
| 3 | 103.73 ± 3.24 |
| 4 | 106.72 ± 4.80 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Kong, X.; Zhang, H. Characteristics of a Novel Manganese Superoxide Dismutase of a Hadal Sea Cucumber (Paelopatides sp.) from the Mariana Trench. Mar. Drugs 2019, 17, 84. https://doi.org/10.3390/md17020084
Li Y, Kong X, Zhang H. Characteristics of a Novel Manganese Superoxide Dismutase of a Hadal Sea Cucumber (Paelopatides sp.) from the Mariana Trench. Marine Drugs. 2019; 17(2):84. https://doi.org/10.3390/md17020084
Chicago/Turabian StyleLi, Yanan, Xue Kong, and Haibin Zhang. 2019. "Characteristics of a Novel Manganese Superoxide Dismutase of a Hadal Sea Cucumber (Paelopatides sp.) from the Mariana Trench" Marine Drugs 17, no. 2: 84. https://doi.org/10.3390/md17020084
APA StyleLi, Y., Kong, X., & Zhang, H. (2019). Characteristics of a Novel Manganese Superoxide Dismutase of a Hadal Sea Cucumber (Paelopatides sp.) from the Mariana Trench. Marine Drugs, 17(2), 84. https://doi.org/10.3390/md17020084