1. Introduction
Before the 1950s, seaweeds were used as traditional folk medicines [
1]. Over the past two decades, polysaccharides of brown seaweeds, such as fucoidan, alginate, and laminarin, have been investigated for their biological activities [
2,
3]. The health-promoting effects of these compounds, especially fucoidan, give brown seaweed great value for developing natural dietary supplements.
Fucoidans are a class of fucose-containing sulfated polysaccharides found in brown seaweed [
4]. Most fucoidans have complex chemical compositions, and their structures and biological activities vary from species to species [
5]. Its non-animal origin has been related to particular pharmacological activities [
6]. Fucoidan has been associated with antitumor [
7,
8,
9], antiviral [
10], anti-inflammatory [
11,
12], anticoagulant [
13], and osteogenic-enhancing differentiation activities [
14]. These activities are closely related to its molecular weight [
15] and sulfate content [
16].
Fucoidan’s immunomodulatory effects have been reported in different experimental models. It has been shown to activate and promote the maturation of human monocyte-derived dendritic cells [
17], and stimulate lymphocyte [
18,
19] and macrophage activity [
18,
20] in vitro. According to Do et al. [
21], fucoidan is an immunomodulating nutrient which alters the sensitivity of glia and macrophages. Jin et al. [
22] found that intraperitoneal (i.p.) fucoidan injection in mice upregulated CD40, CD80, and CD86 expression. It also increased production of interleukin (IL)-6, IL-12, and tumor necrosis factor-α (TNF-α) in spleen dendritic cells (DC). Furthermore, fucoidan promoted interferon (IFN)-γ generation by producing Th1 and Tc1 cells in an IL-12-dependent manner. Zhang et al. [
23] also showed that i.p. administration of fucoidan in the mice promoted natural killer (NK) cell, DC, and T-cell activation. In addition, when mice are injected IP with ovalbumin (OVA) and fucoidan, mice were found to produce remarkably higher amounts of anti-OVA Immunoglobulin G (IgG) than OVA-control mice, and fucoidan was found to promote the generation of effector/memory T-cells based on the surface expression of CD44. Intravenous administration of fucoidan upregulated CD40, CD80, CD86, and the major histocompatibility complex in mouse spleen DCs [
23]. Interestingly, oral administration of 300 mg fucoidan per day to elderly men and women increased their immune responses to influenza vaccination [
24], but the underlying mechanisms of this are still unclear.
There are some reports on the relationship between the structure and immunomodulatory properties of fucoidan [
25]; however, less is known about the relationship between their molecular weight and immunomodulatory properties. In our previous studies, we demonstrated that low-molecular-weight (LMW) fucoidan has more favorable bioactivity in vitro and in vivo than high-molecular-weight fucoidan [
7,
8,
9,
12], with no toxicological effects found in rats after 28 days of repeated oral administration [
26]. LMW fucoidan is expected to be a safe food supplement for immunomodulation.
Mycoplasma pneumoniae is one of the most common agents of community-acquired pneumonia in previously healthy people, and infection can occur at any age [
27].
M. pneumoniae is considered a self-limiting disease, but some patients suffer from protracted and complicated courses [
28]. In addition,
M. pneumoniae can change its cell membrane composition to mimic the host’s cell membrane, thus avoiding immune system detection [
27]. Currently, there is no vaccine to prevent
M. pneumoniae infection. Macrolide is one of the first-line drugs used to treat
M. pneumoniae, but the prevalence of macrolide-resistant
M. pneumonia is increasing worldwide. In 2011, the macrolide-resistance rate was up to 23% in Taiwan [
29]. A recent study has reported that early additional immune-modulators might can prevent
M. pneumoniae disease progression and reduce morbidity [
30]. A natural and safe food supplement that enhances the immune response to prevent
M. pneumoniae infection could address this unmet need.
In this study, we evaluated the immunomodulatory effects of orally administered LMW fucoidan on innate and adaptive immune responses in mice. We also evaluated the effect of LMW fucoidan supplementation on immune responses to the M. pneumonia (MP) antigen.
3. Materials and Methods
3.1. Animal
The female BALB/c mice (Albino mice maintained by Halsey Bagg) were obtained from BioLASCO Taiwan CO., Ltd (Taipei, Taiwan). All mice were weighted and given provisional numbers upon arrival. The animals were subjected to acclimation in a housing facility for at least eight days before starting the experiment. Only healthy mice were selected for use in the study. The mice were eight weeks of age at the start of the experiment, and were housed in a normal, environmentally controlled animal room with free access to pathogen-free feed and water ad libitum. These mice were randomly divided, with ten mice in each group.
3.2. LMW Fucoidan Preparation
LMW fucoidan extracted from
Laminaria japonica were collaboratively manufactured by Hi-Q Marine Biotech International Ltd. (New Taipei City, Taiwan). Seaweed samples were ground to flour with a miniblender, and then dried with a dryer at 50 °C. The 100 g dried seaweed was treated with 5 L of distilled water and boiled at 100 °C for 30 min, and the extract was centrifuged at 10,000×
g for 20 min. The supernatant was added with 4 M CaCl
2 incubated for 1 h to separate alginic acid, and re-centrifuged at 10,000×
g for 20 min. All polysaccharides were dialyzed (cut off 10,000 Da) using deionized water for 48 h, and precipitated by the addition of ethanol at the ratio of 1:3 (V/V) and left overnight to give the crude fucoidan. The fucoidan so obtained was fractionated by anion-exchange chromatography using DEAE-Sephadex A-25 (Cl
− form, Pharmacia, Uppsala, Sweden) using gradually increasing concentrations of sodium chloride (0–2.5 M) at a flow rate of 1 mL/min, and eluents (5 mL/tube) were separately collected. Each fraction was analyzed for sulfate content (
Section 3.3) and monosaccharide composition (
Section 3.4). The fraction of rich sulfate and fucose content (fraction 3) was extracted and hydrolyzed with a mixture of crude glycolytic enzymes, isolated from
Bacillus subtilis by our lab (
Section 3.5). The reaction mixture was passed through a filter (3000 Da cut off) by using Amicon Stirred Cells (Merck Germany) to collect the LMW fucoidan with a molecular weight lower than 3000 Da. The hydrolysis reaction of the fucoidan by the crude glycolytic enzymes was performed at pH 6.5, 37 °C for 24 h and heated at 95 °C for 20 min for inactivation of the enzyme.
3.3. Analysis of Degree of Sulfation
Sulfate content was determined according to the gelatinbarium method [
53] using sodium sulfate (1 mg/mL) as a standard after acid hydrolysis of the polysaccharide extract sample in 6 N HCl at 100 °C for 6 h.
3.4. Monosaccharide Composition Analysis
The monosaccharides compositions of the eluted fractions from anion-exchange chromatography were separated by a High-performance anion-exchange chromatography (HPAEC) (Dionex BioLC system) using an anion-exchange column (DionexTM Carbopac PA-10, 4 × 250 mm, Dionex, Thermo Fisher Scientific, Waltham, MA, USA). The analysis of monosaccharides was carried out at an isocratic NaOH concentration of 18 mM at ambient temperature [
54].
3.5. Preparation of the Crude Enzyme Mixtures from B. subtilis Cultures
One colony of B. subtilis, isolated from natto in Keelung local market, was cultivated in tryptic soy broth (TSB, Becton, Dickinson and Company, Sparks, MD, USA) at 30 °C, 150 rpm for 24 h. The cell cultures were centrifuged to remove bacterial cells to obtain the mixtures of extracellular glycosidic enzymes.
3.6. Animal Experiments
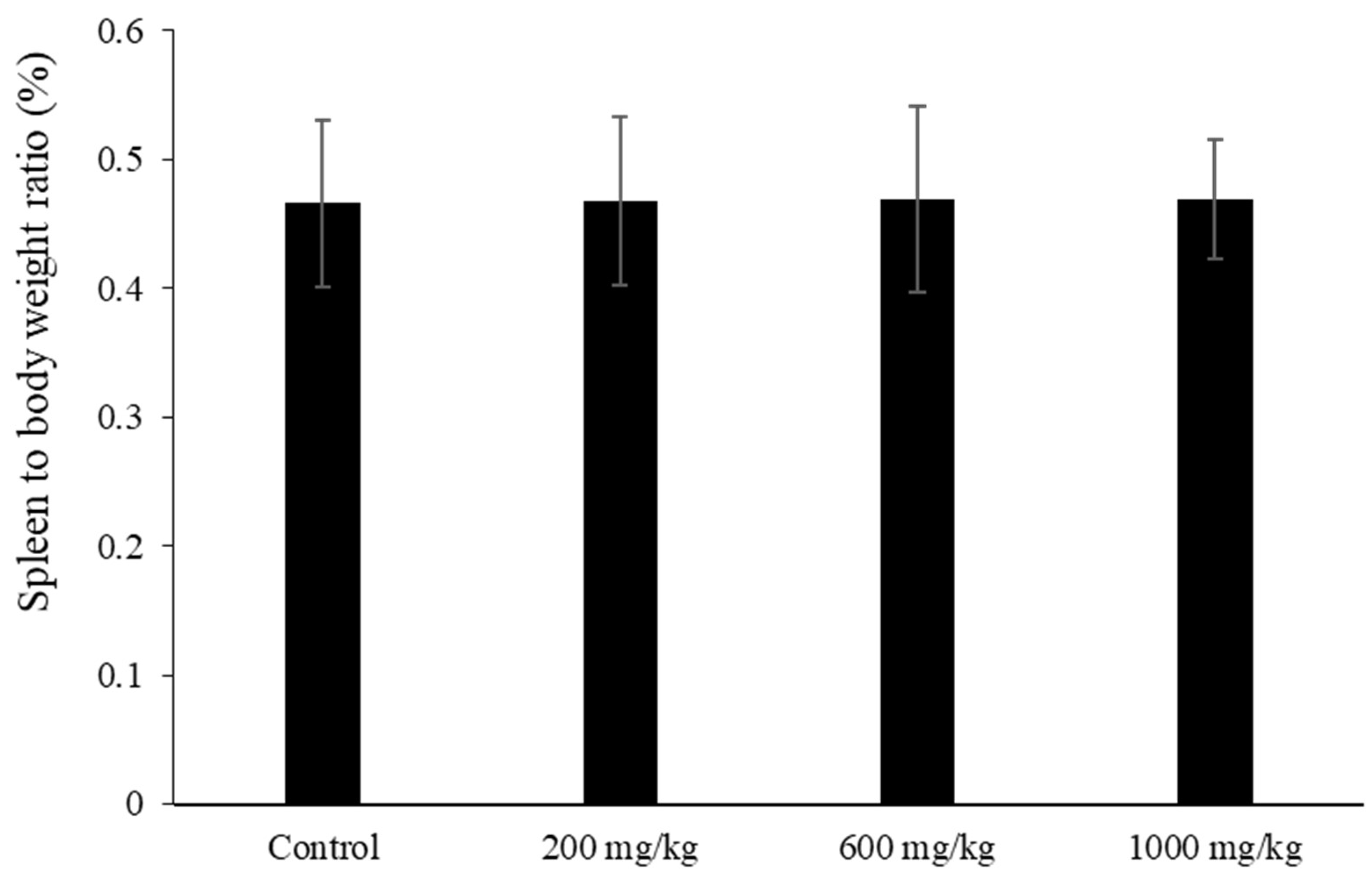
The LMW fucoidan was a light brownish-white powder, and well-soluble below the highest concentration. In this study, three independent experiments were undertaken to evaluate the innate immune, adaptive immune, and MP antigen-inoculated immune response, respectively. The first experiment was performed to assay innate immune, mice were assigned to four groups (n = 10); concentrations of 200, 600, and 1000 mg/kg LMW fucoidan were dissolved in distilled water and then administered by oral gavage in 10 mL/kg on a daily basis for 6 weeks. The control group (0 mg/kg LMW fucoidan) mice were treated identically with equal volumes of distilled water also via oral gavage throughout the study. Mice were weighted and sacrificed, and blood samples were collected at the end of the experiment for serum immunoglobulins assay. Spleens were collected and weighted, and the splenocytes were analyzed for cell proliferation, the cytokines secretion assay, and natural killer cell activity assay. Peritoneal cells were collected form mice abdominal cavity for the phagocytic activity assay. Each mouse was weighted once a week during the study period.
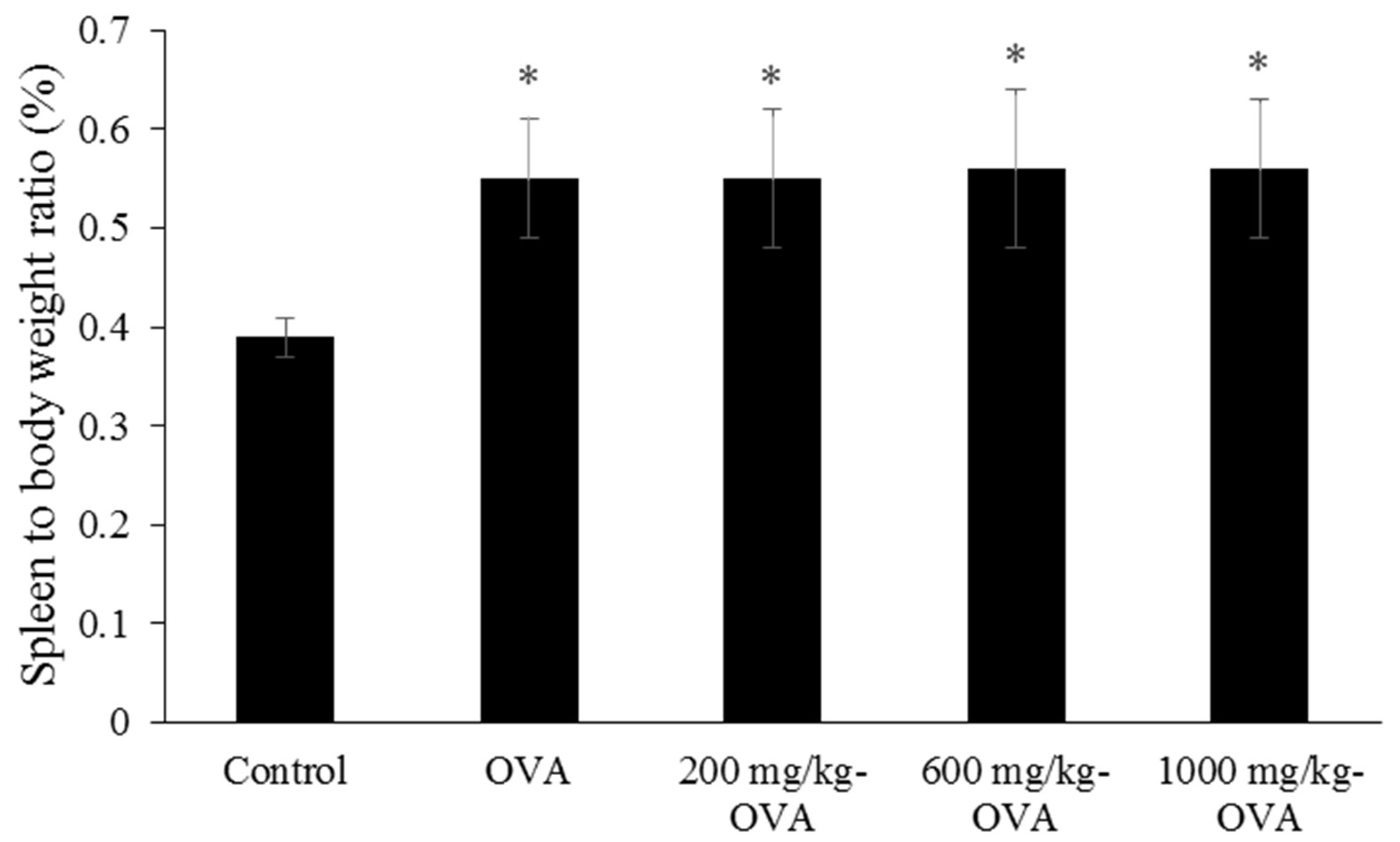
The second experiment was performed byadaptive immune mice assay, and five groups of mice (control, OVA-immunized, 200, 60,0 and 1000 mg/kg LMW fucoidan, then OVA-immunized) were orally administered distilled water or LMW fucoidan for 6 weeks. The first OVA IP immunization was conducted after 3 weeks of administration. 6.25 μg OVA was emulsified in the complete Freund’s adjuvant (CFA) and intraperitoneally injected into the mice. Two weeks after the first OVA immunization, mice were given a second injection of 12.5 μg OVA emulsified with incomplete Freund’s adjuvant (IFA) in order to enhance the OVA-specific immune responses. Blood samples were collected before the OVA inducement at the end of the experiment for the serum immunoglobulins assay. Mice were then sacrificed by isoflurane inhalation. Spleens were collected and weighted, and the splenocytes were analyzed for cell proliferation and the cytokines secretion assay.. Each mouse was weighted once a week during the study period.
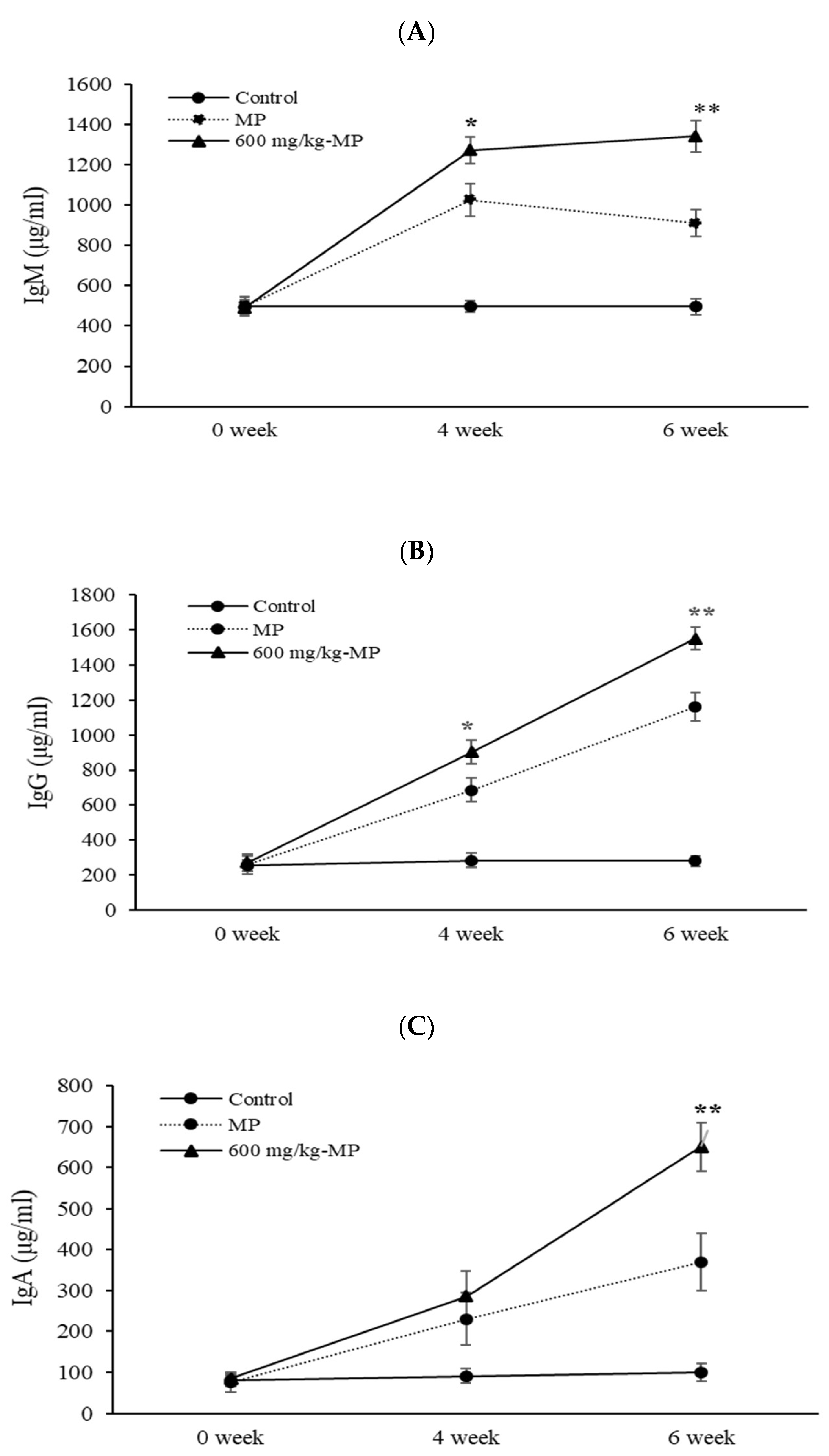
The third experiment was performed to assay MP antigen-inoculated immune responds, three groups of mice (control, MP antigen-stimulation and 600 mg/kg LMW fucoidan then MP antigen stimulation) were orally administered distilled water or LMW fucoidan for 6 weeks. First, MP antigen (10 μg/mouse, MP901, Native Antigen, UK) IP stimulation was conducted after 3 weeks of administration. Two weeks after the first stimulation, mice were given a second injection of MP antigen (10 μg/mouse) [
40]. Blood samples were collected before the first MP antigen stimulation, after one week of stimulation, and at the end of the experiment for the serum immunoglobulins assay. Each mouse was weighted once a week during the study period.
3.7. Splenocyte Proliferation Assay
Splenocytes were prepared from treated mice and resuspended in Roswell Park Memorial Institute (RPMI) 1640 medium supplemented with 10% fetal bovine serum (FBS), 50 U/mL penicillin, 50 mg/mL streptomycin, and 0.5 mg/mL amphotericin B. 2 × 105 cells/well of splenocytes were treated with Con A or LPS for 72 h in an innate immune experiment, and treated with Con A, LPS, or OVA for 72 h in an adaptive immune experiment. Cell proliferation was measured by OD490 using the CellTiter 96 Aqueous One Solution Cell Proliferation Assay (Promega). Results were expressed as the stimulation index, and the formula for calculating stimulation index is shown below:
Stimulation index = OD490 of Con A, LPS, or OVA-stimulated cells/OD490 of un-stimulated cells.
3.8. NK Cell Activity Aassay
NK cell activity was measured by the release of the fluorescent marker from YAC-1 target cells (A lymphoma which was induced by inoculation of the Moloney leukemia virus (MLV) into a newborn A/Sn mouse) [American Type Culture Collection (ATCC) TIB-160, Food Industry Research and Development Institute, Hsinchu, Taiwan]. Splenocytes (effector cells, including NK cells) were cultured in the supplemented RPMI 1640 medium. YAC-1 cells were incubated with 2′-7′-bis (carboxyethyl)-5(6)-carboxyfluorescein (BCECF) (ThermoFisher) for 30 min at 37 °C. After being washed with culture medium, cells were re-suspended in the medium and cultured for 2 h at 37 °C. 2′,7′ –bis-(Carboxyethyl)-5-(and-6)-carboxyfluorescein (BCECF)-labeled YAC-1 cells were mixed with splenocytes as effector cells to make a 10 and 25 ratio of effector cells to target cells (E/T). After incubation for 4 h, the cultured supernatant was harvested and measured fluorescence with excitation and emission wavelengths of 485 nm and 538 nm. The total target cell fluorescence was determined by lysis of cells with 0.25% (v/v) Triton X-100. Spontaneous fluorescence was measured from target cells incubated without effector cells. Test group fluorescence was measured from the effector cell mixing with target cells. Results were expressed as NK cell activity, and the formula for calculating NK cell activity is shown below:
NK cell activity = (test group fluorescence-spontaneous fluorescence)/(total target cell fluorescence- spontaneous fluorescence) × 100%.
3.9. Phagocytic Activity Assay by Peritoneal Cells
Mice were scarified and peritoneal cells were harvested by injecting 5 mL of Hank’s balanced salt solution (HBSS) into the peritoneal cavity and gently shaking it. Then, peritoneal fluid was collected and centrifuged at 450× g, 4 °C for 10 min. The cell pellet was resuspended at 1 × 106 cells/mL in supplemented RPMI 1640 medium. The phagocytosis was performed by monitoring the engulfment of FITC-labeled E. coli by the peritoneal exudate cells. Cells were mixed with FITC-E. coli at Multiplicity of infection (MOI) of 12.5 and 25 for 30 min. Fluorescence from non-internalized FITC-E. coli was quenched by the addition of acidified trypan blue, and the cells were then washed and analyzed by flow cytometry (BD FACSAria, Becton Dickinson Biosciences, San Jose, CA, US). Results were expressed as phagocytic activity, and the formula for calculating phagocytic activity is shown below:
Phagocytic activity = FITC positive peritoneal cells/total peritoneal cells × 100%.
3.10. Cytokines Secretion Assay
Cytokines including IL-2, IL-4, IL-5, TNF-α, and IFN-γ were measured by using enzyme-linked immunosorbent assay (ELISA) kits (R&D Systems Inc., Minneapolis, MN, USA) according to the manufacturer’s instructions. Splenocytes were prepared from treated mice and resuspended in supplemented RPMI 1640 medium. Cells were incubated at 1 × 106 cells/well with 25 μg/mL OVA for 72 h in the second experiment for adaptive immune assay. After 72 h incubation, the cell-free supernatants were collected, centrifuged 450× g, 4°C for 10 min, and cytokines were measured by using the ELISA kits.
3.11. Serum Immunoglobulins Assay
In the second experiment for the adaptive immune assay, blood was centrifuged at 450×
g, 4 °C for 10 min, and serum samples were collected for OVA-specific anti-IgG, OVA-specific anti-IgA, and OVA-specific anti-IgE. OVA-specific antibodies were detected by an indirect ELISA (Bethyl Laboratories). Results were expressed as ELISA units (EU), and the formula for calculating EU is shown below: EU = (OD450 of sample- OD
450nm of blank)/(OD450 of positive- OD450 of blank). A positive OD450 indicated a serum with a high titer of OVA-specific antibodies [
52]. In the third experiment, serum samples were collected for serum immunoglobulins assay by using a mouse immunoglobin quantitation set (Bethyl Laboratories) to measure the concentrations of IgM, IgG, and IgA, respectively.
3.12. Statistical Analysis
Numerical data are presented as means ± standard deviation. The data was analyzed by a one-way analysis of variance (ANOVA) and Student’s t-test.
4. Conclusions
In this study, we investigated the immune modulatory effects of orally administrated LMW fucoidan on the innate immune, adaptive immune, and MP antigen-stimulated immune response. Our results provided evidence that LMW fucoidan could increase splenocyte proliferation, NK cell activity, and phagocytic activity, as well as IL-2, IL-4, and IFN-γ secretion. In addition, LMW fucoidan enhanced antigen-specific antibody production in OVA- and MP antigen-stimulated mice. Our hope is that LMW fucoidan, a natural food supplement, could enhance immune responses, be needed for immunopotentiation, and attenuate the M. pneumoniae infectious disease.








{kind=link}
{kind=link}
{kind=link}
{kind=link}